Abstract
Flowering time (heading date) of the rice plant is considered an important agronomic trait for environmental adaptation and grain yield. It is controlled by multiple genes and is regulated by different environmental factors, such as day length, temperature, soil moisture, etc. So far, approximately 125 genes regulating flowering process and floral organ identity or development directly or indirectly have been reported in rice. Among these genes, Heading date 3a (Hd3a), RICE FLOWERING LOCUS T1 (RFT1), Heading date 5 (Hd5), MORF-RELATED GENE702 (MRG702), Casein kinases, CKI and CK2α, Pseudo-Response Regulator 37 (PRR37), Hd gene family have been reported as the key genes regulating flowering time in rice; however, their functions are mostly inter-related. Hd3a and RFT1 that encode florigens, are known as the floral transition genes in rice. In rice, florigen immediately induces downstream genes in the shoot apical meristem (SAM) to start the transition from vegetative to reproductive phase. RFT1 gene regulates flowering time with pivotal action while Hd3a, regulates under long day conditions. The Hd5 gene regulates flowering time in variation of early heading for adaptation depending on environmental signals. MRG702, a reader protein, promotes flowering. Casein kinases, CKI and CK2α directly influence the function of the early heading regulator PRR37. Hd16/CKI hinders flowering time in the Ehd1-concerned pathway through phosphorylation of Ghd7 and PRR37. Natural variants of Hd1, PRR37, Ghd7, DTH8, Hd6, and Hd16 were found in the rice varieties that are cultivated presently in Asia and Europe, and their variants play significant roles in the down streaming of Ehd1 expression to delay flowering time in natural long day (LD) conditions.
-
Key words: Enzymes, Florigen, Flowering time, Genes, Heading date, Rice
INTRODUCTION
Rice flowering time is triggered by both exogenous and endogenous signals and determines the cropping season and regional adaptability (
Tan et al. 2017). The initiation of flowering is one of the most significant transitions in the life of a rice plant. Genetic complexes that synchronize intrinsic developmental signals with environmental cues such as light-dark cycle and temperature control flowering time of rice plant. The regulation of flowering time by both internal and external signals ensures that flowering time coincides with the appropriate environmental conditions and leads to the successful reproduction of the rice plant (
Maas et al. 2010). Floral transition, the turnover between the vegetative and reproductive stages of plant development, is a vital step in the life span of all crop species. Florigen, the key hormone required for induction of flowering is synthesized in the leaves passing through inductive day length conditions and translocated to the shoot apex where it accelerates the flowering transition (
Naranjo et al. 2014). Rice plants possess two main flowering genes, Heading date 3a (
Hd3a) and RICE FLOWERING LOCUS T1 (
RFT1), and at least two flowering pathways to control the expression of florigen. Recent studies revealed that
RFT1 has pleiotropic effect on yield attributes and plays a key role on rice yield (
Xue et al. 2008;
Zhu et al. 2017). Heading date 1 (
Hd1) is orthologous between rice and Arabidopsis, and Early heading date 1 (
Ehd1) is unique to rice (
Doi et al. 2004). Among the Heading date-QTLs, several natural variants at the genic level have been reported by different researchers (
Se5 by
Izawa et al. 2000,
Hd1 by
Yano et al. 2000,
Hd6 by
Takahashi et al. 2001,
Hd3a by
Kojima et al. 2002,
Ehd1/Hd14 by
Doi et al. 2004,
Ghd7/Hd4 by
Xue et al. 2008,
Hd5/DTH8/Ghd8 by
Wei et al. 2010;
Yan et al. 2011;
Fujino et al. 2013, and
Ef7/OsELF3/Hd17 by
Matsubara et al. 2012;
Saito et al. 2012;
Zhao et al. 2012;
Koo et al. 2013). Molecular analysis revealed diverse combinations of natural variants in
Hd1 alleles, two types of
Hd3a promoter sequences, and various expression levels of
Ehd1 in the major collection of rice cultivars, leading to the diversification in flowering time (
Takahashi et al. 2009). In line with this, natural variation in
Ghd7/Hd4 plays a role in rice cultivation in temperate regions (
Xue et al. 2008). The highly conserved Ser/Thr-specific casein kinases (CKs) regulate various signal transduction processes in eukaryotic organisms (
Knippschild et al. 2005;
Mulekar and Huq 2014). In plants, CKI and CK2 play important roles in the regulation of flowering time and circadian rhythm. Many enzymes that methylate histone-Lys residues have been detected in plants, although the mechanisms to interpret histone methylation are still not well comprehended. The various modifications of histone residues/histone codes seem to serve as cues that are recognized by effector modules (reader proteins), which might implement the down regulating functions of histone codes (
Liu et al. 2010;
Berr et al. 2011;
Yun et al. 2011). Recent studies on such effector modules have revealed that a variety of reader domains, including chromodomain (chromatin organization modifier domain), plant homeodomain (PHD), etc. (
Jin et al. 2015), bind to different methylated Lys residues of histones (
Yun et al. 2011). Many studies showed that a large number of rice genes regulate flowering time through the two flowering integrators. Recently, a major histone methyltransferase (HMTase) gene SET DOMAIN GENE 724 (
SDG724) has been identified, which is required for Histone H3 lysine 36 (H3K36) methylation, promotes rice heading, indicating that flowering in rice might also be regulated by chromatin alteration (
Sun et al. 2012). Knowledge of functional roles of the genes regulating heading date might help understand the genetic mechanism of domestication and adaptation process of rice across geography that would help breeders and geneticists to develop appropriate rice varieties for the target environments. In this paper, we reviewed the functional role of the most recently reported genes and their alleles controlling heading date in rice plants.
Rice heading date is one of the most important determinants of rice adaptability to various seasons and geographical regions. It is a complex trait controlled by multiple genes and is influenced by different environmental factors, particularly day length, temperature, and soil moisture. Several quantitative trait loci (QTLs) controlling flowering time have been mapped using different diverse rice germplasms and several genes have been identified by positional cloning strategies (
Miura et al. 2011;
Tsuji et al. 2011;
Fujino et al. 2013). So far, approximately 125 genes were reported to be involved in the process of flowering in rice (
Table 1). However, the majority of these genes are inter-related and epistatically controlled for their functions. In a QTL mapping work using 12 diverse rice cultivars from Asia,
Ebana et al. (2011) showed that combinations of different alleles of QTLs for heading date could generate a large portion of the phenotypic variations in heading date. QTL analysis for very late heading from an
indica variety, Nona Bokra, suggested that strong photoperiod sensitivity of this variety was mainly due to additive effects of particular alleles (
Uga et al. 2007). It was found that genetic variations could be attributed to the coalition of alleles in loci involved in heading date (
Fujino et al. 2013). We describe the detailed functions of few key genes involved in the regulation of flowering time in the next few sections.
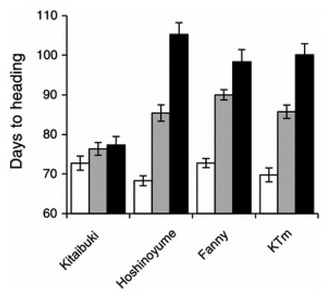
Hd5 gene was identified on the short arm of chromosome 8 from the F
2 population of a cross between Hayamasari and Hoshinoyume (
Nonoue et al. 2008). A previous study revealed that days to heading (DTH) difference between Kitaibuki (
Japonica) and Hoshinoyume was controlled by a single gene (
Fujino 2011). The sequence analysis showed that Kitaibuki carries a loss function of
Hd5 with a 19 bp deletion (Gen Bank accession number AB693201), the same as for Hayamasari, while Hoshinoyume carries the functional
Hd5 (
Fujino et al. 2013); and simultaneously,
Fujino et al. (2013) found differences in heading date and photoperiod sensitivity among different
japonica varieties (
Fig. 1). The transgenic lines overexpressed with functional
Hd5 gene showed delayed DTH than that of Hayamasari under long day (LD) conditions. These results clearly revealed that allelic variations in
Hd5 such as functional and loss-of-function are major factors influencing heading date in Hoshinoyume with Kitaibuki and Hayamasari. In a separate study in Hokkaido, Japan (2003–2004) with
Hd5 gene carrying 19 bp deletion and non-deletion showed a significant (
P = 0.0039) difference in flowering time between the two groups (
Fujino et al. 2013). These results revealed that the loss-of-function
Hd5 has a vital role in generating early heading on variations. Loss of-function of
Hd5 was also reported as a major determinant of early heading among the tested populations by
Kim et al. (2013) and could contribute to extremely early heading behavior in presence of (strong floral repressor)
Ghd7, while
Shibaya et al. (2011) reported loss-of-function of
qDTH7/ghd7 for extremely early heading varieties. In some cases,
Hd5 expressed epistatic interactions with
Hd1 and
Ghd7, thus causing these various effects on dates of heading (
Nonoue et al. 2008;
Xue et al. 2008;
Yan et al. 2011). Nevertheless, heading date genes such as
Hd5 control agronomic features like plant height and yield potential as well (
Wei et al. 2010;
Yan et al. 2011).
Flowering time controlling Heading date gene family and Early heading date 1 (Ehd1)
Heading-date 1 (
Hd1) is the first gene cloned through natural variation incur for rice flowering time (
Yano et al. 2000;
Hori et al. 2016). It is one of the principal flowering time controlling genes, which is an ortholog of Arabidopsis
CONSTANS (
CO), promoting transition to flower in short-day (SD) and suppresses it strongly in LD conditions (
Endo-Higashi and Izawa 2011).
Hd1 is homologous to
SE1 (Photoperiodic Sensitivity 1), one of the major QTLs for photoperiodic sensitivity. Los of-function of
Hd1 causes delayed flowering in SD but early flowering under long day conditions (
Inoue et al. 1992;
Lin et al. 2000;
Gomez-Ariza et al. 2015), demonstrating that
Hd1 may act either as a flowering activator or repressor, but depends on light periods. However, how
Hd1 functions differently between SD and LD is still unknown.
Ishikawa et al. (2005) observed that the overexpression of
Hd1 gene induced late-flowering but they did not notice any phenotypic expression in the OsphyB mutant background suggesting OsPhyB is necessary for the repressive function of
Hd1.
Hd1 is a member of the CO-like proteins family, which contains conserved B-box zinc fingers as well as CCT domains (
Robson et al. 2001;
Griffiths et al. 2003).
A strong flowering time regulating gene, Early heading date 1 (
Ehd1), induces floral transition under short day (SD) rather than in LD conditions (
Endo-Higashi and Izawa 2011;
Zhao et al. 2015).
Ehd1 is successively repressed by the upregulation of
Ghd7 under LD conditions; this partially describes the critical day length identification of Heading date 3a (
Hd3a), a rice florigen transcriptional control (
Itoh et al. 2010). In the case of crop production, controlling of flowering time is often encompassed with yield-related features, as it is speculated that the longer vegetative periods result in larger biomass. In practicality, it has been revealed that
Ghd7 affects grain number in field trials (
Xue et al. 2008). Flowering time genes also control the duration of vegetative growth of plants, resulting in a strong relationship between biomass and grain yield. Nonetheless, how flowering time genes influence biomass, plant architecture and yield is still not well understood. Since the flowering time gene expression pattern is regulated by both circadian clock and encircling light signals (
Itoh et al. 2010), in addition to surrounding temperatures, it is possible that adjacent environmental conditions may regulate the inflorescence architecture of plants through the effect of some flowering time genes. In fact, the flowering time locus is often considered as a trait of yield-related quantitative trait locus (QTL) in breeding science. For instance,
Ghd7 gene is a yield-related gene that can regulate grain number per panicle aside from its critical control of flowering time (
Xue et al. 2008).
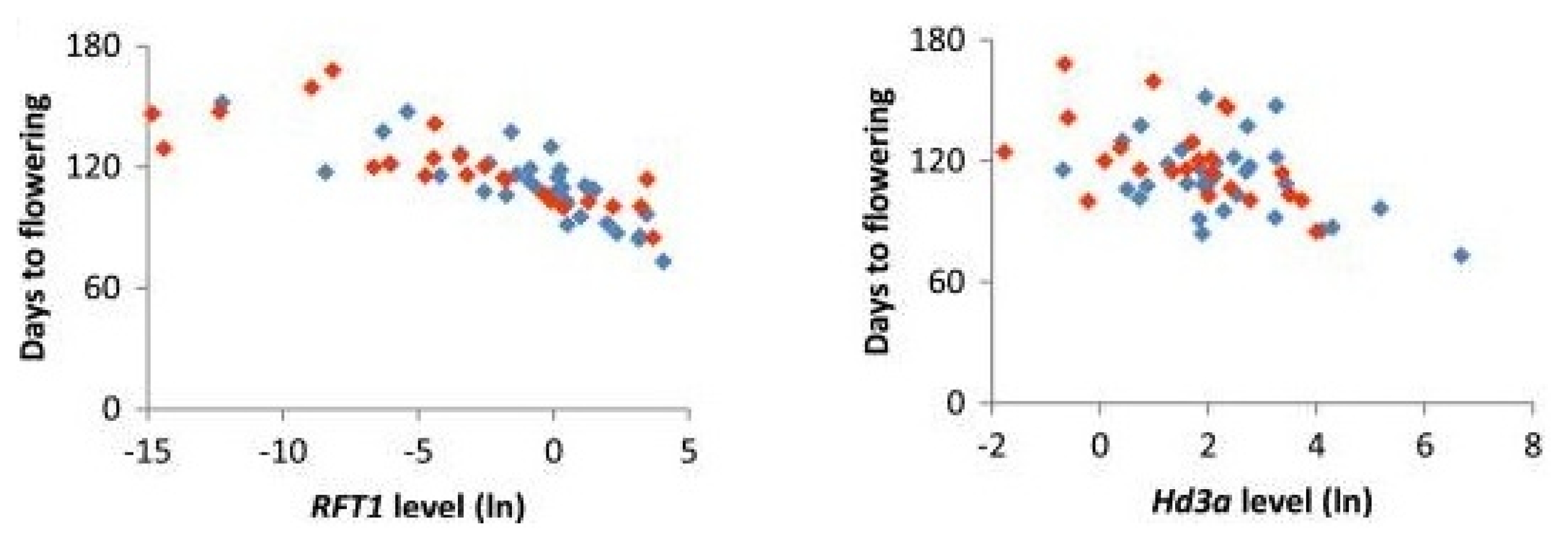
Heading date 3a (
Hd3a) is associated with heading date under SD condition. It can regulate flowering time under natural long day (NLD) also with the epistatic control of
Hd1 gene (
Naranjo et al. 2014;
Gomez-Ariza et al. 2015).
Hd3a encodes an ortholog of Arabidopsis FLOWERING LOCUS T (
FT). Over-expression of Hd3a protein with the constitutive promoter (
Kojima et al. 2002) or vascular-specific promoters results in an early flowering phenotype, and suppression of
Hd3a with RNAi delays flowering.
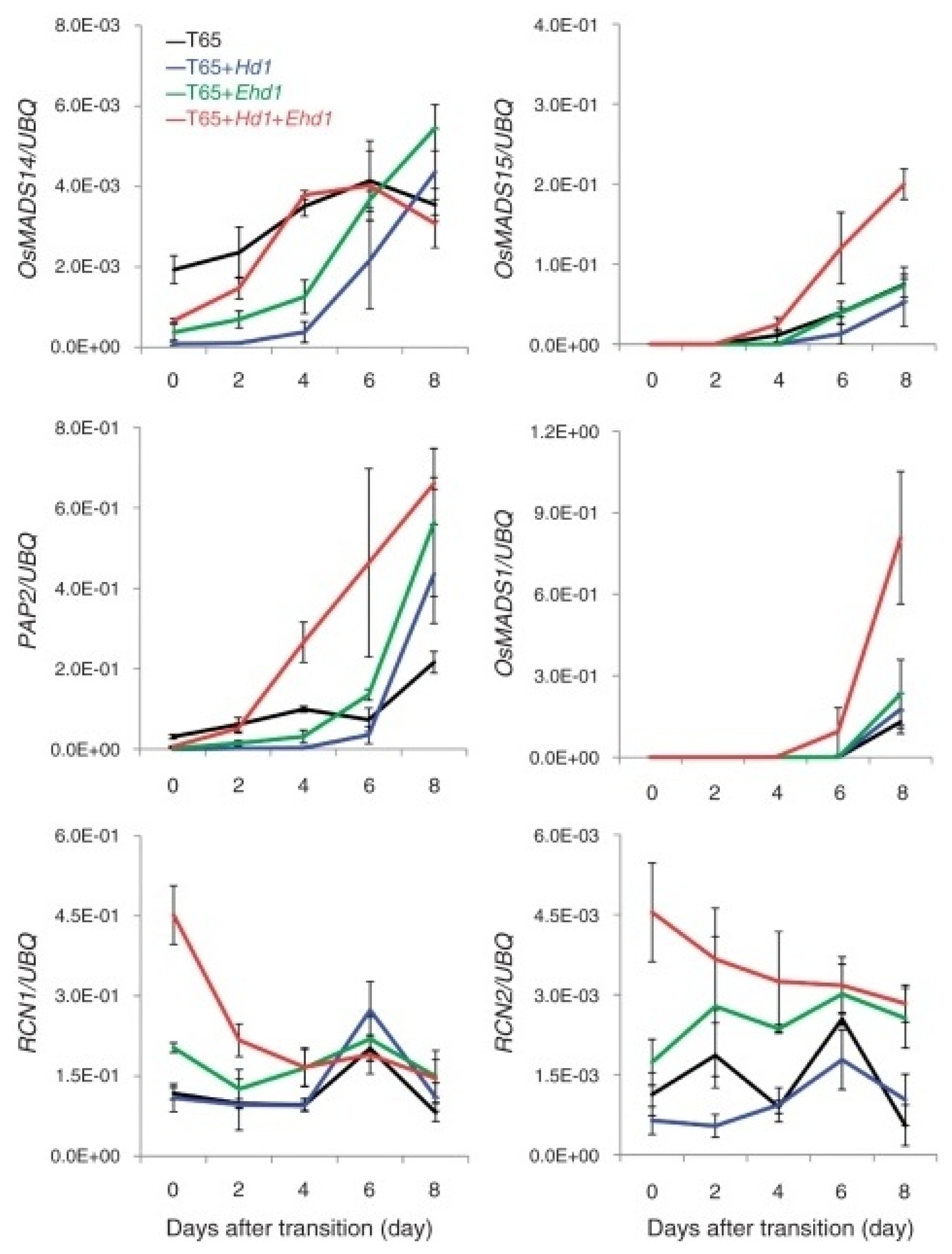
Endo-Higashi and Izawa (2011) reported that the gene expression at the SAMs during panicle development (
Fig. 2), induction of
Hd3a/RFT1 by
Hd1 and
Ehd1 under SD conditions accelerates floral transitions. However, this may not be sufficient to start the floral transitions at the apex since
Hd3a and
RFT1 expression levels in leaves varied at the same developmental stage during panicle development of rice (
Fig. 3). This may create some variations in protein levels of florigen (probably at the apex) after the initiation of floral transition between the plants and may cause precocious expression of panicle development related genes (
Endo-Higashi and Izawa 2011). This precocious expression of the MADS box genes involved in the development of smaller size of panicles. For example, over-expression of
OsMADS14 often results into flower-like organs under in vitro culture. It has been reported that the Terminal Flower 1 gene, an Arabidopsis
RCN1 and
RCN2 ortholog, is a motile signal to regulate plant architecture (
Conti and Bradley 2007). Differential expression of
RCN1 and
RCN2 under the control of
Hd1/Ehd1 may also be involved in panicle development of rice.
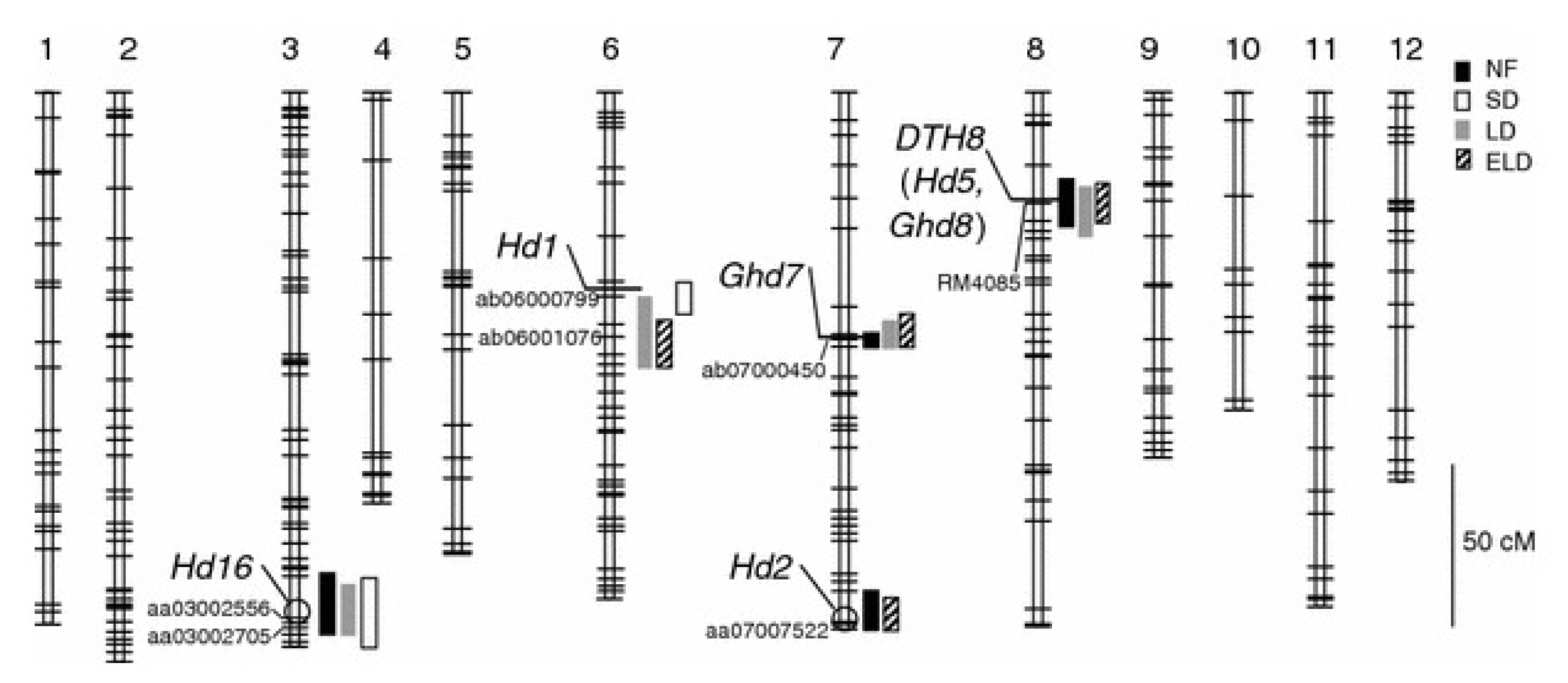
Heading date 2 (
Hd2), one of the QTLs for heading date, located at the distal end of the long arm of chromosome 7 was identified from a cross between Nipponbare (
japonica variety), and Kasalath (
indica variety). Two QTLs concerned to heading date,
Hd6 (
Takahashi et al. 2001) and
Hd16 (
Matsubara et al. 2008), which were previously reported at the distal end of chromosome number 3. In SD conditions, previously detected QTL on chromosome 6, and its position was found to be similar to the
Hd1 (
Yano et al. 2000;
Shibaya et al. 2011). Sequence analysis has revealed that Hayamasari has a functional
Hd1 allele not identical to that of Koshihikari (
Nonoue et al. 2008); and consequently, the QTL discerned on chromosome 6 under SD conditions was found to be
Hd1. In contrast, under LD and ELD conditions, the chromosomal positions of detected QTLs on chromosome 6 were near
Hd1 but not perfectly co-localized of the previously reported
Hd1 (
Fig. 4).
Three major regulators—
Hd1,
OsMADS50, and
Ghd7 suppress flowering mostly under LD condition. For instance,
Hd1 enhances flowering time under short days but delays flowering under long days (
Yano et al. 2000).
OsMADS50, a long day-oriented repressor up-regulates
Ehd1 (
Lee et al. 2004).
Ghd7 is another long day-preferential repressor that has been detected as a vital QTL that influences yield (
Xue et al. 2008). Altered functions of
Hd1 gene has been reported by
Zhang et al. (2017) under differential genetic backgrounds. They reported that heading date is enhanced by
Hd1 through upregulation of
Edh1,
Hd3a and
RFT1 expression under both SD and LD conditions (cv. Zhensan97), while it is suppressed by
Hd1 under LD conditions but is promoted in SD conditions in MH63 background. However,
Kim et al. (2013) observed that the levels of Hd1 and
OsMADS50 transcripts were not significantly altered in Kitaake (an extremely early flowering rice genotype from Hokkaido, Japan). The transcript levels of
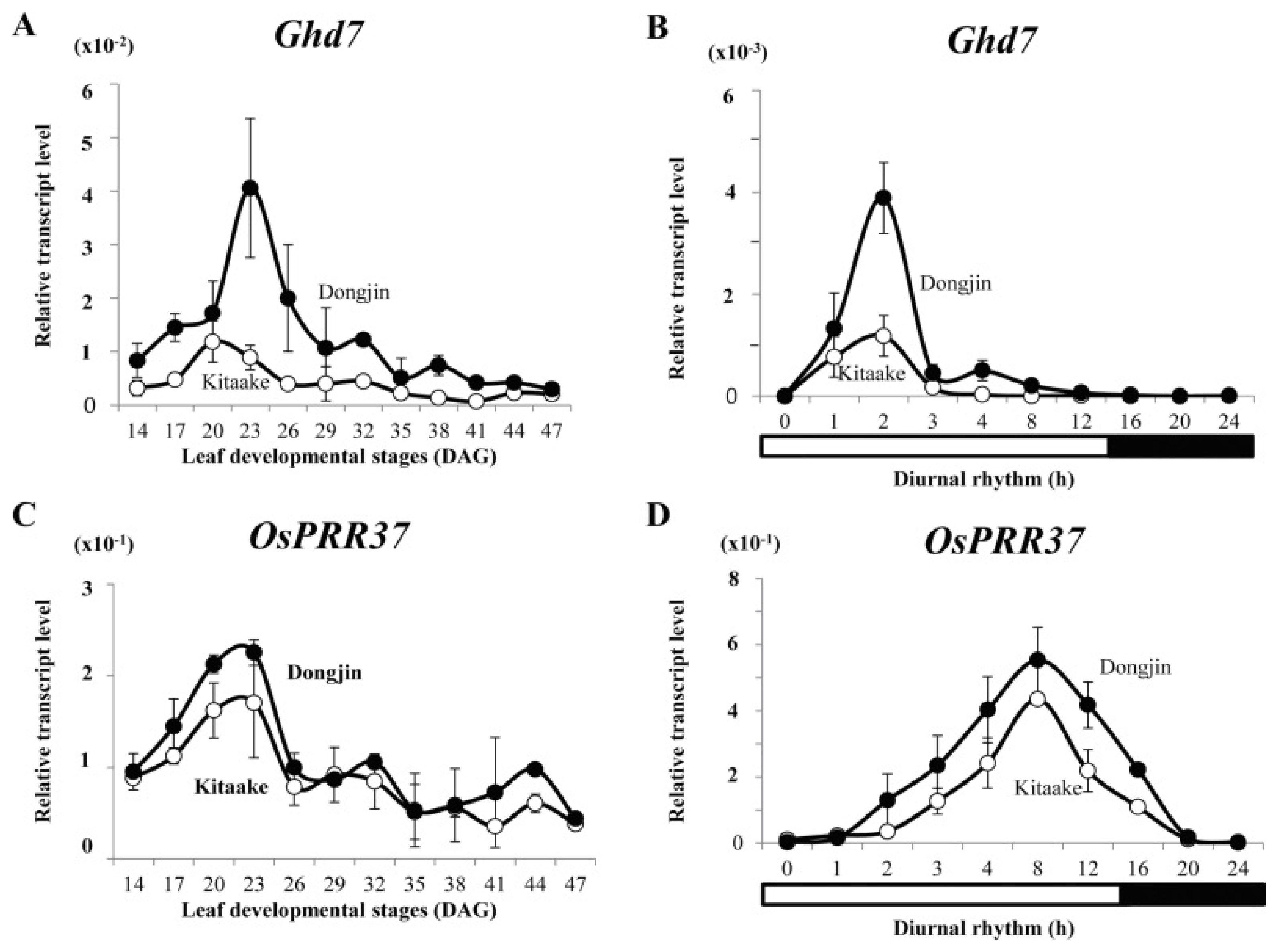
Ghd7 were observed to be much lower in Kitaake than in Dongjin (mid-to late flowering rice cultivar). It was observed that the early flowering phenotype in Kitaake primarily occurs for those having mutations in
Ghd7 and
OsPRR37 (
Fig. 5). Moreover, knocking down
Hd1 showed late flowering under short days and early flowering under long days and Kitaake flowered earlier under both photoperiods (
Kim et al. 2013).
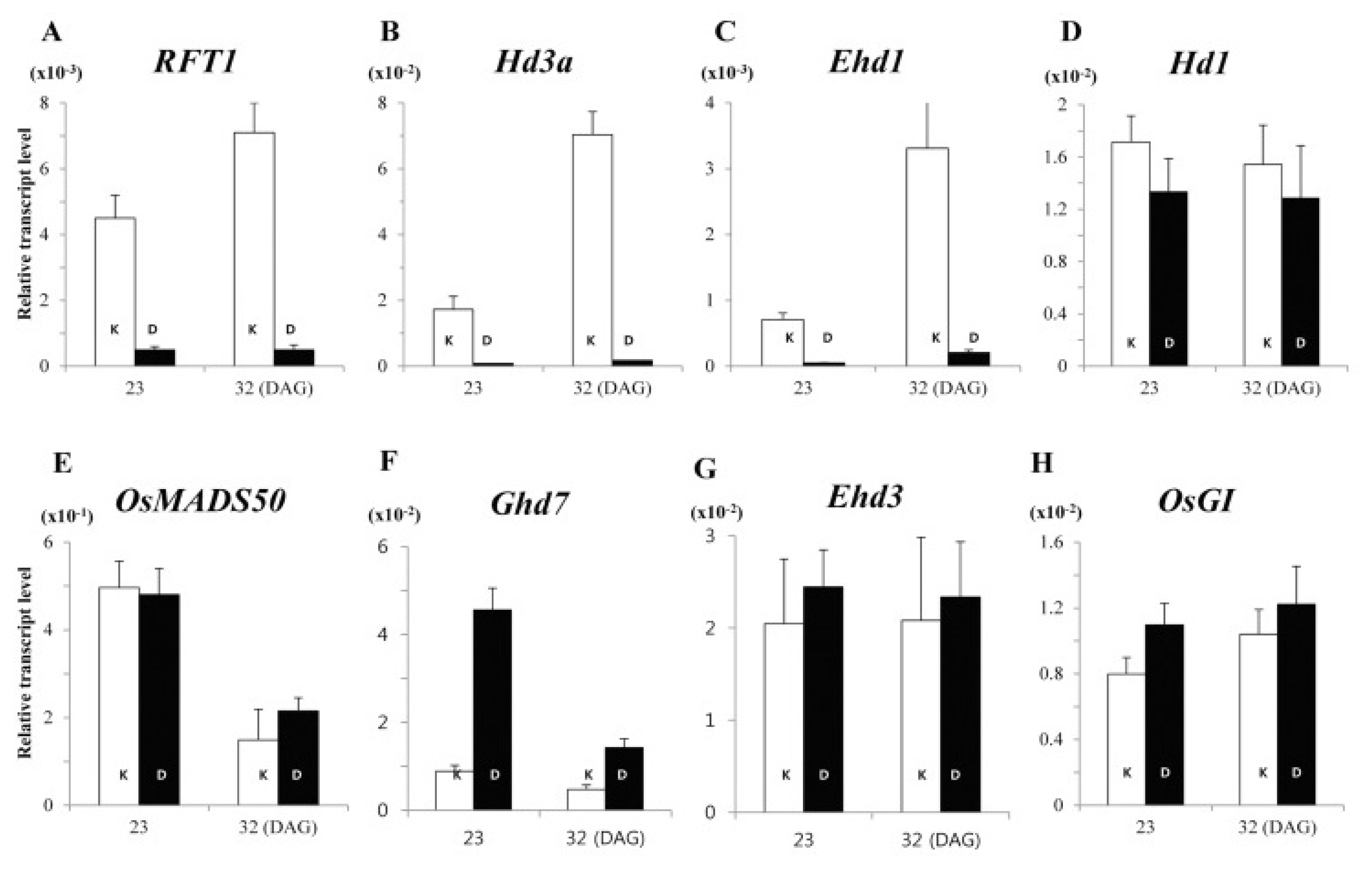
Several varieties might flower under NLD condition despite having LD floral repressor genes, suggesting the occurrence of transcription factors that play a crucial role to reduce the sensitivity to photoperiod. Expression level of
Ehd1 was induced transiently, likewise to
Hd3a and
RFT (
Fig. 6), and in many varieties its expression is high despite the presence of
Ghd7 and/or
Ghd8 wild type alleles.
Gomez-Ariza et al. (2015) reported that the expression of other LD repressors was not reduced seconding the idea that other supplementary factors influence flowering and photosynthesis by repression of
Hd1 and promotion of
Ehd1 expression.
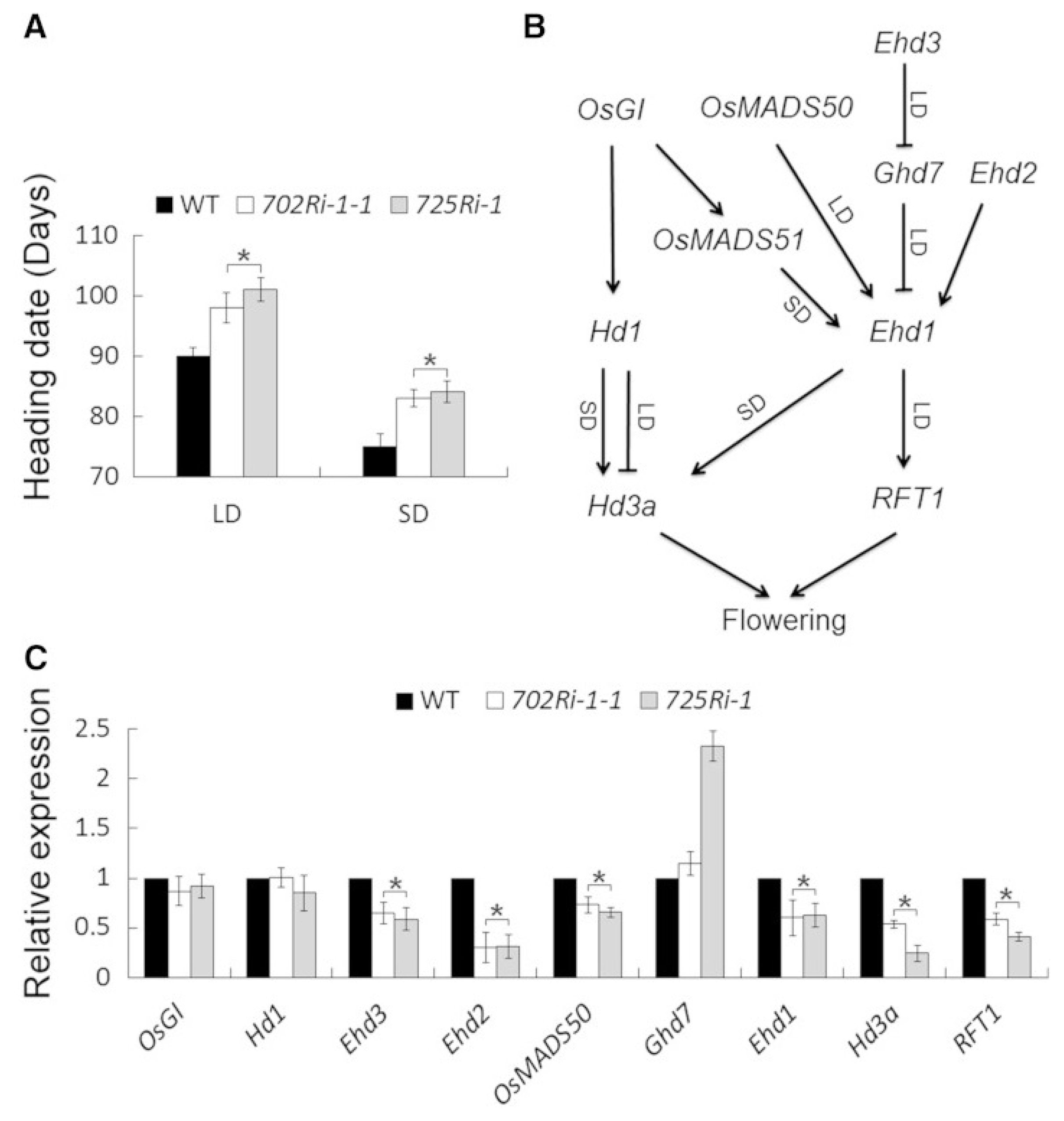
MORF-RELATED GENE702, a Reader Protein (MRG702) plays a primary function in rice growth and development. The rice genome contains two closely associated homologs of Arabidopsis MRG gene: MRG701 and MRG702 (
Xu et al. 2014;
Jin et al. 2015). Some monocots, like corn (
Zea mays), sorghum (
Sorghum bicolor) and wheat (
Triticum aestivum), have only one MRG protein, whilst rice has both. The MRG702 mutants exhibits late-flowering phenotype.
Jin et al. (2015) observed that when MRG702 mutant (702Ri-1-1) and wild-type plants were grown side by side at two locations with different latitudes in China (Shanghai: 31° 0′ 18″ N / 121° 24′ 31″ E under LD conditions, and Sanya: 18° 15′ 12″ N / 109° 30′ 13″ E under SD conditions), the mutant 702Ri-1-1 plants showed a delayed flowering phenotype (
Fig. 7). The heading dates of 702Ri-1-1 mutants were at 95 to 102 days in Shanghai and 81 to 86 days in Sanya, which indicated that MRG702 is likely to be involved in hastening flowering in rice.
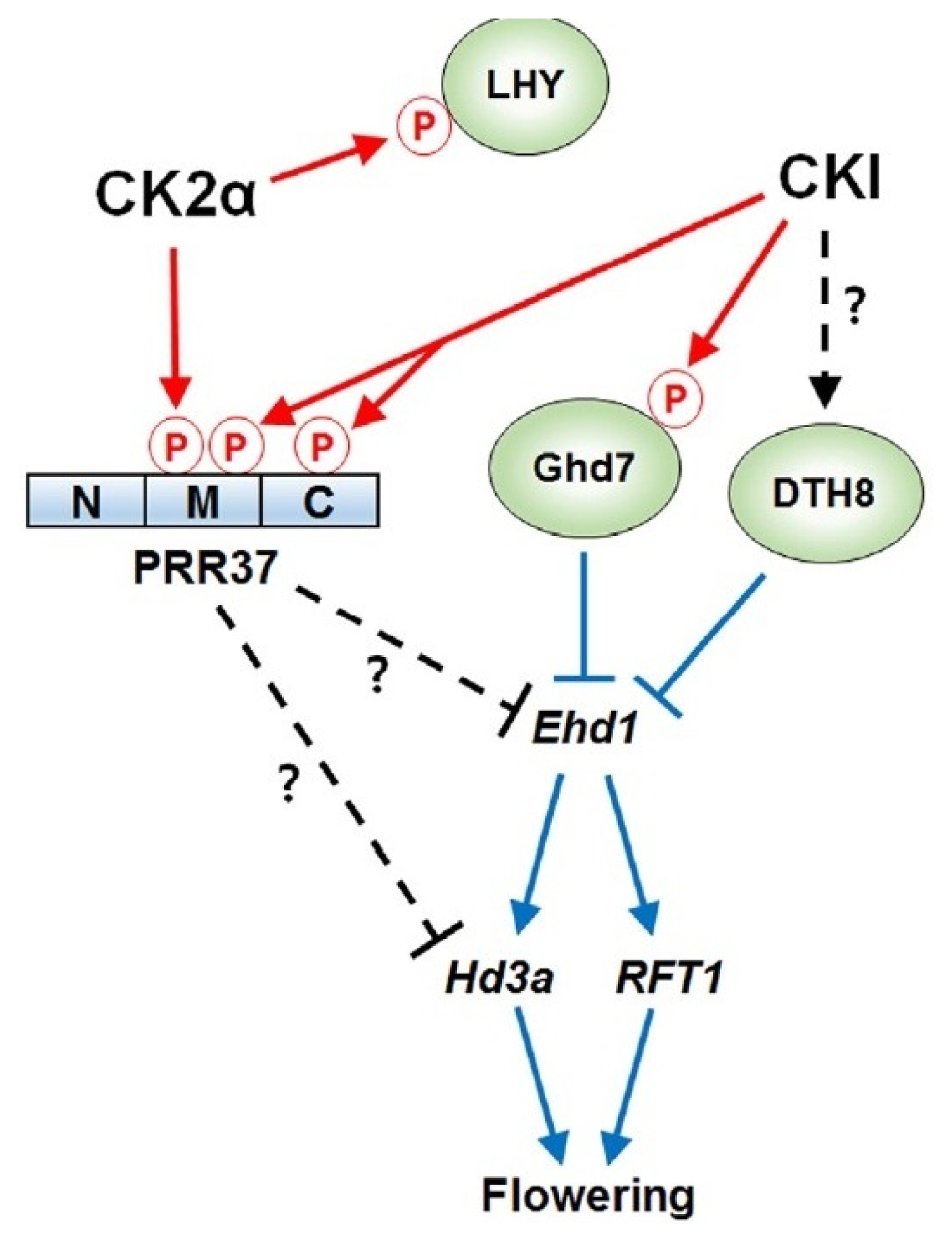
In rice, the Casein kinases CKI and CK2α directly modify Pseudo-Response Regulator 37 (PRR37) at the post-translational level. In non-inductive LD conditions for floral repression Hd6/CK2α phosphorylates downstream LD-dependent floral repressor(s). Many researchers have reported the epistatic interactions between
Hd2/PRR37 and other flowering-time controllers in rice. Firstly, genetic studies revealed that
Hd6 acts by upregulating
Hd2 to a late flowering time, because
hd2 is epistatic to
Hd6 or
hd6 in LD conditions (
Yamamoto et al. 2000). Nonetheless, they augured that CK2α regulates PRR37 at the post-translational level, because CK2α interacts with and phosphorylates PRR37.
Ogiso et al. (2010) observed that under LD conditions, floral repression of
Hd6 requires functional
Hd1 because
hd1 is epistatic to
Hd6 or
hd6 in LD conditions, but CK2α does not associate with or phosphorylate Hd1. Therefore, they theorized that Hd1 activity is regulated by an anonymous regulatory protein phosphorylated by CK2α; besides that, PRR37 is a vital candidate for this unknown protein.
Hd1 genetically acts downstream of
Hd2 to late flowering time in LD conditions (
Lin et al. 2000) but no work has been published on the direct linkage between these two proteins, Hd1 and PRR37 of CCT domain, at the transcript or post-translation level.
Hd16 also acts as an LD-mediated floral repressor by down regulating
Ehd1 expression (
Hori et al. 2013;
Kwon et al. 2014).
Hori et al. (2013) used
in vitro kinase enzyme assays to reveal that CKI interacts with and phosphorylates the LD-dependent floral inhibitor Ghd7, but does not phosphorylate Hd1. This suggests that CKI downregulates
Ehd1 expression by upregulating Ghd7 activity at the post-translational level. Likewise to
Hd6,
Hd16 requires functional
Hd2 to delay flowering in LD conditions (
Yamamoto et al. 2000;
Shibaya et al. 2011;
Hori et al. 2013), indicating that
Hd6,
Hd16, and
Hd2 function in the same floral repression genetic pathway of LD condition. In fact, they observed that CKI also interacts with and phosphorylates PRR37. These results indicate that the post-translational changes of PRR37 by CKI and CK2α likewise affects its activity and stability, which needs to be estimated
in vivo (
Kwon et al. 2015). It is remarkable that CKI and CK2α phosphorylate different locations in the PRR37 protein; CKI phosphorylates the recombinant partial proteins PRR37m and PRR37c, but CK2α phosphorylates only PRR37m (
Fig. 8). This implies that CKI and CK2α mediated phosphorylation levels independently regulate the activity and/or stability of PRR37. Further
in vitro casein kinase assays along with site-specific mutagenesis of PRR37 is necessary to detect the exact sites where CKI and CK2α phosphorylate PRR37. Natural variations in
Hd1,
PRR37,
Ghd7,
DTH8,
Hd6, and
Hd16 were found in different rice varieties grown widely in Asia and Europe, and their variants play significant roles in the downstream of
Ehd1 expression to delay flowering time in natural LD conditions. Hd16/CKI impedes flowering in the
Ehd1-concerned pathway through phosphorylation of Ghd7 (
Hori et al. 2013) as well as PRR37. Nonetheless, the relationship between
PRR37 and
Ehd1 remains obscure, based on two contradictory reports (
Koo et al. 2013;
Yan et al. 2013). Hd6/CK2α might phosphorylate PRR37 to downstream
Hd3a and
RFT1 expression in the
Hd1-concerned pathway (
Ogiso et al. 2010) by anonymous mechanisms. Aside from this, CK2α phosphorylates OsLHY
in vitro, though Hd6/CK2α is unassociated with the circadian rhythm in rice (
Ogiso et al. 2010). The LD-mediated repression of flowering by
Ghd7 and
PRR37 is genetically additive (
Kim et al. 2013;
Koo et al. 2013), revealing that both work independently and synergistically.
FUTURE OUTLOOK AND RESEARCH THRUST
Rice plant produces limited number of flowers simultaneously and the total quantity of flowers is closely associated with the length of the vegetative phase. That is why, genetic and epigenetic mediation of the vegetative period is important in estimating yield of rice. CKI and CK2α are likely to contribute to enhance the photoperiod sensitivity of rice through phosphorylation of PRR37. Therefore, further investigation on CKI and/or CK2α-regulated phosphorylation of PRR proteins is necessary. Furthermore, the significance of pyramiding of floral repressors is yet to be determined. Varieties bearing combinations of different flower regulating genes are necessary for development of climate smart rice genotypes to combat fragile environmental conditions (
Lin et al. 2000). Hd1 activity is thought to be regulated by an unknown controller protein phosphorylated by CK2α and postulated that PRR37 is a principal candidate for this unknown protein. Genetically Hd1 acts downstream of Hd2 to delay flowering time in LD but no research has been undertaken on the aspect of direct linkage between these two CCT domain located proteins, Hd1 and PRR37, at the transcript or post translation level. It is argued that the post-translational changes of PRR37 by CKI and CK2α likewise affect its activity and stability, which needs to be estimated
in vivo (
Kwon et al. 2015). It is mentionable that CKI and CK2α phosphorylate different domain locations in the PRR37 protein; CKI phosphorylates the recombinant partial proteins PRR37m and PRR37c, but CK2α phosphorylates only PRR37m (
Kwon et al. 2015). This implies that CKI and CK2α mediated phosphorylation level regulate the activity and/or stability of PRR37 independently. Further
in vitro enzyme kinase assays along with site-specific mutagenesis of PRR37 are necessary to detect the specific sites for phosphorylation by CKI and CK2α in PRR37. Further incitation into the intricate phenomena of research by examining the biochemical relationships among Hd1, PRR37, and CK2α is important to unravel the molecular mechanism of
Hd6-mediated floral repression under LD conditions. Furthermore, it would be worthwhile to explore whether CK2α, CKI, or both can interact with other OsPRR proteins such as OsTOC1/OsPRR1, OsPRR59, OsPRR73, and OsPRR95 etc. Very recently, a Trithorax group protein, OsTrx1, was found to regulate flowering time in rice via its interaction with
Ehd3 (
Choi et al. 2014), though, OsTrx1 possesses H3K4 methyltransferase activity require to be determined. In addition, molecular and genetic studies are needed to reveal how MRG702 recognizes the histone methylation marks and stimulates gene expression in rice.
Fig. 1Days to heading of rice varieties under three various day length conditions, 10D (
white), 14D (
gray), and 16D (
black) in a growth chamber. Data are mean values and SD (
n = 10). KTm is the advanced backcrossed progeny of Kitaibuki with the functional
Hd5 (
Fujino et al. 2013).
Fig. 2Temporal gene expression patterns of rice panicle development-related genes in SAM regions during early stages of panicle development. Levels of mRNA accumulation were examined by qRT–PCR. Timing of transition was determined by the microscopic observation of SAM parts of test samples for all sampling dates. Day 2 indicates the timing of the start of primary rachis differentiation. The results are the mean ± SE (n = 3 individual plants). Three RT–PCRs were done for each cDNA sample from one plant, mRNAs were made from the respective SAM samples grown under SD conditions. Likewise, data were obtained when normalized by number of SAMs for mRNA preparation (
Endo-Higashi and Izawa 2011).
Fig. 3Correlation of flowering time with
RFT1 and
Hd3a RNA levels of cultivars carrying functional (blue dots) or non-functional Hd1 alleles (red dots). RNA levels were determined by real-time RT-PCR and shown as natural logarithms (
Naranjo et al. 2014).
Fig. 4Chromosomal locations of the heading date QTLs detected in the Koshihikari × Hayamasari BILs under natural field (NF), long day (LD: 14.5 hours), extremely long-day (ELD: 18 hours), and short-day (SD:10 hours) conditions. The lengths of the
rectangles indicate a two-LOD confidence interval for the QTLs. The small horizontal bars and small letters show the positions of the markers and the names of the marker nearest to the LOD peak, respectively. The thick horizontal lines and the circles with italicized names represent the positions of the QTLs for heading date identified in previous studies (
Lin et al. 2003;
Matsubara et al. 2008;
Wei et al. 2010;
Shibaya et al. 2011;
Yan et al. 2011).
Fig. 5Expression profiles of
Ghd7 and
OsPRR37 in cv. Kitaake (open circles) and cv. Dongjin (closed circles) grown under long days. (A and C) Temporal expression patterns of
Ghd7 and
OsPRR37 in leaf blades from 14 to 47 DAG; samples were prepared at 2 h after turning on lights. (B and D) Diurnal rhythm in leaves at 23 DAG. Values are mean of two or more independent experiments and standard deviation. Y-axis, relative values between transcript levels and
Ubi (
Kim et al. 2013).
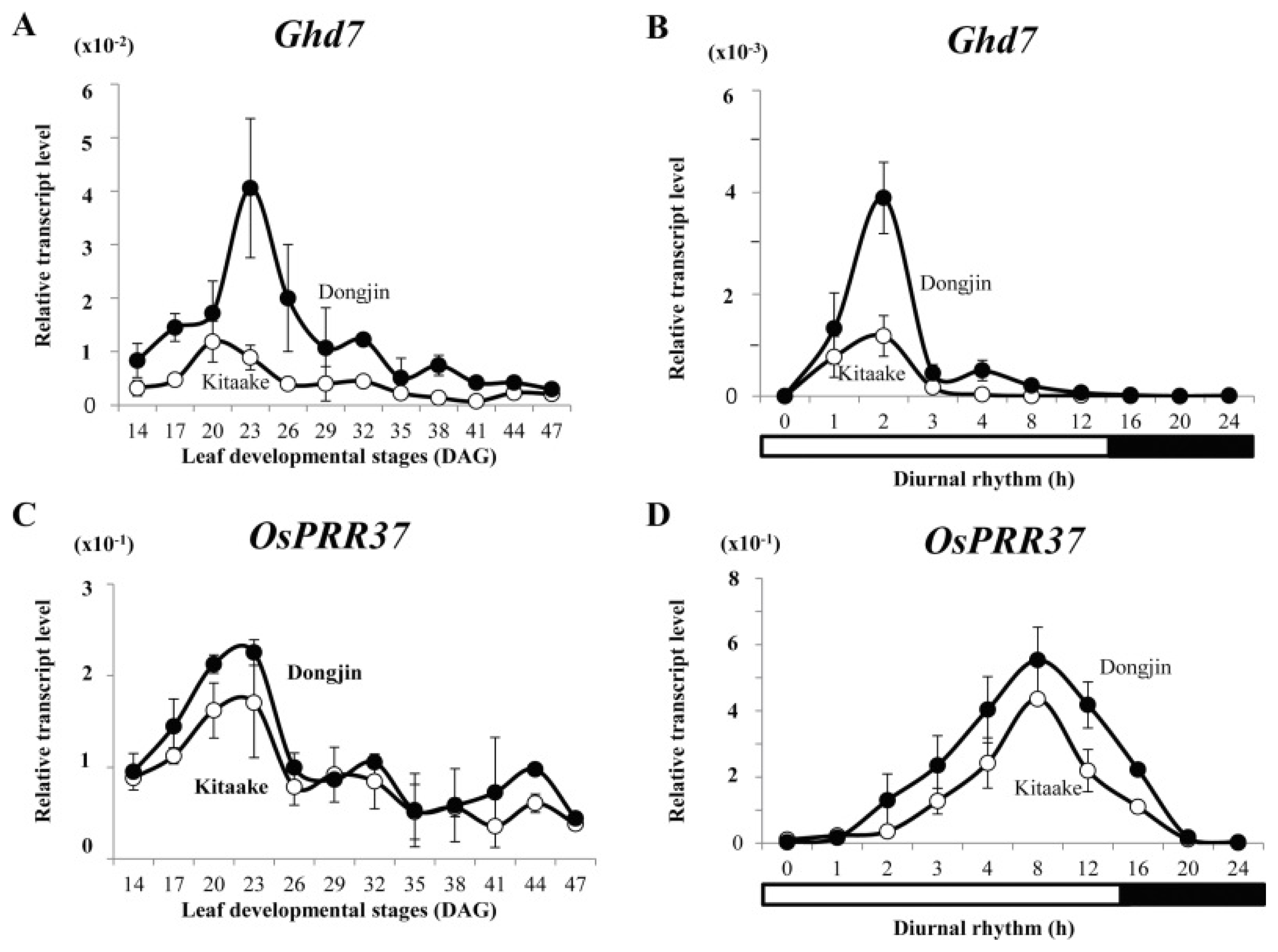
Fig. 6Expression profiles for flowering regulators from cv. Kitaake (K; white) and cv. Dongjin (D; black) at 23 and 32 DAG under long days. RNA was prepared from leaf blades 2 hours after turning on lights. Values are mean of two or more independent experiments with standard deviation. Y-axis, relative values between transcript levels for regulatory gene and
Ubi (
Kim et al. 2013).
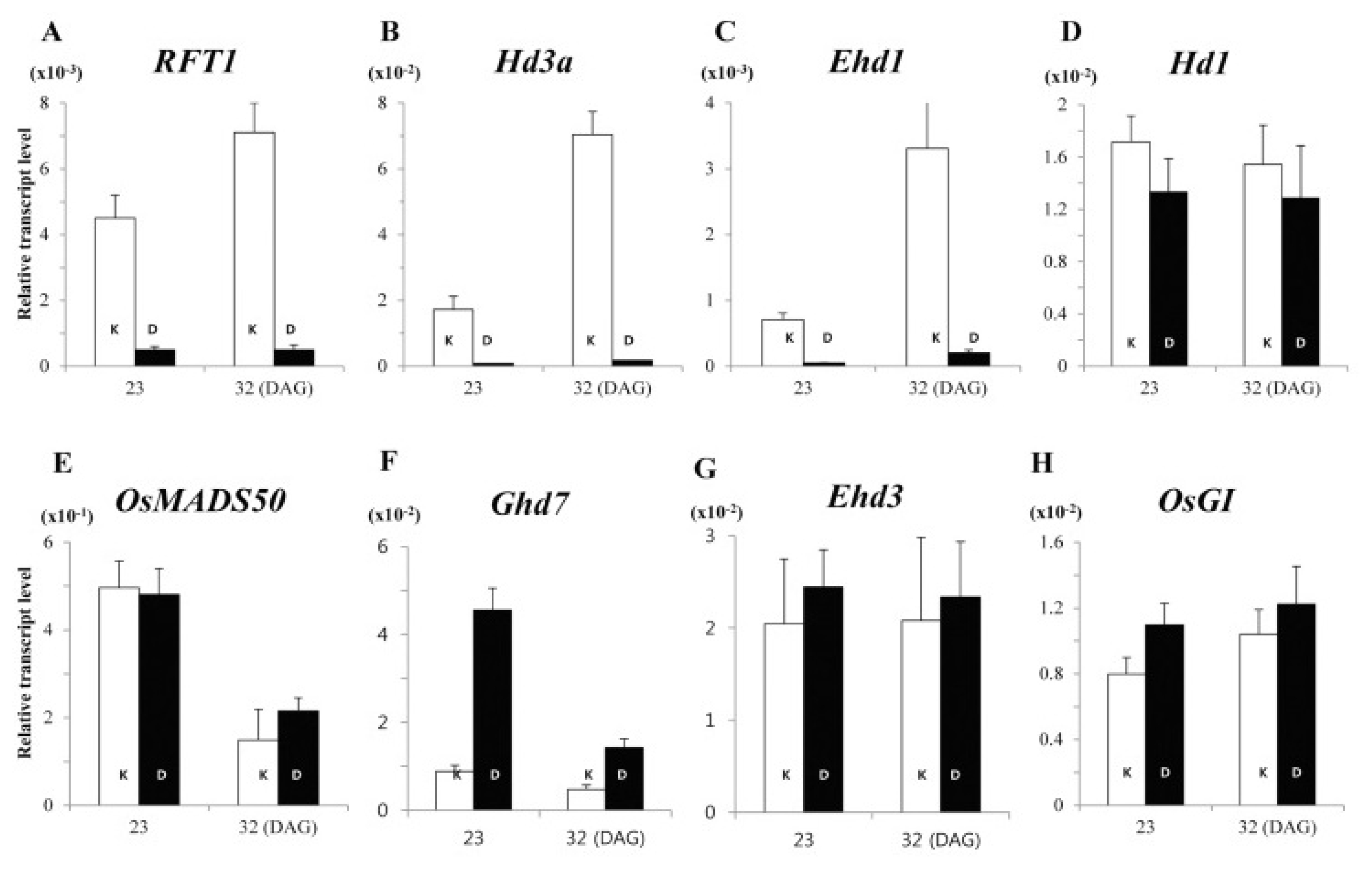
Fig. 7MRG702 promotes flowering in rice. (A) Heading time of wild-type (WT), 702Ri-1-1, and 725Ri-1 plants grown under LD (Shanghai) or SD (Sanya) conditions. Values shown are means 6 SD (n = 30). Asterisks indicate statistically significant differences between the indicated genotypes and the wild type (
P, 0.01). (B) Schematic of two core flowering regulatory pathways in rice. (C) Relative transcript levels of flowering regulatory genes in the indicated plants. Quantitative RT-PCR analyses were performed using leaves collected at 2 hours after dawn from 28-day-old rice seedlings grown in a growth chamber under an LD photoperiod (14 hours of light/10 hours of dark). OsACTIN1 served as the internal control, and fold change relative to the wild-type level is shown. Values shown are means 6 SD from three independent replicates. Asterisks indicate statistically significant differences between the indicated genotypes and the wild type (
P, 0.01) (
Jin et al. 2015).
Fig. 8Schematic model of regulation of PRR37 and other floral integrators by CKI and CK2α, in photoperiodic flowering under long day conditions in rice. Blue and red arrows indicate the regulation at the transcriptional and post-translational levels, respectively (
Kwon et al. 2015).
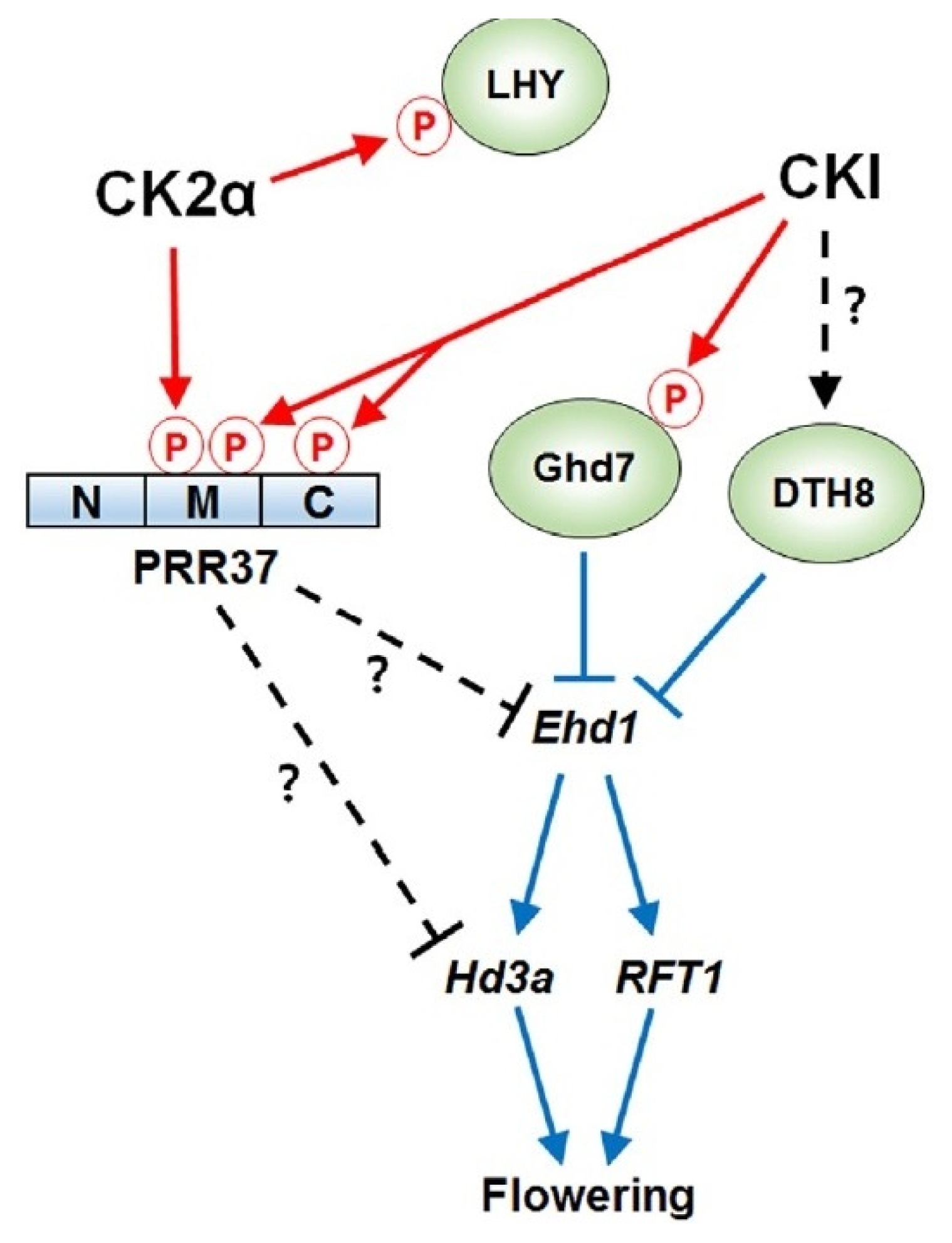
Table 1Major genes involve in flowering time and floral organ development in rice.
Table 1
|
Name and symbol of Genes |
Special traits |
Chr. |
|
phosphatidylinositol 4-phosphate 5-kinase 1(OsPIPK1), GIGANTEA (OsGI), Osmads50 (osmads50), photoperiodic sensitivity 5 (se5), heading date 6 (Hd6), heading date 1(Hd1), EARLY FLOWERING 3-1 (OsELF3-1), NUTRITION RESPONSE AND ROOT GROWTH a (NRRa), NUTRITION RESPONSE AND ROOT GROWTH b (NRRb)
|
Flowering time |
1,3,6 |
|
Heading date 16 (Hd16), RICE FLOWERING LOCUS T 1(RFT1), Pseudo-Response Regulator37 (OsPRR37), CCAAT-box-binding transcription factor (LH8), Trithorax 1 (OsTrx1), embryonic flower 2b (OsEMF2b), semi-dwarf and late flowering (DIF1), RICE FLOWERING LOCUS T 1 (RFT1)
|
Flowering time under long day condition. |
1,3,6,7,8,9 |
|
grain number, plant height and heading date 7(Ghd7), days to heading on chromosome 8 (DTH8), DNA-binding with one finger12 (OsDof12), Heading date 17 (Hd17)
|
Flowering time under short day condition. |
2,3,6, 7,8 |
|
early heading date 3 (ehd3), G-box factor 14-3-3c protein (GF14c), OsMADS51 (OsMADS51), heading date 3a (Hd3a), CONSTANS-LIKE 3 (OsCO3), days to heading on chromosome 3 (dth3), Early heading date 1(Ehd1)
|
Flowering time independent of day length. |
1,3,6, 8,9, 10 |
|
“grain number, plant height and heading date 8” (Ghd8)
|
Flowering time and Transition from vegetative to reproductive phase. |
8 |
|
SPL11-interacting protein1(SPIN1)
|
Flowering time in hybrid Chromatin modification. |
3 |
|
early flowering 7 (ef7)
|
Grain length and width, 1000-grain weight, flowering time. |
6 |
|
VIN3-LIKE 2 (OsVIL2)
|
Flowering time, tiller growth, panicle branching. |
2 |
|
peter pan syndrome (pps), Rice Indeterminate 1(rid1)
|
Floral organ identity, flowering time. |
2,10 |
|
CONSTANS-like 4 (OsCOL4), cryptochrome2(Oscry2), histone deacetylase 1(OsHDT1)
|
Flowering time, deetiolation response, sensitivity to red and far-red light. |
2,5 |
|
rice Dof daily fluctuations 1 (Rdd1), Rice FLO-LFY homolog (RFL), leafy hull sterile1(lhs1)
|
Flowering time, gibberellin and ethylene sensitivity. |
1 |
|
NECK LEAF 1 (nl1)
|
Drought and salinity tolerance, tillering. flowering time, auxin sensitivity. |
5 |
|
phytochromeB (phyB)
|
Flowering time, grain number, grain size. |
3 |
|
phytochromeA (phyA)
|
Flowering time, grain number. |
3 |
|
phytochromeC (phyC)
|
Negative regulator of heading |
3 |
|
Earlier flowering1(ef1)
|
Slower heading and flowering |
3 |
|
ETHYLENE RESPONSE2 (etr2)
|
Delay flowering through the repression of RFT1 by red-light signal mediated H3K4me3 demethylation under long day-length conditions |
4 |
|
CONSTITUTIVE TRIPLE-RESPONSE2 (OsCTR2)
|
Heading date |
2 |
|
OsmiR393 (OsmiR393), pyruvate dehydrogenase kinase (pdhk), early heading date 4 (Ehd4), autophagy associated gene 7 (OsATG7)
|
Flowering time, dwarfism, fertility, germination rate. |
1,5 |
References
- Berr A, Shafiq S, Shen WH. 2011. Histone modifications in transcriptional activation during plant development. Biochim Biophys Acta. 1809: 567-576.
- Choi SC, Lee S, Kim SR, Lee YS, Liu C, Cao X, et al. 2014. Trithorax group protein Oryza sativa Trithorax1 controls flowering time in rice via interaction with early heading date3. Plant Physiol. 164: 1326-1337.
- Conti L, Bradley D. 2007. TERMINAL FLOWER1 is a mobile signal controlling Arabidopsis architecture. Plant Cell. 19: 767-778.
- Doi K, Izawa T, Fuse T, Yamanouchi U, Kubo T, Shimatani Z, Yano M, et al. 2004. Ehd1, a B-type response regulator in rice, confers short-day promotion of flowering and controls FT-like gene expression independently of Hd1. Genes Dev. 18: 926-936.
- Ebana K, Shibaya T, Wu J, Matsubara K, Kanamori H, Yamane H, et al. 2011. Uncovering of major genetic factors generating naturally occurring variation in heading date among Asian rice cultivars. Theor Appl Genet. 122: 1199-121.
- Endo-Higashi N, Izawa T. 2011. Flowering time genes heading date 1 and early heading date 1 together control panicle development in rice. Plant Cell Physiol. 52: 1083-1094.
- Fujino K, Iwata N. 2011. Identification of QTLs controlling heading date on the short arm of chromosome 3 in rice (Oryza sativa L.). Breed Sci. 61: 294-300.
- Fujino K, Yamanouchi U, Yano M. 2013. Roles of the Hd5 gene controlling heading date for adaptation to the northern limits of rice cultivation. Theor Appl Genet. 126: 611-618.
- Gómez-Ariza J, Galbiati F, Goretti D, Brambilla V, Shrestha R, Pappolla A, et al. 2015. Loss of floral repressor function adapts rice to higher latitudes in Europe. J Exp Bot. 66: 2027-2039.
- Griffiths S, Dunford RP, Coupland G, Laurie DA. 2003. The evolution of CONSTANS-like gene families in barley, rice, and Arabidopsis. Plant Physiol. 131: 1855-1867.
- Hori K, Matsubara K, Yana M. 2016. Genetic control of flowering time in rice: integration of Mendelian genetics and genomics. Theor Appl Genet. 129: 2241-2252.
- Hori K, Ogiso-Tanaka E, Matsubara K, Yamanouchi U, Ebana K, Yano M. 2013. Hd16, a gene for casein kinase I, is involved in the control of rice flowering time by modulating the day-length response. Plant J. 76: 36-46.
- Inoue H, Tanisaka T, Okumoto Y, Yamagata H. 1992. An early-heading mutant gene of a mutant line HS66 of rice (in Japanese with English summary). Kinki Journal of Crop Science and Breeding. 37: 47-52.
- Ishikawa R, Tamaki S, Yokoi S, Inagaki N, Shinomura T, Takano M, et al. 2005. Suppression of the floral activator gene Hd3a is the principal cause of the night break effect in rice. Plant Cell. 17: 3326-3336.
- Itoh H, Nonoue Y, Yano M, Izawa T. 2010. A pair of floral regulators sets critical day length for Hd3a florigen expression in rice. Nat Genet. 42: 635-638.
- Izawa T, Oikawa T, Tokutomi S, Okuno K, Shimamoto K. 2000. Phytochromes confer the photoperiodic control of flowering in rice (a short-day plant). Plant J. 22: 391-399.
- Jin J, Shi J, Liu B, Liu Y, Huang Y, Yu Y, et al. 2015. MORF-RELATED GENE702, a reader protein of trimethylated Histone H3 Lysine 4 and Histone H3 Lysine 36, is involved in brassinosteroid-regulated growth and flowering time control in rice. Plant Physiol. 168: 1275-1285.
- Kim SL, Choi M, Jung K-H, An G. 2013. Analysis of the early-flowering mechanisms and generation of T-DNA tagging lines in Kitaake, a model rice cultivar. J Exp Bot. 64: 4169-4182.
- Knippschild U, Gocht A, Wolff S, Huber N, Lohler J, Stoter M. 2005. The casein kinase 1 family: participation in multiple cellular processes in eukaryotes. Cell Signal. 17: 675-689.
- Kojima S, Takahashi Y, Kobayashi Y, Monna L, Sasaki T, Araki T, et al. 2002. Hd3a, a rice ortholog of the Arabidopsis FT gene, promotes transition to flowering downstream of Hd1 under short-day conditions. Plant Cell Physiol. 43: 1096-1105.
- Koo BH, Yoo SC, Park JW, Kwon CT, Lee BD, An G, et al. 2013. Natural variation in OsPRR37 regulates heading date and contributes to rice cultivation at a wide range of latitudes. Mol Plant. 6: 1877-1888.
- Kwon C, Koo B, Kim D, Yoo S, Paek N. 2015. Casein kinases I and 2α phosphorylate Oryza sativa Pseudo-Response Regulator 37 (OsPRR37) in photoperiodic flowering in rice. Mol Cells. 38: 81-88.
- Kwon CT, Yoo SC, Koo BH, Cho SH, Park JW, Zhang ZY, et al. 2014. Natural variation in Early flowering1 contributes to early flowering in japonica rice under long days. Plant Cell Environ. 37: 101-112.
- Lee S, Kim J, Han JJ, Han MJ, An G. 2004. Functional analyses of the flowering time gene OsMADS50, the putative SUPPRESSOR OF OVEREXPRESSION OF CO 1/AGAMOUS-LIKE 20 (SOC1/AGL20) ortholog in rice. Plant J. 38: 754-764.
- Lin HX, Liang ZW, Sasaki T, Yano M. 2003. Fine mapping and characterization of quantitative trait loci Hd4 and Hd5 controlling heading date in rice. Breed Sci. 53: 51-59.
- Lin HX, Yamamoto T, Sasaki T, Yano M. 2000. Characterization and detection of epistatic interactions of three QTLs, Hd1, Hd2 and Hd3, controlling heading date in rice using nearly isogenic lines. Theor Appl Genet. 101: 1021-1028.
- Liu C, Lu F, Cui X, Cao X. 2010. Histone methylation in higher plants. Annu Rev Plant Bio. 61: 395-420.
- Maas LF, McClung A, McCouch S. 2010. Dissection of a QTL reveals an adaptive, interacting gene complex associated with transgressive variation for flowering time in rice. Theor Appl Genet. 120: 895-908.
- Matsubara K, Kono I, Hori K, Nonoue Y, Ono N, Shomura A, et al. 2008. Novel QTLs for photoperiodic flowering revealed by using reciprocal backcross inbred lines from crosses between japonica rice cultivars. Theor Appl Genet. 117: 935-945.
- Matsubara K, Ogiso-Tanaka E, Hori K, Ebana K, Ando T, Yano M. 2012. Natural variation in Hd17, a homolog of Arabidopsis ELF3 that is involved in rice photoperiodic flowering. Plant Cell Physiol. 53: 709-716.
- Miura K, Ashikari M, Matsuoka M. 2011. The role of QTLs in the breeding of high-yielding rice. Trends Plant Sci. 16: 319-326.
- Mulekar JJ, Huq E. 2014. Expanding roles of protein kinase CK2 in regulating plant growth and development. J Exp Bot. 65: 2883-2893.
- Naranjo L, Talón M, Domingo C. 2014. Diversity of floral regulatory genes of japonica rice cultivated at northern latitudes. BMC Genomics. 15: 101-107.
- Nonoue Y, Fujino K, Hirayama Y, Yamanouchi U, Lin S, Yano M. 2008. Detection of quantitative trait loci controlling extremely early heading in rice. Theor Appl Genet. 116: 715-722.
- Ogiso E, Takahashi Y, Sasaki T, Yano M, Izawa T. 2010. The role of Casein kinase II in flowering time regulation has diversified during evolution. Plant Physiol. 152: 808-820.
- Robson F, Costa MMR, Hepworth SR, Vizir I, Pineiro M, Reeves PH, et al. 2001. Functional importance of conserved domains in the flowering-time gene CONSTANS demonstrated by analysis of mutant alleles and transgenic plants. Plant J. 28: 619-631.
- Saito H, Ogiso-Tanaka E, Okumoto Y, Yoshitake Y, Izumi H, Yokoo T, et al. 2012. Ef7 encodes an ELF3-like protein and promotes rice flowering by negatively regulating the floral repressor gene Ghd7 under both short- and long-day conditions. Plant Cell Physiol. 53: 717-728.
- Shibaya T, Yasunori N, Nozomi O, Utako Y, Kiyosumi H, Masahiro Y. 2011. Genetic interactions involved in the inhibition of heading by heading date QTL, Hd2 in rice under long-day conditions. Theor Appl Genet. 123: 1133-1143.
- Sun C, Fang J, Zhao T, Xu B, Zhang F, Liu L, et al. 2012. The histone methyltransferase SDG724 mediates H3K36me2/3 deposition at MADS50 and RFT1 and promotes flowering in rice. Plant Cell. 24: 3235-3247.
- Takahashi Y, Shomura A, Sasaki T, Yano M. 2001. Hd6, a rice quantitative trait locus involved in photoperiod sensitivity, encodes the subunit of protein kinase CK2. Proc Natl Acad Sci USA. 98: 7922-7927.
- Takahashi Y, Teshima KM, Yokoi S, Innan H, Shimamoto K. 2009. Variations in Hd1 proteins, Hd3a promoters, and Ehd1 expression levels contribute to diversity of flowering time in cultivated rice. Proc Natl Acad Sci USA. 106: 4555-4560.
- Tan J, Wu F, Wan J. 2017. Flowering time regulation by the CONSTANS-Like gene OsCOL10. Plant Signal Behav. 12: e1267893
- Tsuji H, Taoka K, Shimamoto K. 2011. Regulation of flowering in rice: two florigen genes, a complex gene network, and natural variation. Curr Opin Plant Biol. 14: 45-52.
- Uga Y, Nonoue Y, Liang ZW, Lin HX, Yamamoto S, Yamanouchi U, et al. 2007. Accumulation of additive effects generates a strong photoperiod sensitivity in the extremely late-heading rice cultivar ‘Nona Bokra’. Theor Appl Genet. 114: 1457-1466.
- Wei X, Xu J, Guo H, Jiang L, Chen S, Yu C, et al. 2010. DTH8 suppresses flowering in rice, influencing plant height and yield potential simultaneously. Plant Physiol. 153: 1747-1758.
- Xu Y, Gan ES, Zhou J, Wee WY, Zhang X, Ito T. 2014. Arabidopsis MRG domain proteins bridge two histone modifications to elevate expression of flowering genes. Nucleic Acids Res. 42: 10960-10974.
- Xue WY, Xing YZ, Weng XY, Zhao Y, Tang WJ, Wang L, et al. 2008. Natural variation in Ghd7 is an important regulator of heading date and yield potential in rice. Nat Genet. 40: 761-767.
- Yamamoto T, Lin H, Sasaki T, Yano M. 2000. Identification of heading date quantitative trait locus Hd6 and characterization of its epistatic interactions with Hd2 in rice using advanced backcross progeny. Genetics. 154: 885-891.
- Yan WH, Liu HY, Zhou XC, Li QP, Zhang J, Lu L, et al. 2013. Natural variation in Ghd7.1 plays an important role in grain yield and adaptation in rice. Cell Res. 23: 969-971.
- Yan WH, Wang P, Chen HX, Zhou HJ, Li QP, Wang CR, et al. 2011. A Major QTL, Ghd8, plays pleiotropic roles in regulating grain productivity, plant height, and heading date in rice. Mol Plant. 4: 319-330.
- Yano M, Katayose Y, Ashikari M, Yamanouchi U, Monna L, Fuse T, et al. 2000. Hd1, a major photoperiod sensitivity quantitative trait locus in rice, is closely related to the Arabidopsis flowering time gene CONSTANS. Plant Cell. 12: 2473-2483.
- Yun M, Wu J, Workman JL, Li B. 2011. Readers of histone modifications. Cell Res. 21: 564-578.
- Zhang Z, Hu W, Shen G, Liu H, Hu Y, Zhou X, et al. 2017. Alternative functions of Hd1 in repressing or promoting heading are determined by Ghd7 status under long-day conditions. Sci Rep. 7: 5388
- Zhao J, Chen H, Ren D, Tang H, Qiu R, Feng J, et al. 2015. Genetic interactions between diverged alleles of Early heading date 1 (Ehd1) and Heading date 3a (Hd3a)/RICE FLOWERING LOCUS T1 (RFT1) control differential heading and contribute to regional adaptation in rice (Oryza sativa). New Phytol. 208: 936-948.
- Zhao J, Huang X, Ouyang X, Chen W, Du A, Zhu L, et al. 2012. OsELF3-1, an ortholog of Arabidopsis early flowering 3, regulates rice circadian rhythm and photoperiodic flowering. PLoS One. 7: e43705
- Zhu Y, Fan Y, Wang K, Huang D, Liu W, Ying J, et al. 2017. Rice Flowering Locus T 1 plays an important role in heading date influencing yield traits in rice. Sci Rep. 7: 4918