Abstract
It is essential to develop tomato (Solanum lycopersicum L.) cultivars with tolerance to low temperature (LT) for reducing production cost and increasing fruit quality in winter. This study was conducted to investigate the effects of LT on 40 tomato accessions for establishing selection criteria to select accessions with LT tolerance. Tomato plants were grown in two polyethylene film greenhouses with night temperature set-points of 10℃ and 15℃. LT significantly affected the photosynthetic parameters as well as both vegetative and reproductive growth in tomato. There was a significant difference in plant height between plants grown in 10℃ and 15℃. Leaf length and width were also significantly lower in 10℃. The stem diameter was generally lower in 10℃ but 12 accessions in 10℃ had thicker stem diameter than those in 15℃. The retarded growth was due to lower photosynthetic rate in 10℃ than 15℃. Significant delay in flowering was observed among tomato accessions in 10℃. For the number of flowers, 62.5% showed the increase but 37.5% did the decrease in 10℃ among 40 tomato accessions, showing genotype specific interaction with LT. Accessions with small fruits showed reduced fruit set in 10℃; however, it was not obvious among accessions with large fruits. Interestingly, 75% of accessions showed increased chlorophyll contents in 10℃ compared to 15℃. An accession ‘VI037163’ was selected for LT tolerance based on good performance in six out of seven traits considered such as plant height, leaf length, stem diameter, chlorophyll contents, days to flowering, the number of flowers and fruit set.
-
Key words: Tomato, Breeding, Low temperature during the night, Flowering, Chlorophyll content, Photosynthesis
INTRODUCTION
Tomato (
Solanum lycopersicum L.) is one of important greenhouse vegetables grown all-year-round and its cultivation area and production in 2017 were 5,782 ha and 355,107 metric tones, respectively, in Korea (Korean Statistical Information Service, http://kosis.kr). Greenhouses with heating system are mandatory in the winter for tomato cultivation in Korea and the heating cost is the most important factor affecting the production cost of tomato fruits. Approximately 16% of energy cost can be reduced if temperature set-point is reduced by 2℃ in greenhouses (
Elings et al. 2005). Therefore, developing tomato cultivars with LT tolerance is crucial for reducing production cost and increasing fruit quality in winter.
Night temperature in greenhouses is more important to reduce energy cost since ambient temperature is generally lower in the night than day. Besides, high temperature during the daytime can compensate for low temperature during the night (LTN), in terms of the growth of tomato plants (
De Koning 1988). The reduction in energy cost can be achieved by using covering materials such as thermal screens but the best way is simply to decrease the night temperature in greenhouses. Therefore, breeding program for tomato cultivars with LT tolerance should focus on the night temperature.
In order to establish the optimal selection criteria in tomato breeding program for LT tolerance, it is important to investigate the response of tomato accessions to LT in large scale. Several studies investigated the effect of the LT on the growth and yield of tomato; however, most of studies were conducted with less than five cultivars and only few studies with more than ten cultivars (Der Van Ploeg and Heuvelink 2005). This study was conducted to evaluate the effect of LTN on 40 tomato accessions in greenhouses with different temperature set-points for establishing optimal selection criteria to select accessions with LT tolerance.
MATERIALS AND METHODS
Plant materials and growing condition
Thirty tomato breeding lines from National Institute of Horticultural and Herbal Science (NIHHS) and 10 commercial cultivars were used in this study (
Table 1). All the accessions were classified according to fruit size: Wild (> 10 g), Cherry (10-30 g), Medium (31 g-80 g) and Large (< 81 g). Seeds were sown in plastic trays containing 1:1 sand: peat (volume) on October 10, 2018 and grown in a glasshouse with 26/18℃ (day/night) temperature. Seedlings about 20-25 cm in height and with the first truss were transplanted on 8 November 2018 and transferred to two plastic film greenhouses where night air temperature set-point was 10℃ and 15℃. A total of 160 plants (4 plants per accession) per greenhouse were planted with 40 × 40 cm intervals between plants.
Plant height, leaf length and width, and stem diameter were measured from three plants per accession in both greenhouses 30, 65, 95 and 125 days after transplanting (DAT). Days to flower were determined by the date when the third flower on the second truss flowered. Fruit set was determined calculating the ratio of fruits to flowers on the average of second and third trusses. Chlorophyll content was measured from mid leaves using SPAD meter (Konica Minolta, Japan) in 30, 65, 95 and 125 DAT. Differences in all growth parameters between plants grown in 10℃ and 15℃ greenhouses, except for fruit set, were calculated by (L/N)*100, where L and N represent the values of growth parameters in 10℃ and 15℃, respectively. Difference in fruit set was calculated by subtracting fruit set of 15℃ from 10℃. Standard errors were calculated using EXCEL 2016 program (Microsoft Co. Ltd., USA).
Data collection on temperature, relative humidity and photosynthetic parameters
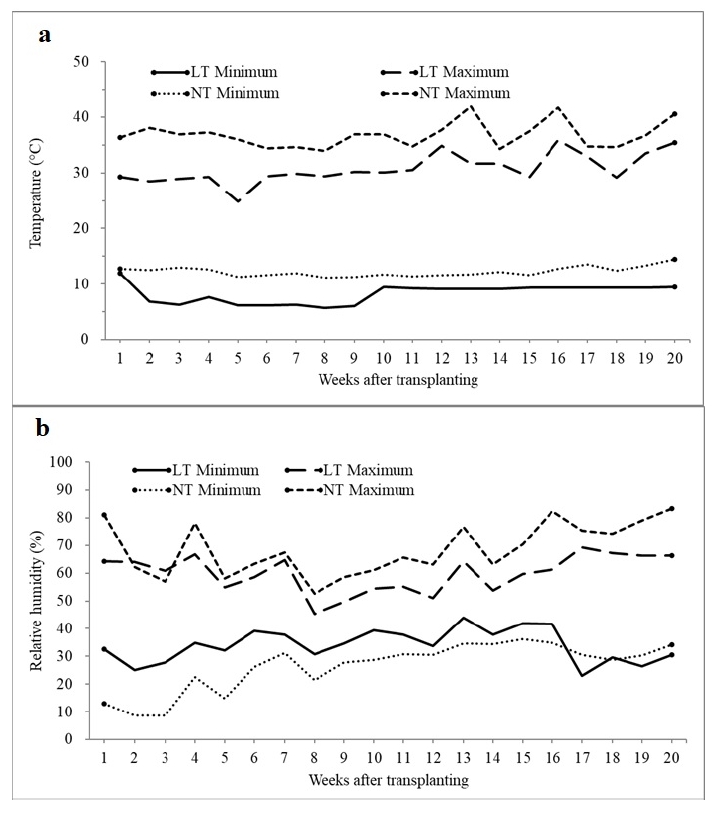
Temperature and relative humidity (RH) during entire growing period were recorded using data logger (WatchDog 1450, Spectrum Technologies Inc., Aurora, USA) in both 10℃ and 15℃ greenhouses (
Fig. 1).
The photosynthetic rate A (µmol CO2 /m2/s), stomatal conductance Cond, gs (mol H2O /m2/s) intercellular CO2 concentration Ci, (µmol CO2 /mol), and transpiration rate Trµmol (µmol H2O /m2/s) measured every 30, 65, 95 and 125 DAT in five randomly selected tomato leaves (T09, T16, T24, T28 and T37). Data were recorded in three plants per accessions in both 10℃ and 15℃ between 11:00 am and 13:00 pm using a portable photosynthesis measurement system (LI-6400, LI-COR Bioscience, Lincoln, NE, USA). Light response curves (PAR) was 500 µmol/m2/s. The leaf chamber temperature was 25℃, and the CO2 concentration was 400 µmol (CO2) /mol. The photosynthetic rate was measured automatically at each irradiation level 3-4 minutes after light exposure.
RESULTS
Growing condition for tomato accessions
Greenhouse condition for both 10℃ and 15℃ was shown in
Fig. 1. Temperature set-point at night in the first week was maintained at 15℃ in both greenhouses for adapting seedlings to the new environment. Minimum and maximum temperatures in a greenhouse with temperature set-point 10℃ were 7℃ and 33℃, respectively, and those in a greenhouse with temperature set-point 15℃ were 11℃ and 38℃ (
Fig. 1a). The mimimum temperature in the greenhouse with temperature set-point 10℃ was slightly lower in the first half of growing period than the second half, probably due to unstable sensor or heating system. However, the minimum temperature after the 9
th week was fully controlled, maintaining a constant temperature of approximately 10℃ (
Fig. 1a). The average RH ranged from 45 to 50% in both greenhouses (
Fig. 1b). Minimum and maximum RH were generally lower and higher, respectively, in the greenhouse with temperature set-point 15℃ than 10℃.
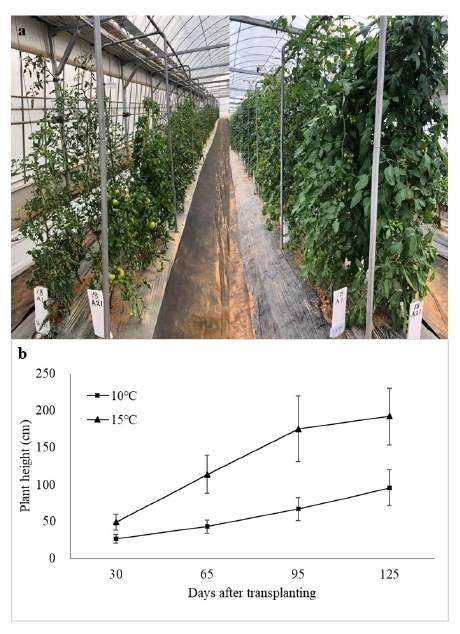
LT significantly affected the plant growth. The plant height in 10℃ was significantly lower than that in 15℃ in all growing period (
Fig. 2a, b). Larger differences in plant height between 10℃ and 15℃, which mean susceptible to LT, were observed in T31, T01, T36 and T05 (35.3, 36.5, 36.9, and 37.3%, respectively) and smaller differences, tolerant to LT, were in T20, T10, T12 and T06 (75.4, 72.9, 69.7, and 66.5%, respectively) (data now shown). Similar to plant height, 40 and 37 accessions have short leaf length and width, respectively, in 10℃ than 15℃. Accessions T33, T10, T20 and T07 showed smallest difference in leaf length such as 89.9, 87.6, 85.9 and 82.5%, respectively (
Table 2). The leaf width of T33 and T10 increased in 10℃ by 19.7 and 8.4%, respectively. However, the other 37 accessions, except for T13, showed decrease in leaf width in 10℃ (
Table 2). The average stem diameter in 40 accessions also tend to be lower in 10℃ than 15℃ but 12 accessions (T10, T33, T07, T37, T27, T17, T14, T26, T21, T40, T04 and T24) in 10℃ had thicker stem diameter than those in 15℃ (
Table 2).
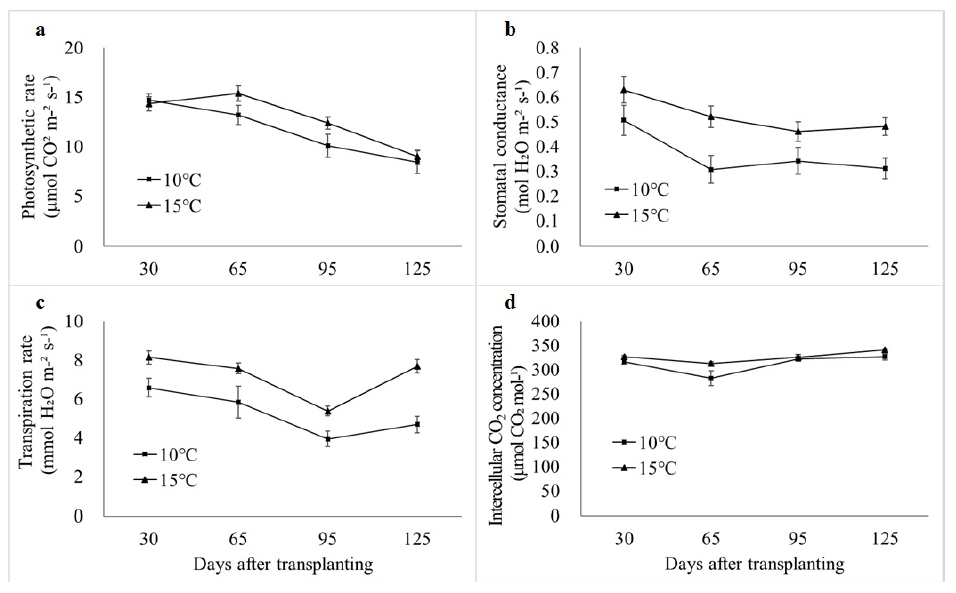
Photosynthesis among 40 accessions significantly reduced in LT (
Fig. 3). In early growing period (until 30 DAT), photosynthetic rate
A was higher in 10℃ than 15℃. However, it was reversed thereafter and the difference was significant between 65 and 95 DAT (
Fig. 3a). Photosynthetic rate
A in both temperature conditions decreased over time during the entire growing period (
Fig. 3a). Stomatal conductance and transpiration rate were also significantly higher in 15℃ than 10℃ in all growing period (
Fig. 3b, c). Intercellular CO
2 concentration was also higher in 15℃ but differences were not significant except for 65 DAT (
Fig. 3d).
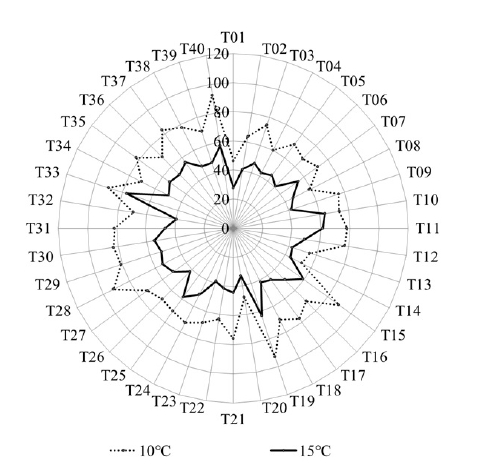
Flowering was significantly delayed in all accessions when grown in 10℃ (
Fig. 4). On average, the flowering initiated 23.9 days later in the accessions grown in 10℃ than those in 15℃. Accessions T01 and T20 flowered earlier than others in both 10℃ (46.3 and 48.0 DAT, respectively) and 15℃ (27.8 and 33.3 DAT, respectively) (
Fig. 4). Difference in days to flowering was greatest in T09 as 82% (76.0 and 41.8 DAT in 10℃ and 15℃, respectively) and smallest in T10 as 15.7% (73.5 and 63.5 DAT in 10℃ and 15℃, respectively).
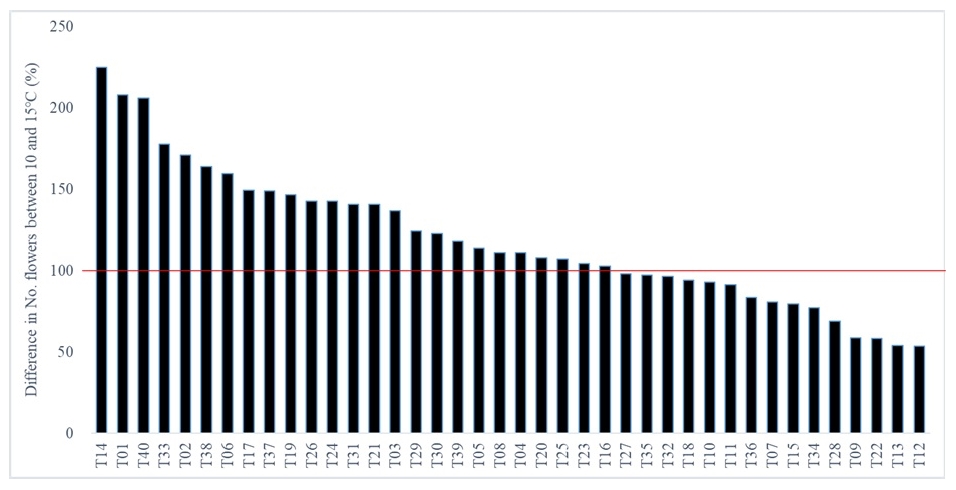
The LT condition significantly influenced the number of flowers among the accessions (
Fig. 5). Among 40, 25 accessions (62.5%) showed the increase in the number of flowers in 10℃ but it decreased in 15 accessions (37.5%). This could be due to the genotype specific interaction with LT. The difference in the number of flowers between 10℃ and 15℃ was largest in T14 (224.7%) and smallest in T12 (53.7%) (
Fig. 4).
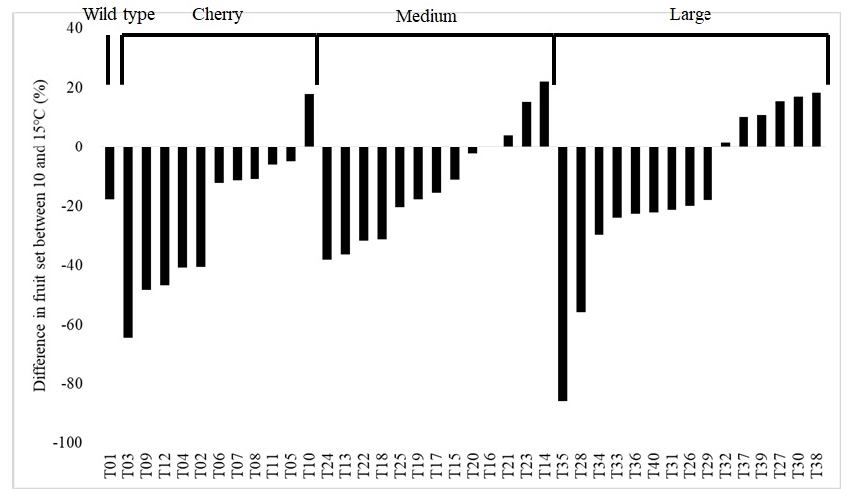
Fruit set tend to decrease in LT condition among the 40 tomato accessions. For instance, the fruit set decreased in 29 accessions (72.5%) whereas increased in 10 accessions (25%) (
Fig. 6) and no difference was observed in a T16 (2.5%). The decrease in fruit set was genotype specific; 79.2% of accessions (19 out of 24) with cherry and medium sized fruits but 40% of accessions (6 out of 15) with large fruits had decreased fruit set ratio in 10℃ (
Fig. 6). The accessions such as T03, T15, T17, T19, T25, T35 and T4 did not bear fruits on both the second and third trusses at 10℃, showing high susceptibility to LT (data not shown). Accessions T10, T14, T30 and T38 displayed highest difference in fruit set but T03, T09, T28 and T35 the smallest (
Fig. 6).
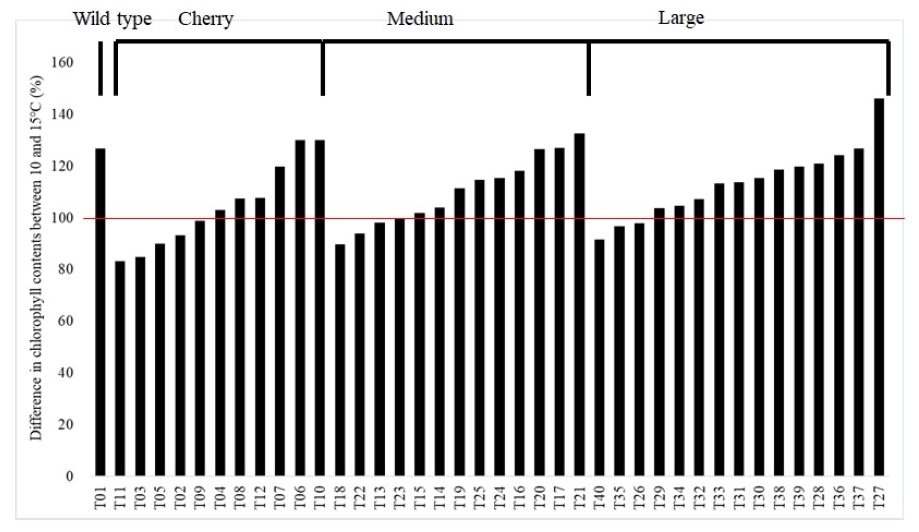
The chlorophyll content in all tomato accessions steadily increased during the entire growing period regardless of temperature conditions in greenhouses. Tomato accessions grown in 10℃ tend to have higher chlorophyll contents than in 15℃ but 10 accessions showed lower chlorophyll content in 10℃ than 15℃ (
Fig. 7). The maximum and minimum chlorophyll contents in 15℃ at 125 DAT were 60.3 (T22) and 32.3 mg/cm
2 (T01), respectively, those in LT were 6.9 (T06) and 37.5 mg/cm
2 (T15), respectively. Differences in chlorophyll contents between 10℃ and 15℃ were highest in T27 (146.2%) and lowest in T11 (83.4%) (
Fig. 7).
To select accessions with LT tolerance, seven traits were chosen such as plant height, leaf length, stem diameter, chlorophyll contents, days to flowering, the number of flowers and fruit set (
Table 3), but leaf width was excluded due to high correlation with leaf length (data not shown). After selecting top four accessions (10th percentile) in each trait, an accession T10, ‘VI037163’, which has cherry type fruits (about 20 g) with brownish skin color and high sugar content, was selected for LT tolerance, based on good performance in six among a total of seven traits (
Table 3).
DISCUSSION
LT has a large effect on the vegetative development in tomato. In our study, the plant growth was significantly delayed in LT in all the 40 tomato accessions (
Fig. 2). Vegetative growth such as plant height, leaf length and width, and stem diameter were significantly reduced in 10℃ compared to 15℃ (
Fig. 2,
Table 2) although some accessions have increased stem diameter in 10℃ (
Table 2). Our results are consistent with previous results that reported reduced growth rate among tomato plants grown in LTN (
Smeets and Garretsen 1986;
Franco 1990;
Nieuwhof et al. 1991; Der Van Ploeg and Heuvelink 2005).
The difference in the growth rate between 10℃ and 15℃ might be the result of the reduced photosynthesis of tomato plants in LT because all photosynthetic parameters were higher in 15℃ than 10℃ (
Fig. 3). The decrease in photosynthetic rate was also observed in wild
Lycopersicon species when day temperature was reduced (
Venema et al. 1999) and
Fang et al. (2018) also reported reduced photosynthetic rate and stomatal conductance in tomato plants under LT stress condition. It is interesting to observe the increased chlorophyll contents among plants grown LT (
Fig. 7) even though photosynthesis was significantly reduced in LT (
Fig. 3). Further study necessary to clarify the relationship between chlorophyll content and photosynthesis in LT condition.
LT also affect the reproductive growth of tomato accessions. Days to flower was significantly delayed in all accessions grown in 10℃ compared to 15℃ (
Fig. 4). However, the delayed flowering may be not the direct cause of LT but delayed growth of tomato plants (
Fig. 2). A previous research supports our assumption, indicating that the delay in the time to flowering of tomato plants in LT is due to the reduced rate of leaf appearance (
Hurd and Graves 1985).
Previously, the only increase in the number of flowers was reported in decreased day (
Rylski 1979), night (
Calvert 1957;
Wittwer and Tuebner 1957) and root temperatures (
Phatak et al. 1966). In our study, however, both the increase (25 accessions) and decrease (15 accessions) in the number of flowers were observed in 10℃ compared to 15℃ (
Fig. 5). Our result suggests that the effect of LT on the number of flowers is genotype specific, showing interaction between genotypes and LT, which is still unknown (
Fig. 5).
Interestingly, 72.0% of accessions (18 out of 25) with small fruits (wild, cherry and medium sized fruits) but only 40% of accessions (6 out of 15) with large fruits had decreased fruit set in 10℃ compared to 15℃ (
Fig. 6), indicating a possible relationship between fruit set and fruit size. The reduced fruit set might be due to poorer quality pollen in LT (reviewed by
Picken 1984) and not effects on stigma level or ovule viability (Fernandez-Munoz and Cuartero 1991).
Seven traits such as plant height, leaf length, stem diameter, chlorophyll contents, days to flowering, the number of flowers and fruit set were used in our study for the selection of tomato accessions with LT tolerance (
Table 3). Selection criteria was based on the difference in those traits between 10℃ and 15℃. The largest percentage in all parameters, except for days to flower and the number of flowers, was assumed to be tolerant to LT (
Table 3). For the days to flowering, accessions that have the smallest percentages were selected. For the number of flowers, accessions that were not affected by LT were selected because both the increase and decrease may be not good for the fruit yield; the increase can make plants use more energy to produce flowers and the decrease can affect the number of fruits produced. The traits chosen may be useful for tomato breeding program because they are easy to investigate in field and the data can be collected in the relatively early growing period.
ACKNOWLEDGEMENTS
This study was supported by a grant (Project No: PJ012662 “Breeding and selection of tomato lines with tolerance to abnormal temperatures stress”) from National Institute of Horticultural and Herbal Science, Rural Development Administration.
Fig. 1Air temperature (a) and relative humidity (b) measured during the entire growth period in greenhouses whose night temperature set-points were 10 and 15℃. LT and NT: greenhouses with night temperature set-points of 10 and 15℃, respectively.
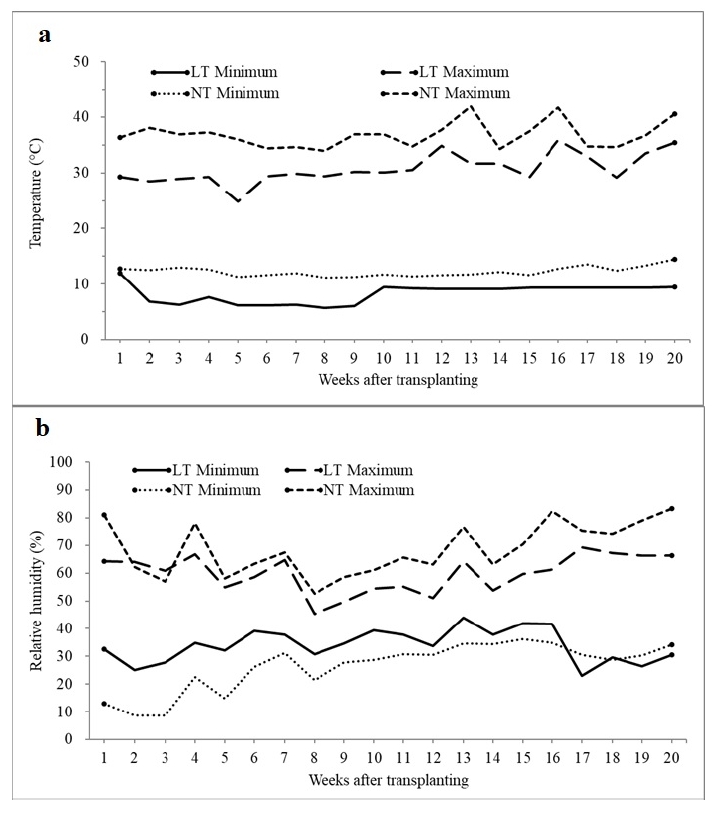
Fig. 2The growth of tomato accessions in 10℃ and 15℃. (a) The tomato plants grown in greenhouses with night temperature set-points of 10℃ (left) and 15℃ (right). Photos were taken 125 days after transplanting. (b) Changes in average plant height of 40 accessions during entire growing period.
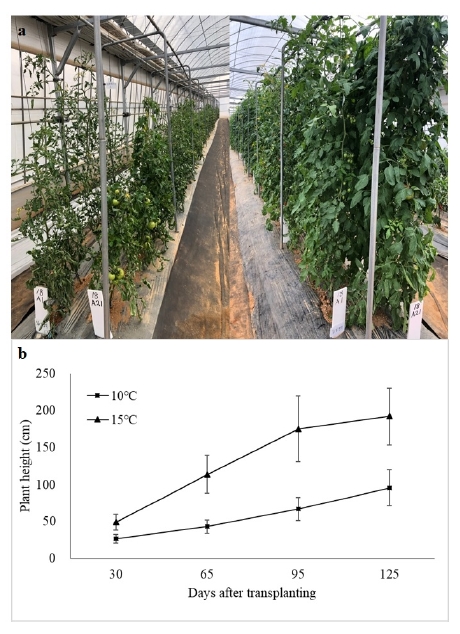
Fig. 3Photosynthetic rate A (a), stomatal conductance (b), transpiration rate (c) and intercellular CO2 concentration of five tomato accessions (T09, T16, T24, T28 and T37), measured 30, 65, 95 and 125 days after transplanting in greenhouses with night temperature set-point of 10℃ and 15℃.
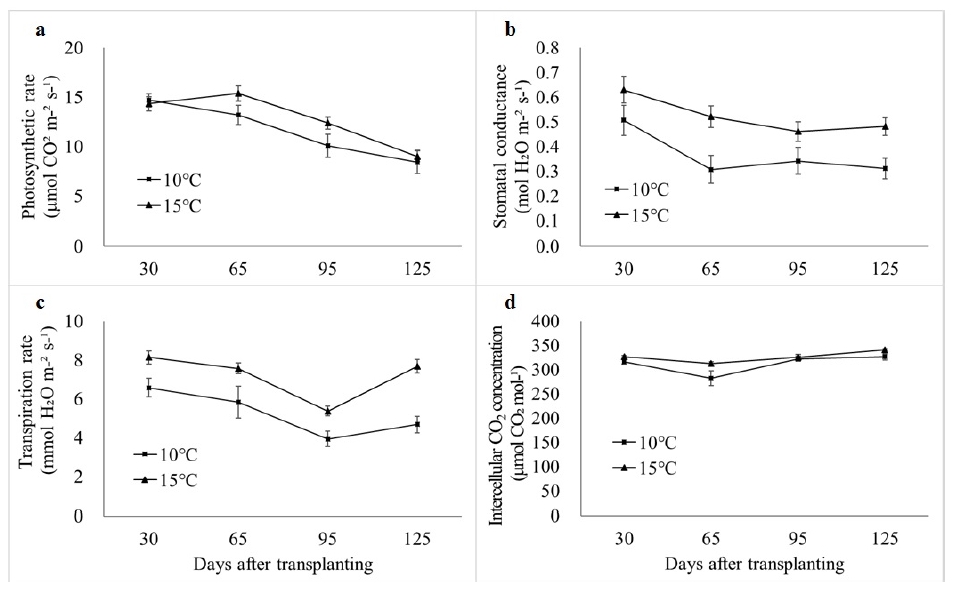
Fig. 4Days to flowering among 40 tomato accessions grown in greenhouses whose night temperature set-point were 10 and 15℃.
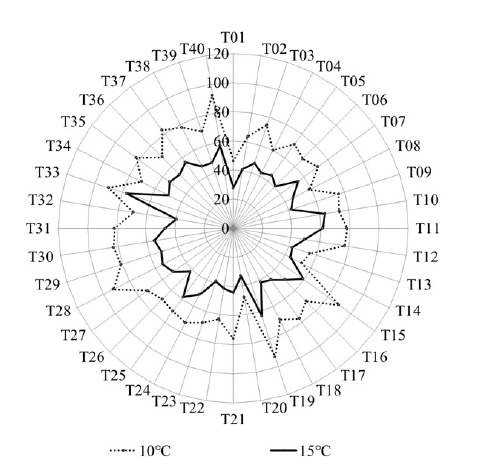
Fig. 5Difference in the number of flowers between 10 and 15℃ among 40 tomato accessions. Lower and higher than 100% means the number of flowers in 10℃ was lower or higher than those in 15℃, respectively. A transverse red line indicates the 100%, no difference in the number of flowers between 10 and 15℃.
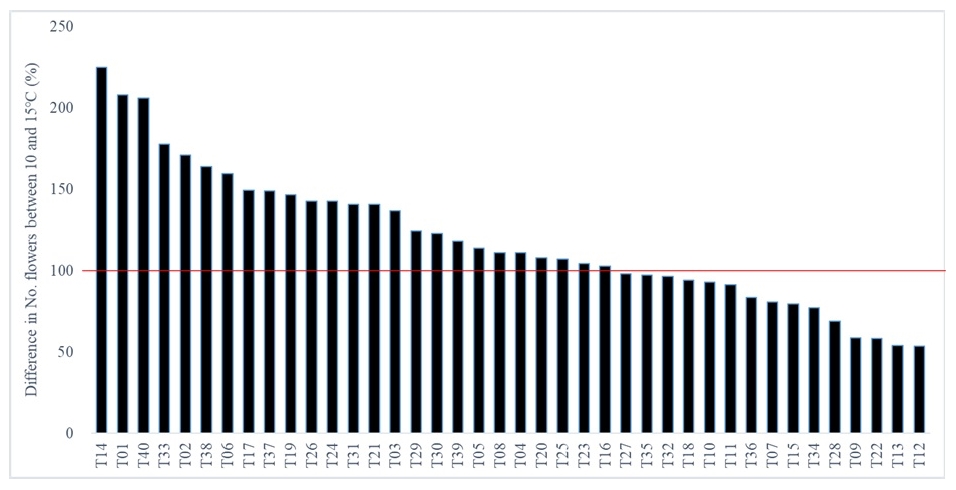
Fig. 6Difference in fruit set between 10 and 15℃ among 40 tomato accessions. Negative percentage means decrease in fruit set of plants grown in 10℃ greenhouse compared to 15℃.
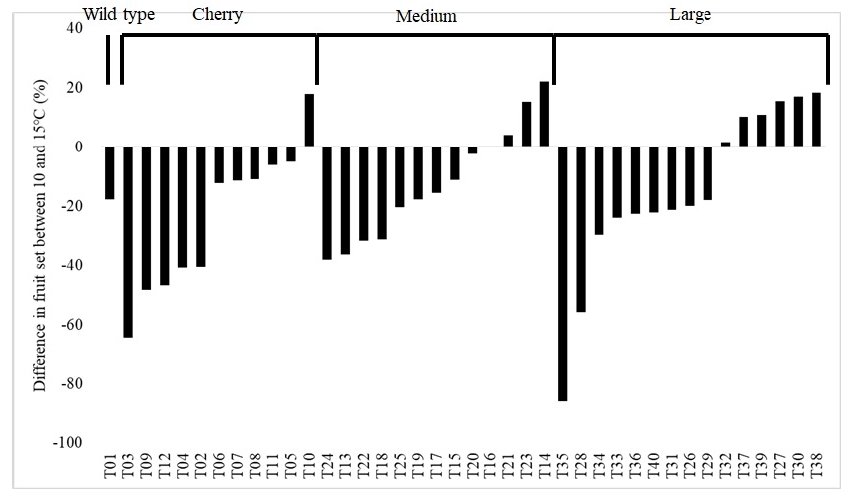
Fig. 7Difference in chlorophyll contents between 10 and 15℃ among 40 tomato accessions. Lower and higher than 100% means the chlorophyll contents of plants grown in 10℃ was lower or higher than those in 15℃, respectively. A transverse red line indicates the 100%, no difference in chlorophyll content between 10 and 15℃.
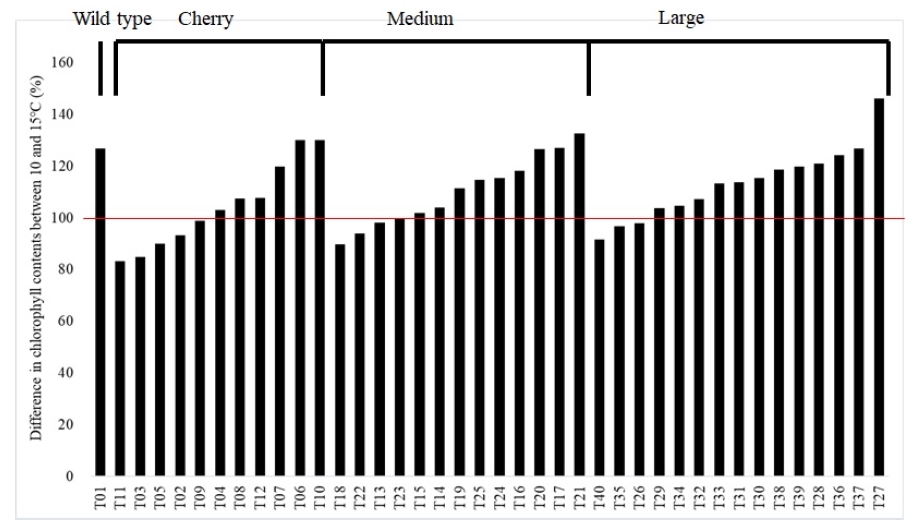
Table 1Accessions used in this study and their fruit types.
Table 1
|
Accessions |
Original name |
Fruit Type |
|
T01 |
L3708 |
Wild |
|
T02 |
Wonhong 1 |
Cherry |
|
T03 |
Wonung 1 |
Cherry |
|
T04 |
Tabtimdaeng T2021 |
Cherry |
|
T05 |
B-Blocking |
Cherry |
|
T06 |
Minichal |
Cherry |
|
T07 |
AVTO1020 |
Cherry |
|
T08 |
Sugar Yellow × 11AVT-2_4, F8 |
Cherry |
|
T09 |
Wonye 9014 |
Cherry |
|
T10 |
VI037163 |
Cherry |
|
T11 |
Wonye 9015 |
Cherry |
|
T12 |
Sincheonggang |
Cherry |
|
T13 |
Tom Little |
Medium |
|
T14 |
Power Guard |
Medium |
|
T15 |
Sugar Yellow × CLN3125-A, F8 |
Medium |
|
T16 |
L05945 |
Medium |
|
T17 |
VI030494 |
Medium |
|
T18 |
TY Strong |
Medium |
|
T19 |
Sugar Yellow × CLN3125-E, F8 |
Medium |
|
T20 |
B-Strong |
Medium |
|
T21 |
12AVT-22 × Bacchus, F2 |
Medium |
|
T22 |
Tom Heart |
Medium |
|
T23 |
IT 032935 (CL80-0-2-0) |
Medium |
|
T24 |
12AVT-14 × Dafnis, F4 |
Medium |
|
T25 |
Tomate RASTEIRO RIO GRANDE |
Medium |
|
T26 |
IT 032964 (CL185-0-1-0) |
Large |
|
T27 |
AVTO1314 |
Large |
|
T28 |
Sinheuksu |
Large |
|
T29 |
Tamseure |
Large |
|
T30 |
AVTO0101 |
Large |
|
T31 |
Pink Top F3 |
Large |
|
T32 |
Avemaria |
Large |
|
T33 |
Tamnara |
Large |
|
T34 |
K151771 |
Large |
|
T35 |
Dafnis |
Large |
|
T36 |
T1-Mobir (Uzbekistan) |
Large |
|
T37 |
Tomate SUPER MARMANDE |
Large |
|
T38 |
SV0244TG |
Large |
|
T39 |
- |
Large |
|
T40 |
IT 247072 |
Large |
Table 2Effects of low temperature on leaf length and width, and stem diameter among 40 tomato accessions.
Table 2
|
Accessions |
Leaf length (cm) |
Difference (%) |
Leaf width (cm) |
Difference (%) |
Stem diameter (mm) |
Difference (%) |
|
|
|
|
10℃ |
15℃ |
10℃ |
15℃ |
10℃ |
15℃ |
|
T01 |
19.0 ± 1.6 |
33.3 ± 1.5 |
57.1 |
19.5 ± 1.6 |
29.8 ± 1.0 |
65.4 |
11.6 ± 0.3 |
12.7 ± 1.1 |
91.3 |
|
T02 |
18.3 ± 1.1 |
45.8 ± 3.4 |
40.0 |
17.6 ± 1.1 |
48.5 ± 3.9 |
35.7 |
15.2 ± 0.5 |
15.8 ± 0.3 |
96.2 |
|
T03 |
23.5 ± 4.5 |
44.7 ± 1.7 |
52.6 |
20.3 ± 3.3 |
43.7 ± 1.5 |
46.5 |
12.0 ± 0.8 |
16.2 ± 0.9 |
74.1 |
|
T04 |
30.0 ± 2.4 |
43.3 ± 2.1 |
69.3 |
31.0 ± 4.0 |
40.0 ± 3.5 |
77.5 |
13.2 ± 0.1 |
12.9 ± 0.1 |
102.3 |
|
T05 |
23.0 ± 1.1 |
36.5 ± 1.8 |
63.0 |
25.6 ± 1.4 |
42.8 ± 2.9 |
59.8 |
17.7 ± 0.3 |
20.2 ± 0.9 |
87.6 |
|
T06 |
31.5 ± 1.1 |
49.5 ± 2.5 |
63.6 |
33.6 ± 1.7 |
43.3 ± 2.5 |
77.6 |
15.4 ± 1.4 |
16.0 ± 0.5 |
96.3 |
|
T07 |
28.3 ± 2.2 |
34.3 ± 3.4 |
82.5 |
27.8 ± 2.5 |
34.8 ± 8.4 |
79.9 |
16.0 ± 0.9 |
14.5 ± 1.7 |
110.3 |
|
T08 |
30.0 ± 0.7 |
45.5 ± 4.6 |
65.9 |
28.6 ± 0.5 |
40.0 ± 3.2 |
71.5 |
13.7 ± 0.4 |
13.7 ± 0.5 |
100 |
|
T09 |
21.3 ± 2.2 |
40.5 ± 2.1 |
52.6 |
25.8 ± 2.9 |
39.0 ± 2.0 |
66.2 |
14.0 ± 0.7 |
16.0 ± 0.9 |
87.5 |
|
T10 |
29.6 ± 1.7 |
33.8 ± 1.6 |
87.6 |
29.8 ± 1.8 |
27.5 ± 2.9 |
108.4 |
15.9 ± 0.8 |
12.5 ± 1.1 |
127.2 |
|
T11 |
23.2 ± 3.0 |
40.0 ± 2.9 |
58.0 |
24.7 ± 4.6 |
39.0 ± 2.4 |
63.3 |
14.9 ± 0.7 |
18.5 ± 1.8 |
80.5 |
|
T12 |
24.5 ± 0.7 |
40.0 ± 3.0 |
61.3 |
27.4 ± 1.3 |
50.5 ± 4.3 |
54.3 |
17.5 ± 0.9 |
18.8 ± 0.9 |
93.1 |
|
T13 |
23.9 ± 0.6 |
29.5 ± 1.9 |
81.0 |
31.0 ± 1.0 |
31.0 ± 1.6 |
100.0 |
13.7 ± 0.3 |
15.2 ± 1.3 |
90.1 |
|
T14 |
29.1 ± 1.2 |
43.3 ± 1.1 |
67.2 |
29.5 ± 1.9 |
42.0 ± 3.0 |
70.2 |
15.0 ± 0.6 |
14.2 ± 0.0 |
105.6 |
|
T15 |
28.6 ± 2.2 |
38.8 ± 2.8 |
73.7 |
28.5 ± 1.4 |
42.5 ± 3.6 |
67.1 |
13.5 ± 0.4 |
16.4 ± 2.0 |
82.3 |
|
T16 |
29.3 ± 2.6 |
40.8 ± 1.9 |
71.8 |
27.5 ± 1.9 |
35.3 ± 1.9 |
77.9 |
15.3 ± 0.8 |
16.3 ± 1.0 |
93.9 |
|
T17 |
18.5 ± 2.9 |
36.8 ± 2.6 |
50.3 |
16.3 ± 2.0 |
34.3 ± 1.5 |
47.5 |
14.1 ± 0.1 |
13.1 ± 0.9 |
107.6 |
|
T18 |
23.6 ± 1.5 |
47.5 ± 4.3 |
49.7 |
27.1 ± 3.2 |
49.3 ± 2.2 |
55.0 |
17.4 ± 0.1 |
18.7 ± 1.0 |
93 |
|
T19 |
29.0 ± 1.5 |
37.3 ± 2.9 |
77.7 |
38.0 ± 2.1 |
46.0 ± 3.1 |
82.6 |
14.8 ± 1.6 |
16.8 ± 0.8 |
88.1 |
|
T20 |
26.9 ± 1.5 |
31.3 ± 1.1 |
85.9 |
29.5 ± 1.7 |
33.0 ± 2.3 |
89.4 |
17.6 ± 0.7 |
18.6 ± 1.1 |
94.6 |
|
T21 |
29.8 ± 2.1 |
49.3 ± 2.7 |
60.4 |
35.5 ± 3.5 |
54.8 ± 6.0 |
64.8 |
15.6 ± 0.6 |
14.9 ± 0.4 |
104.7 |
|
T22 |
25.4 ± 1.1 |
40.5 ± 0.6 |
62.7 |
29.8 ± 1.6 |
39.0 ± 1.7 |
76.4 |
14.0 ± 0.7 |
15.7 ± 0.6 |
89.2 |
|
T23 |
28.4 ± 1.7 |
34.5 ± 1.5 |
82.3 |
30.9 ± 2.2 |
37.5 ± 5.5 |
82.4 |
16.3 ± 0.4 |
16.6 ± 0.9 |
98.2 |
|
T24 |
39.0 ± 0.6 |
60.0 ± 2.0 |
65.0 |
42.0 ± 3.1 |
50.0 ± 0.8 |
84.0 |
15.0 ± 0.4 |
14.8 ± 0.4 |
101.4 |
|
T25 |
30.7 ± 2.4 |
39.5 ± 2.1 |
77.7 |
35.7 ± 5.4 |
40.8 ± 0.5 |
87.5 |
17.8 ± 0.8 |
19.6 ± 1.1 |
90.8 |
|
T26 |
29.6 ± 1.5 |
49.5 ± 4.7 |
59.8 |
30.8 ± 1.2 |
51.0 ± 4.8 |
60.4 |
19.1 ± 0.2 |
18.1 ± 1.3 |
105.5 |
|
T27 |
34.6 ± 0.4 |
44.5 ± 3.2 |
77.8 |
38.8 ± 3.5 |
47.5 ± 4.4 |
79.6 |
19.5 ± 0.3 |
17.9 ± 0.6 |
108.9 |
|
T28 |
28.8 ± 1.2 |
41.0 ± 1.4 |
70.2 |
33.0 ± 1.0 |
46.5 ± 2.1 |
71.0 |
15.4 ± 0.9 |
16.4 ± 1.8 |
93.9 |
|
T29 |
34.5 ± 1.6 |
45.0 ± 1.8 |
76.7 |
39.5 ± 1.2 |
43.5 ± 1.3 |
90.8 |
17.7 ± 0.2 |
18.8 ± 0.8 |
94.1 |
|
T30 |
26.5 ± 1.4 |
42.0 ± 1.8 |
63.1 |
28.0 ± 1.8 |
35.5 ± 2.3 |
78.9 |
17.3 ± 1.1 |
17.5 ± 0.7 |
98.9 |
|
T31 |
30.3 ± 0.7 |
44.0 ± 2.1 |
68.9 |
36.5 ± 2.3 |
47.3 ± 2.6 |
77.2 |
17.2 ± 1.3 |
19.2 ± 1.4 |
89.6 |
|
T32 |
26.3 ± 3.1 |
46.3 ± 1.7 |
56.8 |
32.5 ± 3.3 |
49.8 ± 2.3 |
65.3 |
14.1 ± 0.9 |
16.2 ± 0.2 |
87 |
|
T33 |
32.1 ± 1.7 |
35.7 ± 0.9 |
89.9 |
39.5 ± 2.7 |
33.0 ± 1.0 |
119.7 |
16.4 ± 0.6 |
13.8 ± 1.6 |
118.8 |
|
T34 |
29.7 ± 2.4 |
43.3 ± 3.4 |
68.6 |
32.0 ± 3.5 |
41.5 ± 3.0 |
77.1 |
17.3 ± 1.5 |
18.3 ± 1.6 |
94.5 |
|
T35 |
24.1 ± 1.6 |
51.8 ± 2.3 |
46.5 |
31.0 ± 1.4 |
60.0 ± 1.5 |
51.7 |
17.7 ± 0.4 |
20.3 ± 0.5 |
87.2 |
|
T36 |
20.8 ± 1.3 |
41.8 ± 2.3 |
49.8 |
27.9 ± 2.1 |
47.0 ± 2.6 |
59.4 |
17.3 ± 0.3 |
18.8 ± 0.7 |
92 |
|
T37 |
28.5 ± 1.8 |
39.3 ± 3.1 |
72.5 |
29.8 ± 2.6 |
37.3 ± 2.6 |
79.9 |
18.9 ± 0.4 |
17.2 ± 1.4 |
109.9 |
|
T38 |
29.8 ± 3.0 |
54.0 ± 0.6 |
55.2 |
34.3 ± 2.1 |
53.8 ± 5.4 |
63.8 |
14.8 ± 0.8 |
21.5 ± 1.5 |
68.8 |
|
T39 |
33.5 ± 0.7 |
54.5 ± 2.1 |
61.5 |
37.5 ± 2.9 |
52.8 ± 2.0 |
71 |
18.0 ± 1.2 |
21.0 ± 1.0 |
85.7 |
|
T40 |
29.3 ± 2.4 |
42.8 ± 2.5 |
68.5 |
30.0 ± 1.5 |
46.5 ± 3.3 |
64.5 |
17.1 ± 0.6 |
16.6 ± 0.4 |
103 |
Table 3Selection criteria for accessions with low temperature tolerance and selected accessions for 7 traits.
Table 3
|
Traits |
Selection criteria (difference between 10 and 15℃ ) |
Selected accessions (10th percentile) |
|
Plant height |
The largest percentage |
T20, T10, T12, T06 |
|
Leaf length |
The largest percentage |
T33, T10, T20, T07 |
|
Stem diameter |
The largest percentage |
T10, T33, T07, T37 |
|
Chlorophyll contents |
The largest percentage |
T27, T21, T10, T06 |
|
Days to flowering |
The smallest percentage |
T10, T33, T14, T25 |
|
The number of flowers |
No difference |
T16, T27, T35, T32 |
|
Fruit set ratio |
The largest percentage |
T14, T38, T10, T30 |
References
Reference
- Adams S, Cockshull K, Cave C. 2001. Effect of temperature on the growth and development of tomato fruits. Ann. Bot.. 88: 869-877.
- Calvert A. 1957. Effect of the early environment on development of flowering in the tomato. I. Temperature. J. Hortic. Sci.. 32: 9-17.
- De Koning. 1988. The effect of different day/night temperature regimes on growth, development and yield of glasshouse tomatoes. J. Hortic. Sci.. 63: 465-471.
- De Koning. 2000. The effect of temperature, fruit load and salinity on development rate of tomato fruit. Acta Hortic.. 519: 85-93.
- Der Van Ploeg A, Heuvelink E. 2005. Influence of sub-optimal temperature on tomato growth and yield: a review. J. Hortic. Sci. Biotechnol. 80: 6652-6659.
- Elings A, Kempkes FLK, Kaarsemaker RC, Ruijs MNA, Van De, Dueck TA. 2005. The energy balance and energy-saving measures in greenhouse tomato cultivation. Acta Hortic.. 691: 67-74.
- Fang X, Zaiqiang Y, Liyun Z. 2018. Low temperature and weak light affect greenhouse tomato growth and fruit quality. J. Plant Sci.. 6: 16-24.
- Fernandez-Munoz R, Cuartero J. 1991. Effects of temperature and irradiance on stigma exertion, ovule viability and embryo development in tomato. J. Hortic. Sci.. 66: 395-401.
- Franco T. 1990. Effects of stressful and unstressful low temperature on vegetable crops: morphological and physiological aspects. Acta Hortic.. 287: 67-76.
- Hoek HIS, Hanisch Ten, Keijzer CJ, Schel JH, Dons HJM. 1993. Development of the fifth leaf is indicative for whole plant performance at low temperature in tomato. Ann. Bot.. 72: 367-374.
- Hurd RG, Graves CJ. 1985. Some effects of air and root temperatures on the yield and quality of glasshouse tomatoes. J. Hortic. Sci.. 60: 359-371.
- Khayat E, Ravad D, Zieslin N. 1985. The effect of various night-temperature regimes on the vegetative growth and fruit production of tomato plants. Sci. Hortic.. 27: 9-13.
- Nieuwhof M, Garretsen F, Van Oeveren. 1991. Growth analyses of tomato genotypes grown under low energy conditions. Neth. J. Agr. Sci.. 39: 191-196.
- Nieuwhof M, Keizer LCP, Van Oeveren. 1997. Effects of temperature on growth and development of adult plants of genotypes of tomato (Lycopersicon esculentum Mill.). J. Genet. Breed.. 50: 185-193.
- Paul EMM. 1984. Genotypic variation in the response to suboptimal temperatures of growth in tomato (Lycopersicon esculentum Mill.). New Phytol.. 98: 221-230.
- Phatak SC, Wittwer SH, Teubner FG. 1966. Top and root temperature effects on tomato flowering. J. Am. Soc. Hortic. Sci.. 88: 527-531.
- Picken AJF. 1984. A review of pollination and fruit set in the tomato (Lycopersicon esculentum Mill.). J. Hortic. Sci.. 59: 1-13.
- Rylski I. 1979. Fruit set and development of seeded and seedless tomato fruits under diverse regimes of temperature and pollination. J. Am. Soc. Hortic. Sci.. 104: 835-838.
- Smeets L, Garretsen F. 1986. Growth analyses of tomato genotypes grown under low night temperatures and low light intensity. Euphytica. 35: 701-715.
- Venema JH, Posthumus F, Vanhasselt PR. 1999. Impact of suboptimal temperature on growth, photosynthesis, leaf pigments and carbohydrates of domestic and high-altitude wild Lycopersicon species
. J. Plant Physiol.. 155: 711-718.
- Wittwer SH, Tuebner FG. 1957. Cold exposure of tomato seedlings and flower formation. J. Am. Soc. Hortic. Sci.. 67: 369-376.