Abstract
Traditional rice landraces of coastal areas in Bangladesh are distinct in respect to their phenotypes, responses to salt stress and yield attributes. In characterization of coastal rice landraces, 46 rice genotypes were tested for salt tolerance at their seedling and reproductive growth stages. Through the cluster analysis following standard evaluation score (SES), genotypes were divided into five categories (highly susceptible, susceptible, moderately tolerant, tolerant and highly tolerant) at their seedling stage. Three coastal genotypes, viz. chorbindi, joyni and kedgrangal mota, were grouped as highly saline tolerant at their seedling stage. In studying reproductive stage salinity screening, based on mean score values obtained from different morphological and yield parameters (plant height, number of effective tillers/hill, number of filled grains, panicle length, 100-grain weight, and straw dry weight), kutiakon and kajolshail were identified as the salinity tolerant landraces while tulsimala, dudhkalom, birindi, bushihara, lalmota, chorboleshor, lalchikon and bashful chikon were found as moderately tolerant to salinity. Finally, the molecular characterization using two simple sequence repeat (SSR) markers, viz., RM493 and RM3412 revealed holde mota, dingamoni, kedgrangal mota, sada mota, laxmima, dishari, lal chikon, kalijira, kalo khaya, khaioz, chorbindi, kajolshail, kutiakon, bamonkhir, dudhmora, sakhorkhora, mota dhan, and sorbimaloti as tolerant landraces. These identified salt-tolerant landraces can be used as promising germplasm resources for breeding salt-tolerant high-yielding rice varieties in the future.
-
Key words: SSR markers, Coastal rice, Salinity, Seedling and reproductive stage
INTRODUCTION
Bangladesh is a country of over 147,570 km
2 having 20% land under the coastal area. Out of 2.85 million hec-tares of the coastal and offshore areas, about 0.83 million hectares are cultivable lands, which cover over 30% of the total cultivable lands of Bangladesh, and are affected by varying degrees of salinity. Like other parts of the world, salinity is also becoming a predominant factor for poor crop productivity. Land use in these areas is very poor, which is roughly 50% of the country’s average (
Petersen and Shireen 2001). Soil salinity is believed to be mainly responsible for low land use as well as cropping intensity in the area (
Rahman and Ahsan 2001). Rice,
Oryza sativa (2n = 24), is the staple food of nearly 50% of the world’s population (
Singh and Mishra 1997). Rice is the main food grain of Bangladeshi people and Bangladesh is the 4
th largest rice producing country in the world. The people of the south and south-west region of Bangladesh largely depend on agriculture, especially, on rice cultivation. The currently cultivated rice varieties in this region are not fit for survival with the increased soil salinity. Salinity in-trusion adversely affects coastal agriculture of Bangladesh through limiting fresh water availability (
Singh et al. 1994). The cropping pattern of this region is mainly transplanting
aman rice dominated where the farmers still depend on the local rice landraces as they withstand high water levels and varying degrees of salinity. However, the main concern is the average rice yield of these regions which is much lower than that of the national (2.94 t/ha) (BBS 2012). There are many good landraces of rice which have good potentials to be developed as high yielding variety (HYV) with greater stress/salinity tolerance. The adverse environmental con-dition and low narrow-sense heritability of salt tolerance made conventional plant selection method much harder. Use of DNA marker seems to be the suitable option for ef-ficient assessment and selection of plant materials (
Gregorio 1997). SSR (Simple Sequence Repeat) microsatellite markers were proved to be ideal for marker assisted selection, making genetic maps and studying genetic diversity in different germplasms (
Islam 2004). SSR markers are an important tool to identify gene for salt tolerance that might be helpful for plant breeders to develop new cultivars.
The present study was designed to assess 46 coastal rice landraces for salinity tolerance at their seedling and repro-ductive stages at the Field Agronomy Laboratory and Genetic Engineering and Molecular Breeding Laboratory of Patuakhali Science and Technology University. The main objective of this study was to uncover prospective rice landraces tolerant to salinity that could be an important baseline data to undertake recombination breeding pro-gram with desired high yielding genotypes to develop salt tolerant rice varieties.
MATERIALS AND METHODS
The experiment was done at the Stress Agronomy Lab., net house of Agronomy Field Laboratory and Genetic Engineering and Molecular Breeding Laboratory in Patuakhali Science and Technology University. The phenotypic study was done through hydroponic culture in the net house of Agronomy field laboratory maintaining semi-controlled environment by wrapping transparent polyvinyl sheet on the roof. Molecular screening of the collected germplasm was done in Genetic Engineering and Mole-cular Breeding Lab. using simple sequence repeat (SSR) markers to find out salinity tolerant landraces. The experi-ment was carried out during the period from January 2018 to December 2018.
Collection and screening of germplasms for salinity tolerance
The 46 rice germplasms were collected from different sources, mainly from the coastal area of Patuakhali region, Bangladesh Rice Research Institution (BRRI) and Bangladesh Institutions of Nuclear Agriculture (BINA), Mymensingh. Phenotypic study was done primarily during the seedling stage at a salinity level of 10 dS/m using SES (Standard Evaluation Score;
Gregoria et al. 1997) under a hydroponic system according to Yosida
et al. (1976). Twenty-eight (28) germplasms showing better performance at seedling stages were used for phenotypic study against salinity (EC 10 ds/m) at reproductive stages based on plant height (cm), number of effective tillers/hill, number of filled grain/hill, panicle length (cm), 100 grain weight (g) and straw dry weight/ hill (g) following the IRRI standard protocol (
Gregoria et al. 1997). To categorize the salinity tolerance at the repro-ductive stage, a mean score (ms) was calculated for each genotype using the score derived from morphological growth parameters (
Tahjib-Ul-Arif et al. 2018). Finally, 38 genotypes were selected for molecular study using micro-satellite (SSR) markers.
Genomic DNA isolation
Genomic DNA was extracted from 25-day-old rice seedlings using Promega A1120 DNA purification kit (Promega, USA) following the manufacturer’s instruc-tions. The amount of extracted DNA was estimated by Nano-100 micro spectrophotometer (BOYN, China). A desirable concentration, 25 ng/µL, was obtained through dilution.
Amplification of SSR markers and evaluation of genotypes
Primarily, a set of 10 SSR primer pairs were selected for the analysis of variability to salinity tolerance among the collected landraces (
Table 1). Based on intensity of bands, consistency within individuals, presence of smearing and potential for population discrimination, two polymorphic SSR markers RM3412 and RM493 linked to
saltol QTL (quantitative trait loci) of chromosome 1 (
Ganie et al. 2014) were selected to evaluate rice germplasms for salt tolerance.
A 15 µL of polymerase chain reaction (PCR) cocktail was prepared by mixing 1 µL forward primer, 1 µL reverse primer, 1.0 µL of extracted rice DNA (25 ng/µL), 7.5 µL Go Taq® Green Master Mix (Promega, USA) and 4.5 µL of nuclease free water. PCR (Applied Biosystems, USA) was performed following the conditions as follows: initiali-zation at 94℃ for 5 minutes and 35 cycles of amplification at 94℃ for 45 seconds, 60℃ for 45 seconds, 72℃ for 1 minute and final extension at 72℃ for 5 minutes. After PCR, electrophoresis was done in 2% agarose gel added with ethidium bromide to detect band. Banding patterns were visualized with ultraviolet transilluminator gel docu-mentation system. The banding patterns of 38 rice ger-mplasms were scored comparing with tolerant and sus-ceptible control variety. The genotypes having similar banding pattern to the improved salt tolerant variety were considered as tolerant and similar to improved salt sus-ceptible variety were considered as salt susceptible.
Statistical analyses
Data recorded on crop characteristics were subjected to statistical analysis through computer based statistical pro-gram JMP 8 (JMP®8, SAS Institute Inc., Cary, NC, 1989-2019) following the basic principles. To characterize the level of salinity tolerance of the 46 aman rice genotypes, the least significant mean values of genotypes for SES were used in multivariate cluster analysis (by UPGMA; un-weighted pair group method with arithmetic mean) using JMP8 software. To test the treatment effects on six yield contributing characters (plant height, number of effective tillers, panicle length, and number of filled grain, 100-grain weight, and straw dry weight reduction), data were sub-jected to analysis of variance (ANOVA) and means were compared using Tukey’s honestly significant difference (HSD) post hoc test.
RESULTS
Morphological screening against Salinity at seedling stage
In our study, the modified SES scoring system was employed to characterize salt-tolerant rice genotypes dur-ing the seedling stage under saline conditions (NaCl at 10 dS/m). Growing of rice genotypes in saline condition caused some visual foliar symptoms. The foliar symptoms include partial or total browning and necrosis of leaf blades. Initially the intensity of the necrosis was minor. As the duration of salt treatment increased, injury symptoms were intensified in all rice genotypes. Growth of the plants stopped by the 2
nd or 3
rd day, followed by chlorosis and leaf bleaching from the tip of the leaf blade to the leaf base on the 4
th day or 5
th day. By the 7
th to 9
th days of post sal-inization, susceptible seedlings were dead. Tolerant var-ieties also showed the same early response to salinity stress, but on the 4
th or 5
th day, they showed some signs of re-covery, such as leaf greening and growing of the youngest leaf (
Fig. 1).
The degree of tolerance pattern of genotypes falling into five clusters is given in
Fig. 2. The distribution pattern indicated that cluster I, the largest cluster, comprised of 19 genotypes followed by cluster III (14), cluster IV (7), cluster II (4), and cluster V (2). The clustering pattern re-vealed that based on SES means, cluster I was assigned as tolerant (T), with the average group mean of 3.4. The genotypes such as
holde mota, dudhkalom, kutiakon, dudhmora, chaulmagi, laxmima, tulsimala, kajolshail, kachamota, bouhari, sorbimaloti, BRRIdhan 41,
birindi, biyonti monkhusi, chorboleshor, kamola mota, bushihara, bashful chikon and
bamonkhir were grouped together in cluster I. Cluster II had a group SES mean of 1.2 and was considered as a highly tolerant (HT) group including
kedgrangal mota,
joyni,
Chorbindi, and check tolerant variety BINAdhan 10. Cluster III had a group SES mean of 5.3 and was classified as medium tolerant (MT). Genotypes
motha mota, sakhorkhora, bashful, lal chikon, calendar mota, dingamoni, sada mota, dishari, khaioz, mothadhan, and
monteshor were grouped in this cluster. Cluster IV was classified as sensitive (S) with a group SES mean of 7.4 where sensitive check BRRIdhan 49 and other popular coastal genotypes such as
kalijira, barimota, choulamuthahari, dholkochua, moulata were found. The highest group SES mean of 9.2 was observed for cluster V and hence classified as highly sensitive (HS). It included the sensitive coastal genotypes
chinigura and
nakuchimota.
Based on seedling stage salinity screening results (
Fig. 2), we selected 28 genotypes where two tolerant and one susceptible check were also included. Six yield contri-buting characters
viz. plant height, number of effective tillers/hill, panicle length, number of filled grain, 100-grain weight, and straw dry weight were investigated main-taining a saline (10 dS/m) and non-saline (at base value) condition to study salinity tolerance of 28 genotypes at the reproductive stages. Data on the set parameters of both conditions (saline and non-saline) were compared assess-ing the reduction percentages which are represented in
Fig. 3,
Table 2.
Plant height (cm): We found variation in plant height among the genotypes. Plant height was inhibited in all genotypes, percent plant height reduction was highly sig-nificant (
P < 0.01) among genotypes against salinity treat-ment (
Fig. 3,
Table 2). Among the check genotypes, BRRIdhan 49 showed highest shoot length reduction (28%) followed by BRRIdhan 41 (10%) while BINAdhan 10 was reduced by 7%. Almost all coastal genotypes displayed highest shoot growth reduction, indicating the sensitivity of coastal genotypes to salt stress. Among the coastal genotypes,
laxima showed highest shoot length reduction (29%) followed by
kedrengal mota (28%),
motha mota (26%), and the lowest shoot length reduction was found in
khaioz, bamonkhir and
kalo khaya at the rate of 12.5, 12.6, 13%, respectively.
Effective tillers/hill: Plants’ survivability is very dif-ficult in saline conditions as their normal physical growth is severely affected. In our experiment, the reduction per-centage of effective tillers ranged between 0% to 95.12%, where the genotype
kutiakon recorded the lowest reduction of effective tillers at 0%, while the highest reduction of effective tillers (95.12%) was recorded in the variety
shorbimaloti. On the other hand, tolerant check varieties BRRIdhan 41 and BINAdhan 10 respectively showed 15.15% and 33.33% effective tiller reductions under saline conditions (
P < 0.01;
Table 2).
Panicle length (cm): Panicle length is one of the im-portant reproductive characters and is sensitive against salinity. Due to salinity panicle length was reduced sig-nificantly (
P < 0.01) than that under non-saline conditions (
Fig. 3,
Table 2). Panicle length reduction ranged from 0.59% to 77.43%, where local
aman genotype
kutiakon showed the lowest and
kamola mota showed the highest value (
Table 2). Tolerant check BRRIdhan 41 and BINAdhan 10 showed 3.59% and 20.57% panicle length reduction, respectively. Besides,
lal chikon (5%),
birindi (5%),
kajolshail (11%),
dudhkalom (13%),
holde mota (17%),
tulsimala (22%),
lal mota (29%),
bushihara (18%),
bashful chikon (20%) and
joyni (18%) showed relatively lower reduction percentages.
Filled grains/hill: Salinity affects the plants nutrient uptake from their available sources. Highly significant reduction percentage in case of filled grain/hill was found in our study (
P < 0.01;
Table 2). Filled grain reduction percentage was found lowest in
tulsimal (0.32%) and highest (100%) in
sorbimaloti. Surprisingly, both the tolerant check varieties BRRIdhan 41 and BINAdhan 10 showed 83.4% and 77.53% filled grains reduction, respect-ively.
Kajolshail (1%),
dudhkalom (42%),
kutiakon (62%) had relatively lower reduction percentages in case of filled grain.
Hundred (100) grain weight (gm): Grain weight loss under salinity is also an important characteristic effect. Due to lack of proper nutrient and loss of physical strength, rice genotypes are not able to produce healthy seeds. Thus, grain weight loss was another factor to consider in this study. By measuring 100-grain weight, significant reduc-tion in weight loss was evident (
P < 0.01;
Table 2). Highest amount of reduction was found in
sorbimaloti (100%) and lowest in
dudhkalom (8.31%) which was less than the tolerant check BINAdhan 10 (14.18%).
Tulsimala (17%),
chorbindi (13%),
kutiakon (12%) also showed lower grain weight loss.
Straw dry weight (gm): Dry matter is an important trait to assess the performance of rice genotypes for salt tolerance. Due to improper nutrition and loss of various ions (exo-osmosis) from the plant body, it reduces the strength of plant and final loss of their weight. In our study, straw dry weight reduced remarkably in rice under salinity (
Fig. 3,
Table 2). There was 1.53% to 84.46% reduction of weight loss.
Kutiakon (2%) had the lowest dry weight loss while
joyni (85%) showed the highest straw weight loss in re-lation to check BRRIdhan 41 (5.77%).
Kajolshail (4%),
chorbindi (5%),
khaioz (35%),
calendar mota (39%) and
chorboleshor (20%) showed lower straw weight reduction (
Table 2).
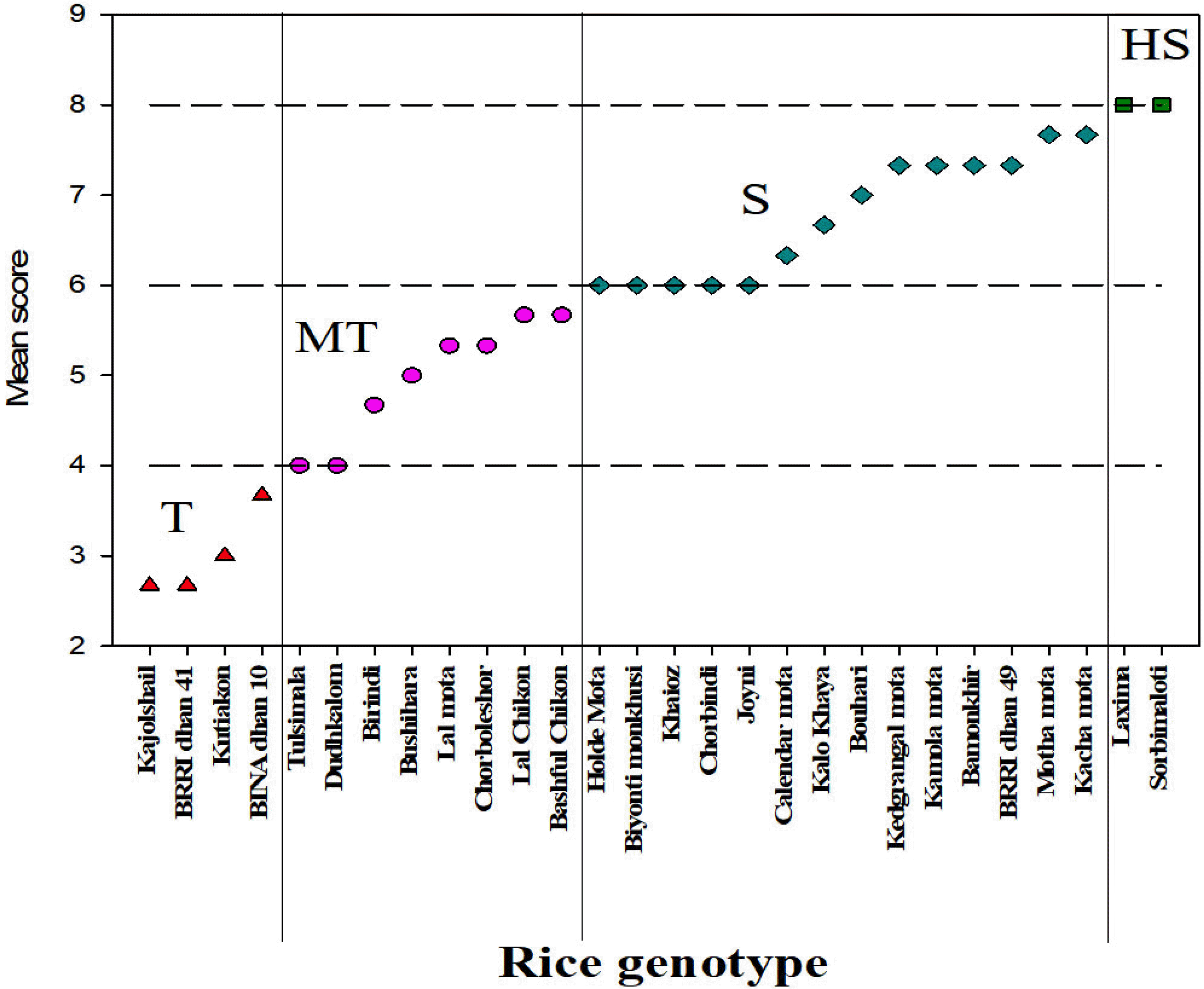
To sum up the above results, according to Tahjib-Ul-Arif
et al. (2018), based on the reduction percentage (%R) for growth parameters each genotype has been given a score and finally their mean score (ms) for each genotype was calculated to categorize the salinity tolerance at the reproductive stage (
Fig. 4). Here we found four (04) cate-gories of salinity tolerance levels
viz., tolerant (T), medium tolerant (MT), susceptible (S) and highly susceptible (HS). We found
kajoshail,
kutiakon and two tolerant check BRRIdhan 41 and BINAdhan 10 to be categorized under the tolerant group (1 < mean score ≥ 3).
Tulshimala,
dudhkalom, birindi, bushihara, lalmota, chorboleshor, lalchikon and
bashful chikon were grouped into moderately tolerant category (3 < mean score ≥ 5). In the susceptible category (5 < mean score ≥ 7), we found
holde mota,
biyonti monkhushi, khaioz, chorbindi, joyni, calendar mota, kalo khaya, bouhari, kedgrangl mota, kamola mota, bamonkhir, motha mota, kach mota, and sensitive check variety BRRIdhan 49. Lastly,
laxima and
sorbimaloti were grouped into the highly susceptible group because of their highest mean score values (mean score < 7). We could not find any rice genotypes in the highly tolerant group that we reported in their seedling stage screening (
Fig. 2). Further-more, we noticed that members of previously categorized seedling stage screening showed inconsistent tolerance patterns at their reproductive stage screening (
Figs. 2,
4). Specifically,
chorbindi, joyni and
kedgrangl mota were found to be highly tolerant against salinity at the seedling stage, and subsequently at their reproductive stage screen-ing they were grouped into susceptible group with high mean score values (
Fig. 4). This indicates that stress re-sponsiveness of plants may vary with their growth and developmental stages.
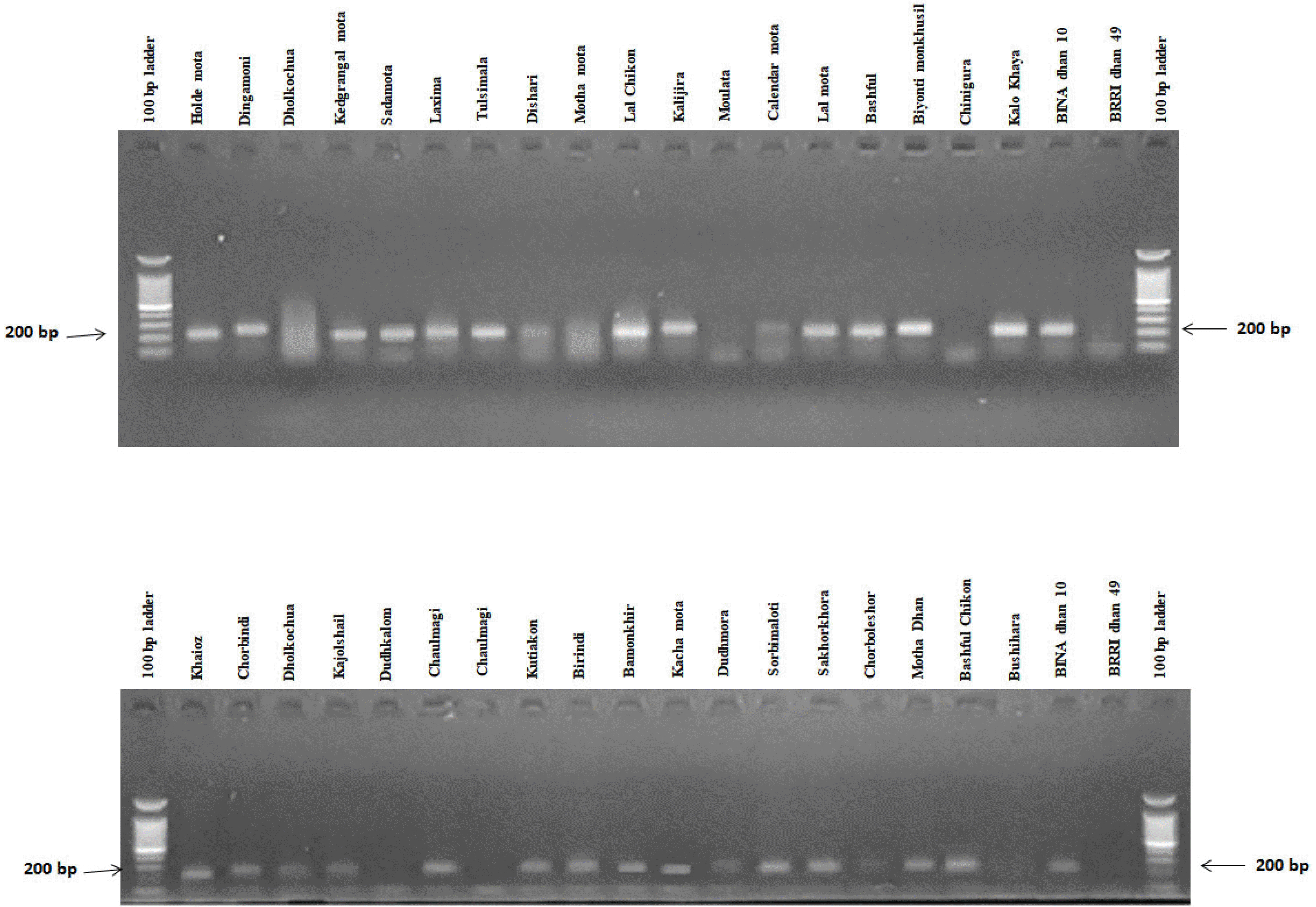
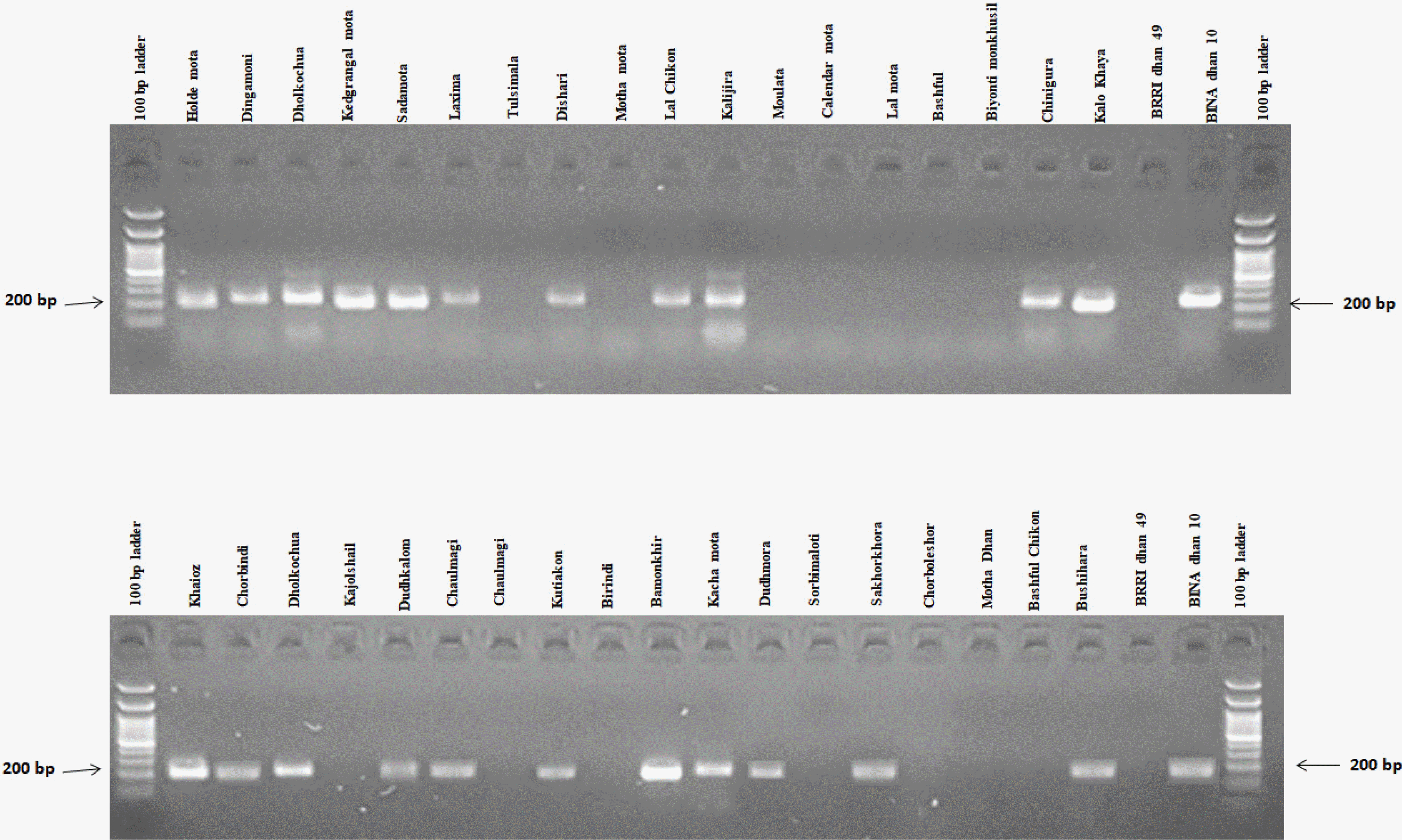
In this experiment, RM1032, RM11133, RM11669, RM315, RM12213, RM8094, RM493, RM3412, RM8095 and RM7075 primers were used for polymorphism survey with selected rice germplasms (
Table 1). Among all pri-mers, two SSR markers viz., RM3412 and RM493 showed high polymorphism, and afterward they were selected to screen rice germplasms for salt tolerance. According to the phenotypic performance, BINAdhan 10 was considered as tolerant and BRRI dhan 49 was considered as susceptible. The lines having similar banding pattern to BINAdhan 10 were considered as tolerant and those similar to BRRIdhan 49 were considered as salt susceptible (
Table 3,
Figs. 5,
6). In respect to RM493 primers, 28 genotypes were found to show banding patterns similar to tolerant genotype BINAdhan 10. In the same reaction 9 genotypes were found sus-ceptible compared to the susceptible variety BRRIdhan 49 (
Fig. 5). Twenty-two (22) genotypes were detected as tolerant and 16 susceptible genotypes were found by comparing to the tolerant variety BINAdhan 10 and sus-ceptible variety BRRIdhan 49 in case of reaction with RM3412 (
Fig. 6). From the summarized
Table 3, it is shown that 19 genotypes
viz. holde mota, dingamoni, kedgrangal mota, sada mota, laxmima, dishari, lal chikon, kalijira, kalo khaya, khaioz, chorbindi, kajolshail, kutiakon, bamonkhir, dudhmora, sakhorkhora, mota dhan, sorbimaloti and BINAdhan 10 were found as tolerant and 5 genotypes
viz. BRRIdhan 49,
bashful chikon, birindi, moulata and
motha mota were found susceptible against the tested markers. Moreover, 10 genotypes
viz.
tulsimala, calendar mota, lal mota, biyonti monkhusi, bashful, dudhkalom, kachamota, chorboleshor, bushihara and
kamola mota were found tolerant only with RM493, and 3 genotypes
joyni,
chaulmagi and
chinigura were found tolerant with RM3412. From the phenotypic data (SES and %R of dif-ferent growth characters),
kajolshail, BRRIdhan 41,
kutiakon and BINAdhan 10 were identified as salt tolerant rice genotypes, whereas BRRIdhan 49,
dholkochua, laxmima, sorbimaloti, motha mota, holde mota, dingamoni and
joyni were identified as salt susceptible. From SSR screening we observed that
kutiakon, kajolshail and
tulsimala showed higher polymorphism.
DISCUSSION
The present study was designed to assess 46 Bangladeshi coastal rice landraces for salinity tolerance at their seedling and reproductive stages to uncover prospective rice landraces tolerant to salinity that could be an important baseline data to undertake recombination breeding pro-gram with desired high yielding genotypes to develop salt tolerant rice varieties. Reports revealed that cluster analysis is a promising tool to screen a large number of germplasms based on their similarity (
Ali et al. 2014;
Chunthaburee et al. 2016). Here we used the simple visual salt injury scoring technique (SES) which is widely used for characterization as it represents overall plant responses to salt stress (
Gregorio 1997). In the seedling stage clustering of the genotypes, we found a wide variation in salinity tolerance and identified 5 clusters of tolerance levels: cluster I to cluster V (
Fig. 2). Specifically, here we identified 37 rice genotypes under varying levels of salinity tolerance (highly tolerant, tol-erant, and medium tolerant) based on SES. A wide vari-ation in phenotypes between tolerant and susceptible lines using SES was also observed in several studies (
Islam 2004;
Rubel et al. 2014). In the subsequent evaluation of the well performed genotypes in the seedling stage, they were employed to uncover their salinity tolerance at the reproductive stage which is considered to be another sens-itive stage to salinity stress (
Table 2,
Fig. 4;
Gregorio 1997;
Tahjib-Ul-Arif et al. 2018). Among the traits evaluated for salt stress responses, genotypes varied significantly for shoot parameters, suggesting that salinity tolerance is more likely controlled in the shoot. Salinity stress might affect cell division or cell enlargement which causes reduced plant height (
Mojakkir et al. 2015). This possibly explained the occurrence of induced DNA methylation in shoots as compared to some rice varieties tested for salinity response (
Karan et al. 2012). In the case of evaluating effective tiller numbers per hill, we found a wide variation where reduc-tion percentage of effective tillers ranged between 0% to 95.12% with genotype
kutiakon recording the lowest re-duction of effective tillers of 0% while the highest reduc-tion of effective tillers (95.12%) was recorded in variety
shorbimaloti (
Table 2).
Rashid et al. (2017) also reported that effective tiller number was drastically reduced under 12 dS/m compare to the control. Other studies also reported that tiller numbers decreased progressively with increasing salinity levels (
Desai et al.1975;
Saxena and Pandey 1981). Panicle length is one of the important reproductive charac-ters sensitive to salinity. In our study panicle length re-duced significantly (
P < 0.01) against salinity compared to non-saline conditions.
Marassi et al. (1989),
and Abdullah et al. (2001) also denoted similar findings where panicle length was significantly decreased due to salinity stress. Salinity affects the plant’s nutrient uptake from available sources. Highly significant %R in case of filled grain/hill was found in our study (
P < 0.01).
Rad et al. (2012) observed that effect of different growth stages and levels of salinity on total number filled grains per panicle was highly significant (
P < 0.01) where number of filled grains re-duced drastically in saline conditions compare to non-saline conditions. Other studies pointed out that salinity affects yield through reducing number of filled grains. This possibly happens from the reduced seed set in the panicle which might be the result of decreased pollen viability or malfunctioning of the stigma surface, or both (
Khatun and Flowers 1995a;
Abdullah et al. 2001). We noticed signifi-cant effects of salinity on grains being observed by many researchers. And thus, salinity causes significant yield loss by affecting the number and weight of grains (
Katerjia et al. 2003). Again, dry matter is an important trait to assess the performance of rice genotypes for salt tolerance. Due to improper nutrition and loss of various ions (exo-osmosis) from the plant body, it reduces the strength of plant and final loss of their weight. In our study, straw dry weight reduced remarkably in rice under salinity. There was 1.53% to 84.46% reduction of weight loss were evident (
Table 2). This result is consistent with the results from other studies where tolerant genotypes also showed lower reduction of dry weight compared to susceptible genotypes (
Islam 2004). Other researchers reported that increase in salinity levels reduced the fresh biomass and dry biomass (
Roy et al. 2002;
Javed et al. 2006). Finally, at the reproductive stage screening we identified four (04) categories of salinity tolerance levels
viz., tolerant (T), medium tolerant (MT), susceptible (S), and highly susceptible (HS) (
Fig. 4). We noticed that members of previously categorized seedling stage screening showed inconsistent tolerance pattern at their reproductive stage salinity screening (
Figs. 2,
4). Specifically,
chorbindi, joyni and
kedgrangl mota were found as highly tolerant against salinity at the seedling stage and subsequently at their reproductive stage screen-ing they were grouped into susceptible group with high mean score value (
Fig. 4). This indicates that stress responsiveness of plants may vary with their growth and develop-mental stages. Our result is also consistent with another study where different growth stages of rice were found to have different sensitivity to salinity (
Rad et al. 2012).
Besides, for determining genetic variations and un-raveling the cultivar’s identity, molecular marker-based genotyping is considered as a very useful technique. SSR markers are widely used due to their robustness to differ-entiate individual germplasm accessions (
Xu et al. 2004;
Jeung et al. 2005) and they have higher coverage for the rice genome (
McCouch et al. 2002).
In our study, for the molecular validation stage we used two polymorphic SSR markers RM3412 and RM493 which are linked to
saltol QTL of chromosome 1 of rice (
Ganie et al. 2014). These two primer sets showed poly-morphisms in the studied rice germplasms as they exhi-bited different banding patterns and discriminate tolerant lines from susceptible with relation to the tolerant variety BINAdhan 10 and susceptible variety BRRIdhan 49 (
Figs. 5,
6). Finally, comparing banding profiles of RM3412 and RM493, nineteen genotypes
viz. holde mota, dingamoni, kedgrangal mota, sada mota, laxmima, dishari, lal chikon, kalijira, kalo khaya, khaioz, chorbindi, kajolshail, kutiakon, bamonkhir, dudhmora, sakhorkhora, mota dhan, sorbimaloti, and BINAdhan 10 were found as tolerant and five geno-types
viz. BRRIdhan 49,
bashful chikon, birindi, moulata and
motha mota were found as susceptible. These SSR markers were also reported to show different combinations of alleles (haplotypes) in 94 rice genotypes (
Krishnamurthy et al. 2016).
Ganie et al. (2014) also reported that of RM3412 and RM493 had higher number of allelic vari-ations in their studied rice genotypes. Likewise,
Thomson et al. (2010),
and Babu et al. (2014) used RM3412 and RM493 for their discriminating abilities in salt tolerance.
Islam et al. (2012) reported 10 alleles using markers RM3412 and RM493 where each of them showed poly-morphism information content (PIC) of 0.81 and were suc-cessfully used to identify haplotype diversity in salt tol-erant genotypes.
To compare the results of our phenotypic and SSR based genotyping of the coastal rice genotypes, it is clearly evident that some genotypes showed tolerant features against salinity in the phenotypic study, but they were not consistent in the DNA profiling using SSR markers (viz dudhkalom, chaulmagi, tulsimala,kacha mota, biyonti monkhusi, kamola mota, bashful chikon, bushihara and joyni). On the contrary, some genotypes showed banding patterns as similar as the tolerant check (BINAdhan 10), but they showed susceptibility to salt stress in both the phenotypic screening (viz, dingamoni, sada mota, dishari, lal chikon, kalijira, kalo khaya, khaioz, sakhorkhora, and mota dhan). This might be the fact that our markers (RM493 and RM3412) may not be linked strongly with QTLs or genes which are responsible for salt tolerance. Further study should be done using a considerable number of markers polymorphic to the salt tolerance.
CONCLUSION
Here we identified some salt tolerant lines both phe-notypically (screening at seedling and reproductive stage) and by using SSR markers RM493 and RM3412 from 46 coastal rice genotypes. These two markers could be ef-ficiently used to identify salt tolerant lines in rice and can also be used in marker-assisted selection (MAS) for breed-ing, QTL mapping, studying genetic diversity in germplasms and gene pyramiding in rice salinity breeding. In a nutshell, considering all the results from different studies, we identi-fied three salt tolerant (kajolshail, tulsimala and kutiakon), eight medium salt tolerant (tulsimala, dudhkalom, birindi, bushihara, lal mota, chorboleshor, lal chikon and bashful chikon), thirteen salt susceptible (holde mota, biyonti monkhusi, khaioz, chorbindi, joyni, calendar mota, kalo khaya, bouhari, kedgrangal mota, kamola mota, bamonkhir, motha mota, and kacha mota), and two (laxmima and sorbimaloti) highly salt susceptible genotypes. Our salt tolerant high yielding coastal genotypes could be utilized for recom-bination breeding program with desired high yielding genotypes to increase grain quality, tolerance and higher yield.
ACKNOWLEDGEMENTS
The research work has been carried under the project fund provided by Ministry of Science and Technology (MoST) of Government of Bangladesh (39.00.0000.09.06.2017/ BS-S-217). Authors are grateful to Professor Dr. Abul Kashem Chowdhury, Department of Plant Breeding and Genetics of Patuakhali Science and Technology (PSTU) for laboratory help.
-
AUTHOR CONTRIBUTIONS
GS and NUA planned this study. FI conducted research work under supervision of GS and NUA and wrote the manuscript. GS critically edited the manuscript. All authors read and approved the final version of the manuscript.
-
CONFLICT OF INTEREST
No potential conflict of interest relevant to this article was reported.
Fig. 1Showing phenotypic differences between the salt tolerant and salt susceptible rice genotypes at the seedling stage.

Fig. 2Clustering of 46 genotypes by UPGMA based on SES responses to salinity stress at seedling stage. T: Tolerant, HT: Highly Tolerant, MT: Moderately Tolerant, S: Susceptible, HS: Highly Susceptible, and ‘I-V’ indicate five (5) clusters.

Fig. 3Showing phenotypic differences between the salt tolerant and salt susceptible rice genotypes at their active tillering [a (i) &b (iv)], ripening [a (ii)&b (v)] and grains in a panicle [a (iii)&b (vi)].

Fig. 4Comparative salinity tolerance ranking of 26 rice genotypes at reproductive stage [T (1 < mean score ≥ 3) = Tolerant, MT (3 < mean score ≥ 5) = Moderately Tolerant, S (5 < mean score ≥ 7) = Susceptible, HS (mean score < 7) = Highly Susceptible].
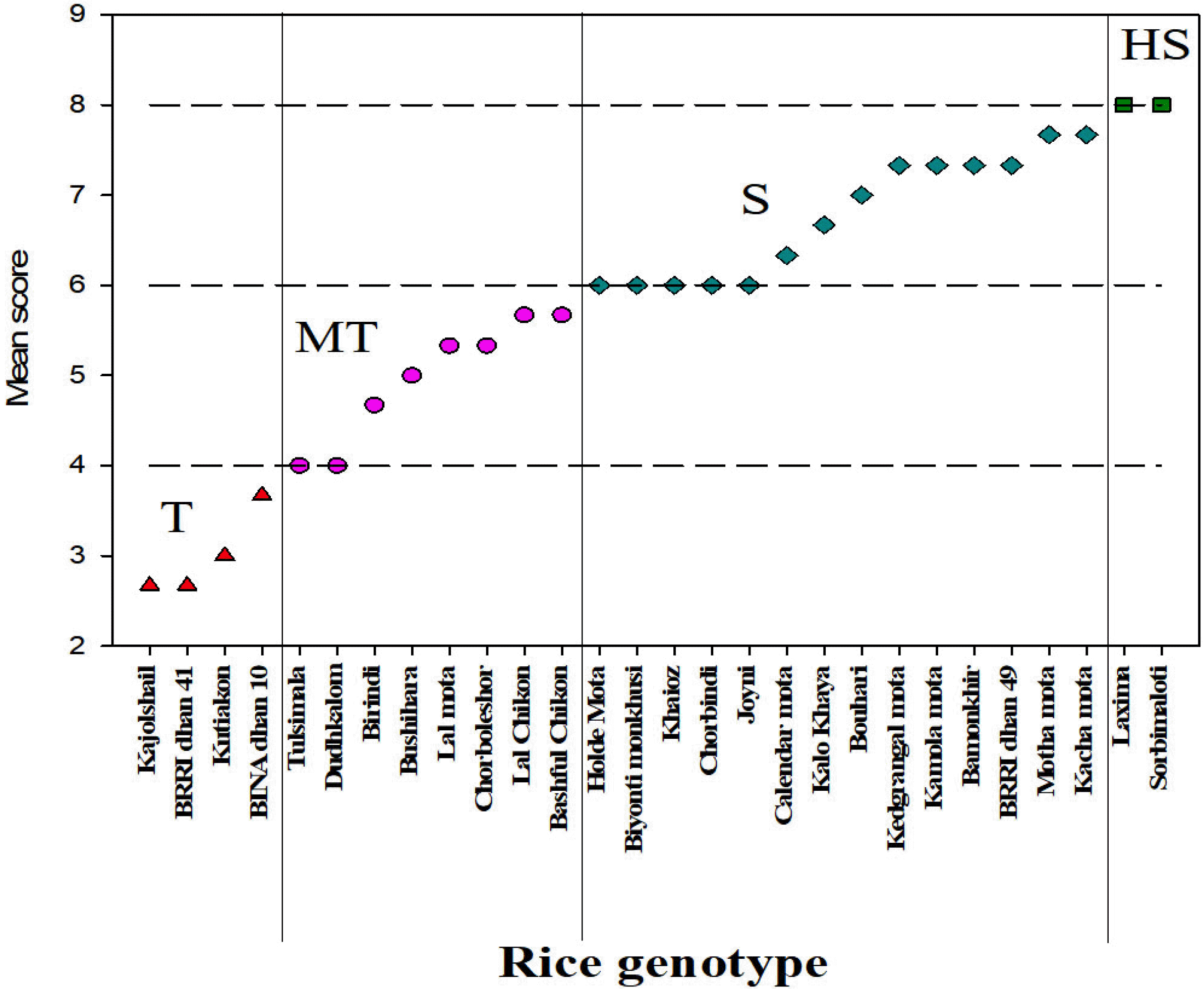
Fig. 5Banding profiles of rice genotypes using primer RM493.
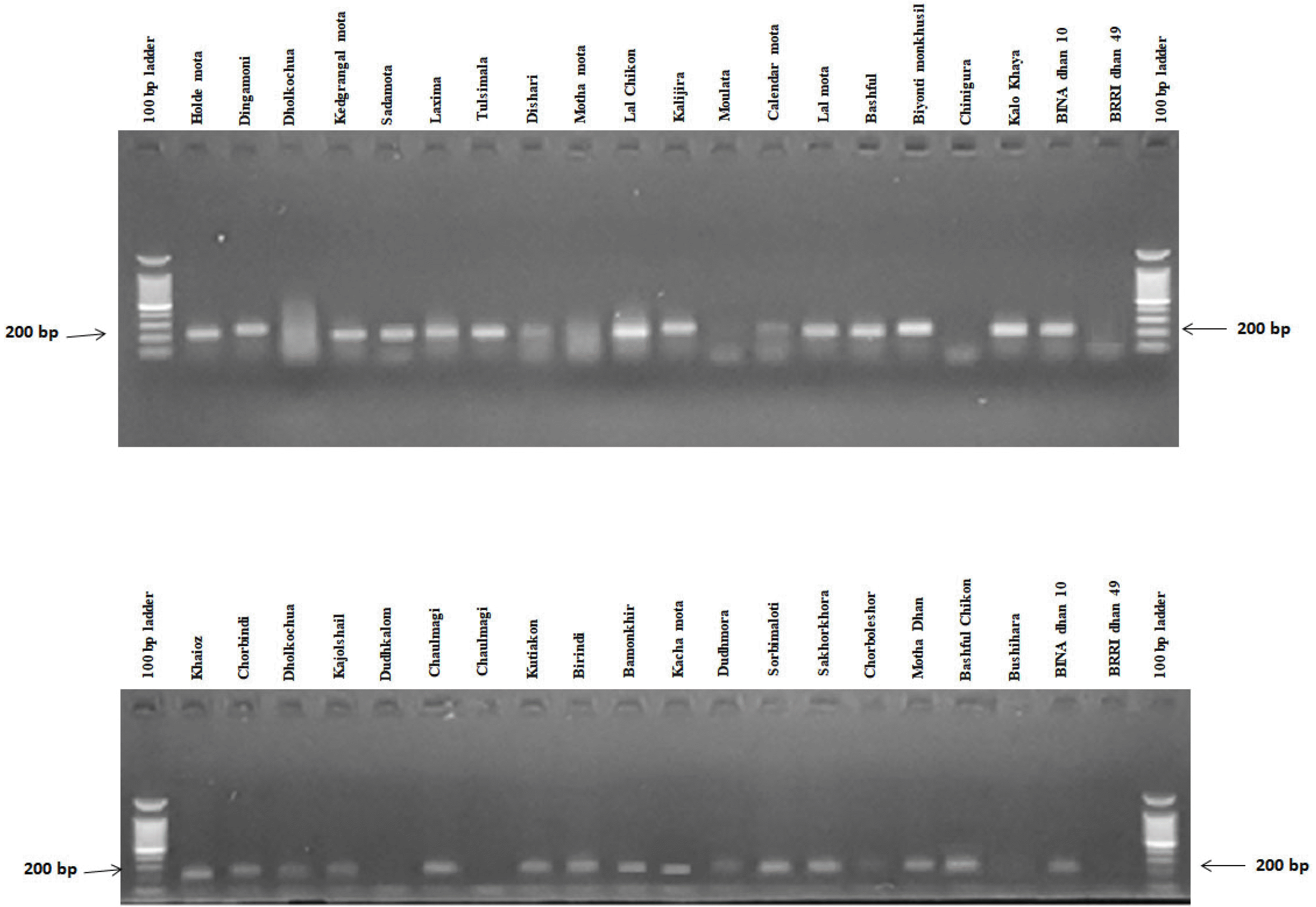
Fig. 6Banding profiles of rice genotypes using primer RM3412.
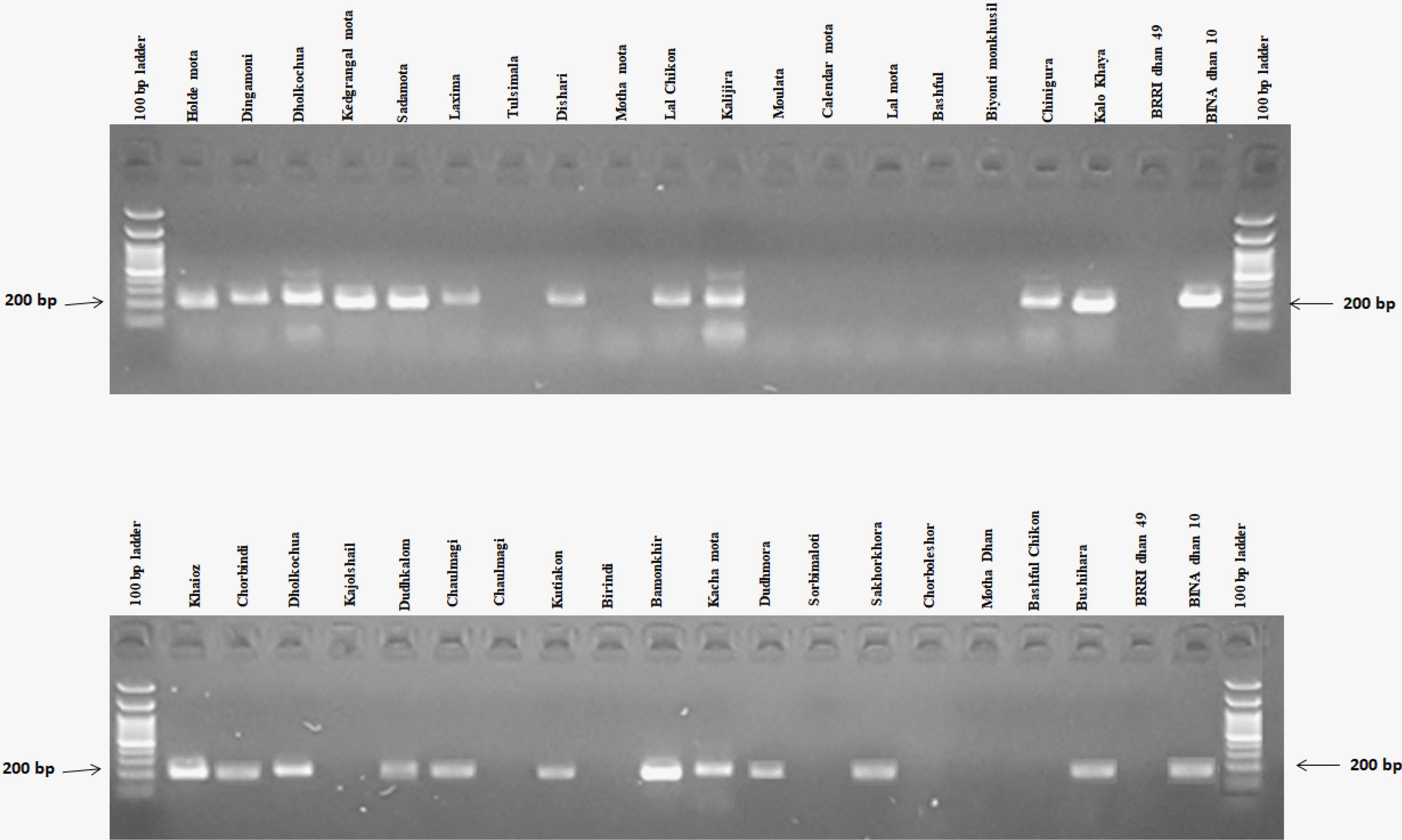
Table 1List of 10 SSR markers used for polymorphism profiling.
Table 1
|
Sl. |
Name |
Forward |
Reverse |
Prod. Size (bp) |
Annealing (℃) |
|
1 |
RM1032 |
TGGCACTTCACGTAGCAAAC |
TGGTTCTGTTCTTGTGGCTG |
149 |
50 |
|
2 |
RM11133 |
GTCCCGGGATTATTTACGTTGG |
TCATTATCAATCCCGTCCACTAGC |
162 |
50 |
|
3 |
RM11669 |
AAACCGTTCCAGGGAGACTGACC |
TCGTCTGATCCATCCATCCATCC |
168 |
50 |
|
4 |
RM315 |
GAGGTACTTCCTCCGTTTCAC |
AGTCAGCTCACTGTGCAGTG |
133 |
55 |
|
5 |
RM12213 |
GGTCCTAGTGCACGGCTTGTCC |
ACCGGTTGTGCACACAAATTGC |
98 |
50 |
|
6 |
RM8094 |
AAGTTTGTACACATCGTATACA |
CGCGACCAGTACTACTACTA |
209 |
55 |
|
7 |
RM493 |
TAGCTCCAACAGGATCGACC |
GTACGTAAACGCGGAAGGTG |
211 |
55 |
|
8 |
RM3412 |
AAAGCAGGTTTTCCTCCTCC |
CCCATGTGCAATGTGTCTTC |
211 |
55 |
|
9 |
RM8095 |
TTCCGTGGACATGATGAATC |
AAGGTTTAGAACATACACACCGTT |
111 |
55 |
|
10 |
RM7075 |
TATGGACTGGAGCAAACCTC |
GGCACAGCACCAATGTCTC |
155 |
50 |
Table 2Different morphological parameters of rice genotypes along with their individual score and mean score values.
Table 2
|
Genotype |
Plant height (cm) |
|
Effective tillers hill‒1
|
|
Panicle length (cm) |
|
Filled grain hill‒1
|
|
100-Grain wt. (g) |
|
Straw dry wt. hill‒1 (g) |
|
ms |
|
|
|
|
|
|
|
|
nsl
|
sl
|
%R
|
S
|
nsl
|
sl
|
%R
|
S
|
nsl
|
sl
|
%R
|
S
|
nsl
|
sl
|
%R
|
S
|
nsl
|
sl
|
%R
|
S
|
nsl
|
sl
|
%R
|
S
|
|
|
Holde Mota
|
124 |
100 |
19fgh
|
5 |
|
12 |
4 |
64j
|
7 |
|
22 |
18 |
17k
|
3 |
|
725 |
77 |
89gh
|
9 |
|
3 |
2 |
30ij
|
3 |
|
103 |
20 |
81a
|
9 |
|
6 |
|
Kedgrangal mota
|
145 |
104 |
28ab
|
9 |
|
14 |
2 |
88cd
|
9 |
|
25 |
12 |
55d
|
7 |
|
669 |
46 |
93def
|
9 |
|
3 |
1 |
51g
|
5 |
|
41 |
22 |
47h
|
5 |
|
7.33 |
|
Laxima
|
136 |
96 |
29a
|
9 |
|
14 |
2 |
88cd
|
9 |
|
26 |
13 |
49e
|
7 |
|
285 |
2 |
99ab
|
9 |
|
2 |
1 |
65e
|
7 |
|
71 |
24 |
67bc
|
7 |
|
8 |
|
Tulsimala
|
141 |
106 |
25bcd
|
7 |
|
11 |
5 |
53k
|
5 |
|
22 |
17 |
22i
|
3 |
|
314 |
313 |
0n
|
1 |
|
1 |
1 |
17l
|
1 |
|
69 |
21 |
70b
|
7 |
|
4 |
|
Motha mota
|
139 |
103 |
26abc
|
9 |
|
12 |
2 |
81f
|
9 |
|
25 |
11 |
56d
|
7 |
|
1133 |
46 |
96bcd
|
9 |
|
3 |
2 |
51g
|
5 |
|
52 |
20 |
61de
|
7 |
|
7.67 |
|
Lal Chikon
|
140 |
106 |
24cd
|
7 |
|
15 |
4 |
75g
|
7 |
|
22 |
21 |
5m
|
1 |
|
726 |
52 |
93def
|
9 |
|
2 |
1 |
33i
|
3 |
|
60 |
20 |
67bc
|
7 |
|
5.67 |
|
Calendar mota
|
121 |
98 |
19fgh
|
5 |
|
14 |
2 |
88cd
|
9 |
|
22 |
11 |
47e
|
7 |
|
162 |
13 |
92efg
|
9 |
|
2 |
1 |
47h
|
5 |
|
41 |
25 |
39j
|
3 |
|
6.33 |
|
Lal mota
|
130 |
104 |
20efg
|
5 |
|
16 |
4 |
74gh
|
7 |
|
25 |
18 |
29h
|
3 |
|
1049 |
38 |
96bcd
|
9 |
|
3 |
2 |
29j
|
3 |
|
59 |
29 |
52g
|
5 |
|
5.33 |
|
Biyonti monkhusi
|
115 |
94 |
19fgh
|
5 |
|
10 |
3 |
70i
|
7 |
|
23 |
12 |
48e
|
7 |
|
200 |
40 |
80jk
|
7 |
|
3 |
1 |
58f
|
5 |
|
32 |
15 |
52g
|
5 |
|
6 |
|
Kalo khaya
|
120 |
105 |
13jk
|
3 |
|
15 |
4 |
76g
|
7 |
|
24 |
13 |
47e
|
7 |
|
288 |
3 |
99ab
|
9 |
|
3 |
1 |
75c
|
7 |
|
62 |
19 |
69b
|
7 |
|
6.67 |
|
Khaioz
|
111 |
97 |
13jk
|
3 |
|
9 |
2 |
82ef
|
9 |
|
21 |
10 |
54d
|
7 |
|
161 |
23 |
86hi
|
9 |
|
3 |
1 |
56f
|
5 |
|
32 |
21 |
35k
|
3 |
|
6 |
|
Chorbindi
|
112 |
98 |
13jk
|
3 |
|
10 |
3 |
66j
|
7 |
|
21 |
6 |
71b
|
9 |
|
190 |
31 |
84i
|
9 |
|
3 |
1 |
77bc
|
7 |
|
20 |
19 |
5mn
|
1 |
|
6 |
|
Kajolshail
|
140 |
109 |
22def
|
7 |
|
11 |
5 |
56k
|
5 |
|
22 |
20 |
11l
|
1 |
|
110 |
109 |
1n
|
1 |
|
3 |
2 |
13mn
|
1 |
|
23 |
22 |
4mn
|
1 |
|
2.67 |
|
Dudhkalom
|
145 |
112 |
22def
|
7 |
|
8 |
6 |
29m
|
3 |
|
24 |
21 |
13l
|
3 |
|
973 |
569 |
42m
|
5 |
|
2 |
2 |
8° |
1 |
|
33 |
18 |
45hi
|
5 |
|
4 |
|
Kutiakon
|
143 |
111 |
22def
|
7 |
|
7 |
7 |
0° |
1 |
|
20 |
20 |
1n
|
1 |
|
1158 |
451 |
61l
|
7 |
|
1 |
1 |
12mn
|
1 |
|
22 |
21 |
2n
|
1 |
|
3 |
|
Kacha mota
|
131 |
104 |
20efg
|
5 |
|
13 |
2 |
85de
|
9 |
|
21 |
7 |
66c
|
9 |
|
1415 |
77 |
95cde
|
9 |
|
2 |
1 |
67de
|
7 |
|
63 |
27 |
57f
|
7 |
|
7.67 |
|
Sorbimaloti
|
140 |
115 |
18gh
|
5 |
|
14 |
1 |
95a
|
9 |
|
22 |
7 |
67c
|
9 |
|
832 |
0 |
100a
|
9 |
|
2 |
0 |
100a
|
9 |
|
70 |
27 |
62de
|
7 |
|
8 |
|
Chorboleshor
|
131 |
108 |
18gh
|
5 |
|
9 |
1 |
85de
|
9 |
|
20 |
12 |
37f
|
5 |
|
929 |
9 |
99ab
|
9 |
|
2 |
1 |
29j
|
3 |
|
27 |
22 |
20l
|
1 |
|
5.33 |
|
Bashful chikon
|
137 |
106 |
23cde
|
7 |
|
13 |
4 |
71hi
|
7 |
|
24 |
19 |
20ijk
|
3 |
|
1330 |
73 |
95cde
|
9 |
|
2 |
1 |
22k
|
3 |
|
57 |
27 |
53g
|
5 |
|
5.67 |
|
Bushihara
|
129 |
98 |
24cd
|
7 |
|
14 |
4 |
71hi
|
7 |
|
20 |
16 |
18jk
|
3 |
|
768 |
82 |
89gh
|
9 |
|
2 |
2 |
10no
|
1 |
|
68 |
41 |
40j
|
3 |
|
5 |
|
Kamola mota
|
132 |
109 |
17ghi
|
5 |
|
9 |
1 |
88cd
|
9 |
|
25 |
6 |
77a
|
9 |
|
102 |
2 |
98abc
|
9 |
|
3 |
1 |
78bc
|
7 |
|
35 |
16 |
53g
|
5 |
|
7.33 |
|
Joyni
|
139 |
106 |
24cd
|
7 |
|
16 |
5 |
66j
|
7 |
|
23 |
19 |
18jk
|
3 |
|
1156 |
46 |
96bcd
|
9 |
|
3 |
2 |
15lm
|
1 |
|
103 |
16 |
84a
|
9 |
|
6 |
|
Bamonkhir
|
122 |
107 |
13jk
|
3 |
|
13 |
1 |
92ab
|
9 |
|
21 |
6 |
72b
|
9 |
|
194 |
1 |
99ab
|
9 |
|
3 |
1 |
77bc
|
7 |
|
68 |
24 |
64cd
|
7 |
|
7.33 |
|
Bouhari
|
125 |
107 |
14ij
|
3 |
|
10 |
1 |
90bc
|
9 |
|
22 |
6 |
71b
|
9 |
|
274 |
1 |
100a
|
9 |
|
3 |
1 |
79b
|
7 |
|
40 |
19 |
53g
|
5 |
|
7 |
|
Birindi
|
135 |
114 |
16hij
|
5 |
|
12 |
4 |
64j
|
7 |
|
19 |
18 |
5m
|
1 |
|
582 |
59 |
90fg
|
9 |
|
2 |
2 |
10no
|
1 |
|
49 |
28 |
42ij
|
5 |
|
4.67 |
|
BRRIdhan 41 |
115 |
103 |
10kl
|
1 |
|
11 |
9 |
15n
|
1 |
|
20 |
19 |
3mn
|
1 |
|
560 |
94 |
83ij
|
9 |
|
2 |
2 |
22k
|
3 |
|
17 |
16 |
6m
|
1 |
|
2.67 |
|
BRRIdhan 49 |
99 |
71 |
28ab
|
9 |
|
10 |
4 |
65j
|
7 |
|
24 |
16 |
33g
|
5 |
|
2063 |
203 |
90fg
|
9 |
|
2 |
1 |
70d
|
7 |
|
41 |
15 |
63de
|
7 |
|
7.33 |
|
BINAdhan 10 |
107 |
99 |
8l |
1 |
|
13 |
9 |
33l
|
3 |
|
26 |
21 |
21ij
|
3 |
|
1215 |
273 |
78k
|
7 |
|
2 |
2 |
14lm
|
1 |
|
57 |
23 |
60ef
|
7 |
|
3.67 |
|
Score range |
|
8-10 = 1
11-15 = 3
16-20 = 5
21-25 = 7
26-29 = 9 |
|
|
0-20 = 1
21-40 = 3
41-60 = 5
61-80 = 7
81-100 = 9 |
|
|
1-15 = 1
16-30 = 3
31-45 = 5
46-60 = 7
61-77 = 9 |
|
|
0-20 = 1
21-40 = 3
41-60 = 5
61-80 = 7
81-100 = 9 |
|
|
0-20 = 1
21-40 = 3
41-60 = 5
61-80 = 7
81-100 = 9 |
|
|
2-20 = 1
21-40 = 3
41-55 = 5
56-70 = 7
71-84 = 9 |
|
|
Table 3Genotypic performance of 38 rice germplasm using SSR markers related to salinity tolerance.
Table 3
|
Genotype |
Predicted salt tolerance response using SSR |
Genotype |
Predicted salt tolerance response using SSR |
|
|
|
RM 493 |
RM3412 |
RM 493 |
RM3412 |
|
Holde mota
|
T |
T |
Chorbindi
|
T |
T |
|
Dingamoni
|
T |
T |
Kajolshail
|
T |
T |
|
Dholkochua
|
S |
T |
Dudhkalom
|
T |
S |
|
Kedgrangal mota
|
T |
T |
Chaulmagi
|
S |
T |
|
Sadamota
|
T |
T |
Kutiakon
|
T |
T |
|
Laxima
|
T |
T |
Birindi
|
S |
S |
|
Tulsimala
|
T |
S |
Bamonkhir
|
T |
T |
|
Dishari
|
T |
T |
Kacha mota
|
T |
S |
|
Motha mota
|
S |
S |
Dudhmora
|
T |
T |
|
Lal chikon
|
T |
T |
Sorbimaloti
|
T |
T |
|
Kalijira
|
T |
T |
Sakhorkhora
|
T |
T |
|
Moulata
|
S |
S |
Chorboleshor
|
T |
S |
|
Calendar mota
|
T |
S |
Motha dhan
|
T |
T |
|
Lal mota
|
T |
S |
Bashful chikon
|
S |
S |
|
Bashful
|
T |
S |
Bushihara
|
T |
S |
|
Biyonti monkhusi
|
T |
S |
Kamola mota
|
T |
S |
|
Chinigura
|
S |
T |
Joyni
|
S |
T |
|
Kalo khaya
|
T |
T |
BRRIdhan 49 (S) |
S |
S |
|
Khaioz
|
T |
T |
BINAdhan 10 (T) |
T |
T |
References
- Abdullah Z, Khan MA, Flowers TJ. 2001. Causes of sterility in seed set in rice under salinity stress. J. Agron. Crop Sci.. 187: 25-32.
- Ali NM, Yeasmin L, Gantait S, Goswami R, Chakraborty S. 2014. Screening of rice landraces for salinity tolerance at seedling stage through morphological and molecular markers. Physiol. Mol. Biol. Plants. 20: 411-423.
- Babu NN, Vinod KK, Krishnan SG, Bhowmick PK, Vanaja T, Krishnamurthy SL, et al. 2014. Marker based haplotype diversity of Saltol QTL in relation to seedling stage salinity tolerance in selected genotypes of rice. Indian J. Genet.. 74: 16-25.
- Bangladesh Bureau of Statistics (BBS).2012. Statistical Yearbook of Bangladesh. Ministry of Planning, Govern-ment of the People's Republic of Bangladesh. pp. 33-36.
- Chunthaburee S, Dongsansuk A, Sanitchon J, Pattanagul W, Theerakulpisut P. 2016. Physiological and biochemical parameters for evaluation and clustering of rice cultivars differing in salt tolerance at seedling stage. Saudi J. Biol. Sci.. 23: 467-477.
- Desai AD, Rao TS, Hirekeru LR. 1975. Effect of saline water on growth and yield of rice. J. Indian Soc. Soil Sci.. 5: 13- 16.
- Ganie SA, Karmakar J, Roychowdhury R, Mondal TK, Dey N. 2014. Assessment of genetic diversity in salt-tolerant rice and its wild relatives for ten SSR loci and one allele mining primer of salT gene located on 1st chromosome. Plant. Syst. Evol.. 300: 1741-1747.
- Gregoria GB, Senadhira D, Mendoza RD. 1997. Screening rice for salinity tolerance. IRRI Discussion Paper Series No. 22. Manila (Philippines), International Rice Research Institute. pp. 1-30.
- Gregorio GB. 1997. Tagging salinity tolerant genes in rice using Amplified Fragment Length Polymorphism (AFLP). Ph.D. Thesis University of the Philippines Los Baños College. Laguna, Philippines, 118.
- Islam MR, Gregorio GB, Salam MA, Collard BCY, Singh RK, Hassan L. 2012. Validation of SalTol linked markers and haplotype diversity on chromosome 1 of rice. Mol. Plant Breed.. 3: 103-114.
- Islam MM. 2004. Mapping salinity tolerance genes in rice (Oryza sativa L.) at reproductive stage. Ph.D. Dissertation. University of the Philippines Los Baños College. Laguna, Philippines, 103-149.
- Javed MA, Ishii T, Kamijima O, Misoo S. 2006. Discrepancy of two ecotypes of Oryza sativa L. to salinity at germina-tion and seedling stages. Ann. Biol.. 22: 201-211.
- Jeung JU, Hwang HG, Moon HP, Jena KK. 2005. Fingerprinting temperate japonica and tropical indica rice genotypes by comparative analysis of DNA markers. Euphytica. 146: 239-251.
- Karan R, DeLeon T, Biradar H, Subudhi PK. 2012. Salt stress induced variation in DNA methylation pattern and its influence on gene expression in contrasting rice geno-types. PLoS One. 7: e40203
- Katerji N, Van Hoorn JW, Hamdy A, Mastrorilli M. 2003. Salinity effect on crop development and yield, analysis of salt tolerance according to several classification methods. Agric. Water Manag.. 62: 37-66.
- Khatun S, Flowers TJ. 1995a. Effects of salinity on seed set in rice. Plant Cell Environ. 18: pp. 61-67.
- Krishnamurthy SL, Sharma SK, Kumar V, Tiwari S, Singh NK. 2016. Analysis of genomic region spanning Saltol using SSR markers in rice genotypes showing differential seedlings stage salt tolerance. J. Plant Biochem. Biotechnol.. 25: 331-336.
- Marassi JE, Collado M, Benavidez R, Arturi MJ, Marassi JJN. 1989. Performance of selected rice genotypes in alkaline, saline and normal soils and their interaction with climate factors. Int. Rice Res. Newsl.. 14: 10-11.
- McCouch SR, Teytelman L, Xu Y, Lobos KB, Clare K, Walton M, et al. 2002. Development and mapping of 2240 new SSR markers for rice (Oryza sativa L.). DNA Res.. 9: 199- 207.
- Mojakkir AM, Tareq MZ, Mottalib MA, Hoque ABMZ, Hossain MA. 2015. Effect of salinity at reproductive stage in rice. Int. J. Bus.. 3: 7-12.
- Petersen L, Shireen S. 2001. Soil and water salinity in the coastal area of Bangladesh. Dhaka: Bangladesh Soil Re-source Development Institute (SRDI). Dhaka, Bangladesh.
- Rad HE, Aref F, Rezaei M. 2012. Response of rice to different salinity levels during different growth stages. Res. J. App. Sci. Eng. Technol.. 4: 3040-3047.
- Rahman M, Ahsan M. 2001. Salinity constraints and agri-cultural productivity in coastal saline area of Bangladesh, soil resources in Bangladesh: Assessment and utilization. In: Proceedings of the Annual Workshop on Soil Resources; 14-15 February 2001; Soil Resources Development Insti-tute, Dhaka. pp 1-14.
- Rashid MM, Hassan L, Begum SN. 2017. Phenotypic per-formance of rice landraces under salinity stress in repro-ductive stage. Progress. Agric.. 28: 1-6.
- Roy SK, Patra SK, Sarkar KK. 2002. Studies on the effect of salinity stress on rice (Oryza sativa L.) at seedling stage. J. Interacademicia. 6: 254-259.
- Rubel MH, Hassan L, Islam MM, Robin AHK, Alam MJ. 2014. Evaluation of rice genotypes under salt stress at the seedling and reproductive stages using phenotypic and molecular markers. Pak. J. Bot.. 46: 423-432.
- Saxena MK, Pandey UK. 1981. Physiological studies on salt tolerance of ten rice varieties growth and yield aspect. Indian J. Plant Physiol.. 24: 61-68.
- Singh RK, Mishra B. 1997. Stable genotypes of rice for sodic soils. Indian J. Genet.. 57: 431-438.
- Singh VP, Neue HU, Akbar M. Senadhira D., 1994. Coastal saline soils for rice cultivation. editor. Rice and problem soils in South and Southeast Asia. Discussion Paper Series No. 4. International Rice Research Institute (IRRI). Manila, Philippines: pp. 20-35.
- Tahjib-Ul-Arif M, Sayed MA, Islam MM, Siddiqui MN, Begum SN, Hossain MA. 2018. Screening of rice landraces (Oryza sativa L.) for seedling stage salinity tolerance using morpho-physiological and molecular markers. Acta Physiol. Plant. 40: 70
- Thomson MJ, de Ocampo M, Egdane J, Rahman MA, Sajise AG, Adorada DL, et al. 2010. Characterizing the Saltol quantitative trait locus for salinity tolerance in rice. Rice. 3: 148-160.
- Xu Y, Beachell H, McCouch SR. 2004. A marker-based ap-proach to broadening the genetic base of rice in the USA. Crop Sci.. 44: 1947-1959.
- Yoshida S, Forno DA, Cock JH, Gomez KA. 1976. Laboratory manual for physiological studies of rice, 3rd Edition. International Rice Research Institutes. Manila, Philippines: pp. 61.