Abstract
Cowpea (Vigna unguiculata (L.) Walp.) is one of the important legume crops in which all stages of plant growth are used in human food and animal feed. In this study, morphological and biochemical changes were investigated in cowpea plants exposed to proton-beams and gamma-rays. ‘Okdang’ cultivar seeds were irradiated with 100, 200, 300, 400, and 500 Gy of gamma-rays and proton-beams. The gamma-ray and proton-beam irradiations significantly delayed emergence period, but had small effects on emer-gence rates. Compared to a control, the proton-beam treatments showed a significant decrease in survival rate above 200 Gy. The gamma-ray treatments significantly reduced survival rate above 300 Gy. The Lethal Dose 50 (LD50) for both irradiations were 200-300 Gy. Plant height and fresh shoot weight tended to decrease with increasing radiation levels. The Reduction Dose 50 (RD50) of plant height was between 100 and 200 Gy for the proton-beam and 200 to 300 Gy for the gamma-ray treatments. The RD50 of fresh shoot weight was expected to be 500 Gy or higher for the proton-beam and gamma-ray treatments. The malonaldehyde (MDA) content in both treatments were slightly decreased at 200 Gy compared to a control, but increased at 300 Gy. Overall, antioxidant enzymes reacted more to proton-beams than to gamma-rays. However, gamma-rays appear to have a greater effect on chlorophyll content than proton-beams. Our research will be valuable as a basic research comparing the mutagenic effects on cowpea induced by two ionizing radiations.
-
Key words: Cowpea, Gamma-ray, Proton-beam, Radio sensitivity
INTRODUCTION
Cowpea (
Vigna unguiculata L. Walp.) is a widely cultivated legume in Asia, Latin America, and Africa (
Sombié et al. 2018). While cowpea is a major source of protein, it also contains vitamins, essential minerals, re-sistant starch, dietary fiber, and other beneficial components for human health (
Sreerama et al. 2012;
Nassourou et al. 2016). Cowpea protein can significantly reduce total and non-HDL cholesterol levels, which has helped in the treatment of hypercholesterolemia and steatosis (
Frota et al. 2008). The large amount of phenolic compounds in cowpea can also significantly remove and inhibit free radicals or reactive oxygen species (ROS), which are known to cause various diseases such as gastric ulcer, cardiovascular disease, diabetes, neurological disorders, and premature aging (
Xiong et al. 2013). Cowpea extracts activate pro-inflammatory mediators that may be helpful in treating inflammatory responses (
Sombié et al. 2018). Cowpea has high nutritional value; however, many important agronomic traits such as yield, plant height, and flowering time still need to be improved for consumers and pro-ducers.
Improving plant breeding crops requires a wide genetic diversity of useful agronomic traits (Novak and Brunner 1992). Mutation breeding enables the improvement of agri-cultural properties such as resistance to stress, increased content of useful components such as oils and proteins, and early maturation (
Oladosu et al. 2016). Therefore, plant breeding programs that use mutagens to induce numerous gene mutations are highly desirable. According to the mutant variety database (MVD), approximately 78% of all registered mutant varieties result from physical mutagens. Approximately 69% of these varieties were created by gamma-rays and about 22% by X-rays. These values indi-cate that ionizing radiations are the most common type of mutagens in mutation breeding. In cowpea, 16 mutant vari-eties were registered with improved characteristics such as higher yield, seed size, number of pods, and resistance to diseases by gamma-rays and dimethyl sulfate (DMS) mu-tagen.
Radiation mutation effects are related to linear energy transfer (LET), which is the amount of energy transferred per unit length of the track of an ionizing particle (
Zirkle and Tobias 1953;
Yokota et al. 2003). The LET of gamma-ray is 0.2 keV/µm, which can be changed by controlling the ion source and velocity (
Schmittgen and Livak 2008). He, C, N, Ne, and Ar ions were mainly used in this mutation breeding study and the LET ranges of these ions were 9-66, 22.5-309, 28.5-61.5, 61.5-549, 188-656 keV/µm, respec-tively (
Tanaka et al. 2010;
Abe et al. 2015). In particular, the LET value of a proton-beam (10 MeV-9,000 MeV) belonging to an ion beam is 0.23-4.6 keV/µm (
Yang 1999), which is within the range of a low LET that is similar to a gamma-ray, to a relatively high LET.
Plants are directly or indirectly damaged by ionizing radiation. Direct damages are caused by ionization or ex-citation of biomolecules through direct energy deposition, and indirect damages are caused by the generation of re-active oxygen species (ROS) that react with biomolecules (
Esnault et al. 2010;
Van Hoeck et al. 2017). ROS includes •OH molecules that are primarily produced by radiation ionizing water molecules and by H
2O
2 and O
2•
‒ molecules being secondarily produced through a chain reaction (
Esnault et al. 2010). Usually, ROS are constantly produced because of various metabolic pathways in plant cells and their or-ganelles, such as chloroplasts, mitochondria, and peroxisomes (
Garg and Manchanda 2009). ROS levels are physiologi-cally balanced in steady cells because the generated ROS are removed through the antioxidant defense system (
Foyer and Noctor 2005). However, environmental stresses such as ionizing radiation, salinity, drought, and mineral de-ficiencies, cause an imbalance between ROS production and antioxidant defense mechanisms that cause “oxidative stress” in cells (
Ahmad et al. 2008;
Goh et al. 2014). Cells in this state are severely damaged by the oxidation of lipids, proteins, and nucleic acids, which can lead to physiological effects that can affect plant growth and development (
Pitzschke et al. 2006;
Sharma et al. 2012). Oxidative stresses can be reduced by plant antioxidant enzymes such as ascorbate peroxidase (APX) and peroxidase (POD) (
Gill and Tuteja 2010).
The optimal dose of a radiation mutagen is estimated by a radio sensitivity test that indicates the degree of effect from radiation exposure (
van Harten 1998;
Mba et al. 2010). Studies on seed germination, plant survival, and seedling growth have been conducted to determine the morphological effects of radiation (
Ariharasutharsan et al. 2019;
Chauhan et al. 2019). Chlorophyll content is an im-portant parameter in determining physiological responses to radiation because of how it functions in the absorption and utilization of light energy and photosynthetic efficiency (
Kim et al. 2012a). Radiation induced-oxidative stress in plants can be monitored indirectly by measuring malon-aldehyde (MDA) levels, which is the end product of lipid peroxidation induced cell membrane damage, and the acti-vity of antioxidant enzymes (APX and POD) that can elim-inate ROS (
Wang et al. 2010;
Kim et al. 2015).
Today plant breeders are constantly striving to improve crop environmental adaptability, yield, and nutritional quality. Therefore, the continuous development of muta-tion induction techniques is essential for the generation of new genetic traits. This study investigated the morpho-logical and biochemical responses of cowpea to two dif-ferent ionizing radiations (gamma-rays and proton-beams) and determined the optimal doses for mutation induction.
MATERIALS AND METHODS
Plant materials and radiation treatments
Seeds of the cowpea cultivar ‘Okdang’ were obtained from the Jeollanamdo Agricultural Research and Extension Services (JARES) and used in this study. ‘Okdang’ is an erect plant with an intermediate plant habit that has a high lodging resistance (
Kim et al. 2018). The ‘Okdang’ seeds were irradiated with proton-beams and the gamma-rays.
Proton-beam irradiation: The seeds were irradiated with a 57 MeV proton-beam at 100 MeV using the proton linear accelerator at the Korea Multi-purpose Accelerator Com-plex (KOMAC) in Gyeongju, Korea. The ‘Okdang’ seeds were exposed to five different doses of proton-beam (100, 200, 300, 400, and 500 Gy). A total of 200 seeds were irradiated.
Gamma-ray irradiation: Gamma-ray irradiation was carried out using the low-level irradiation facility containing 60CO as a source at the Korea Atomic Energy Research Institute in Jeongeup, Korea. The irradiation doses were the same as the proton-beam irradiation. A total of 200 seeds were irra-diated.
Seedling emergence and survival
The irradiated M1 seeds and non-irradiated control seeds were planted in 50-cell plastic trays (27 × 53 × 11.2 cm) that were filled with a potting mix (coco peat, peat moss, zeolite, pearlite, caldolomite, a wetting agent, and fertilizer). Each seed was planted in an individual cell. Four replicates were performed for each treatment, and each replicate comprised a tray of 50 plants. The seedling emergence rates were calculated for each tray as the ratio of emerged seeds to the total number of sown seeds between 5 to 10 days after sowing. Seeds were considered to have emerged once their shoots emerged and were exposed to light (
Liu et al. 2017). The seedling survival rates were calculated for each tray as the ratio of surviving plants to the total number of plants four weeks after sowing.
Plant height and fresh shoot weight were measured four weeks after sowing. The samples were randomly selected from the center rows of each tray. Three replicates were performed for each treatment, and each replicate included 10 plants. Plant height is the length from the first node to the tip of plant, and fresh shoot weight is the weight of shoot above the first node.
Biochemical responses
Fresh leaves from irradiated and non-irradiated plants were used to determine MDA content, APX and POD activities, and chlorophyll content. Leaves were randomly selected four weeks after planting and stored at ‒80℃ for later biochemical analyses. The fresh leaf samples (0.2 g) were ground in liquid nitrogen using a mortar and pestle.
Measurement of malondialdehyde (MDA) content
The MDA content, an indication of the degree of lipid peroxidation, was measured using the trichloroacetic acid (TCA) method described by
Zhang et al. (2011). Briefly, fresh leaf samples (0.2 g) were homogenized with 1.5 mL 10% TCA for 30 minutes in a refrigerator. Samples were vigorously vortexed before centrifugation at 10,000 rpm for 20 minutes at 4℃ and supernatants were mixed with an equal volume of 0.67% 2-thiobarbituric acid. The mixtures were heated at 95℃ for 30 minutes followed by cooling on ice for 5 minutes and centrifugation at 10,000 rpm for 5 minutes at 4℃. The MDA concentration was calculated using the formula below by measuring the absorbance at 450, 532, and 600 nm using an Epoch microplate spectro-photometer (Bio Tek Instruments Inc., USA).
The total soluble protein contents in irradiated and non-irradiated plants were determined using the Bradford method (
Bradford 1976). A powdered sample of 0.2 g was place in a 2.0 mL tube and mixed with 1.2 mL of 0.2 M potassium phosphate buffer (pH 7.8 + 0.1 mM EDTA) and thoroughly homogenized, followed by centrifugation at 10,000 rpm for 20 minutes at 4℃. To determine the total soluble protein content, 2 µL of supernatant was added into 8 µL of ddH
2O and 190 µL of 1X dye reagent. The absor-bance at 595 nm was determined using a spectrophoto-meter (Bio Tek Instruments Inc., USA) and compared to a standard curve plotted using bovine serum albumin (BSA) (Bio-Rad Laboratories, Inc., USA) with standard concen-trations of 0, 125, 250, 500, 750, 1,000, 1,500, and 2,000 µg/mL. The total soluble protein content of the samples is ex-pressed in mg/g fresh weight (FW) of plant material.
APX activity was measured using a modified method described by
Elavarthi and Martin (2010). The reaction solution (200 µL) contained 50 mM potassium phosphate buffer (pH 7.0), 0.5 mM ascorbate, 0.1 mM EDTA, 0.5 mM H2O2, and 10 µL sample. Enzyme activity was measured using a UV spectrophotometer to detect a decrease in absorbance at 290 nm for 5 minutes due to oxidation of ascorbate. The extinction coefficient of ascorbate is 2.8 mM
‒1cm
‒1.
Unit Activity (Units/min/g FW) = [(change in absorbance / minute) × Vol. Total (µL)] / Ext. coeff. × Vol. sample (µL).
POD activity was measured using a modified method described by
Kim et al. (2012a). The reaction solution (200 µL) contained 100 mM potassium phosphate (pH 6.0), 20 mM pyrogallol, 10 mM H
2O
2, and 10 µL sample. Enzyme activity was measured as the change in absorbance at 420 nm for 1 minute due to formation of the pyrogallol dehydrogenation product. The extinction coefficient of purpuro-gallin at 420 nm is 12 mM
‒1cm
‒1.
Unit Activity (Units/min/g FW) = [(change in absorbance / minute) × Vol. Total (µL)] / Ext. coeff. × Vol. sample (µL).
Measurement of chlorophyll content
Chlorophyll content was measured using grounded fresh leaf samples (0.2 g) that were homogenized with 1.5 mL of 80% acetone in a 2 mL tube. The homogenates were kept overnight in the dark at 4℃ and centrifuged the next day at 3,000 rpm at 4℃ for 15 minutes. The supernatant was transferred to a new tube. The absorbance was measured using a UV spectrophotometer at 663 nm and 645 nm. The chlorophyll (a, b, and total) contents were calculated ac-cording to
Ni et al. (2009).
Three to four replicates from each treatment were used to assess seedling emergence, seedling survival, morphologi-cal responses, and biochemical responses. The results were subjected to an analysis of variance (ANOVA) using the SPSS version 25 software package (SPSS Inc., USA). Se-paration of means was performed using the Least Signifi-cant Difference (LSD) at P < 0.05.
RESULTS
Effects of gamma-ray and proton-beam irradiation on cowpea seedling emergence and survival
The emergence rates of the control and treatment groups irradiated with gamma-rays and proton-beams were con-tinuously examined 5 to 10 days after sowing (
Table 1). Five days after sowing, the emergence rates for both radi-ations significantly decreased as irradiation increased. However, the emergence rates were severely reduced in the proton-beam irradiation. All treatments, except for the 500 Gy dose, showed emergence rates over 90% seven days after sowing. The emergence rates for gamma-rays at the 500 Gy dose were lower than in proton-beam until eight days after sowing.
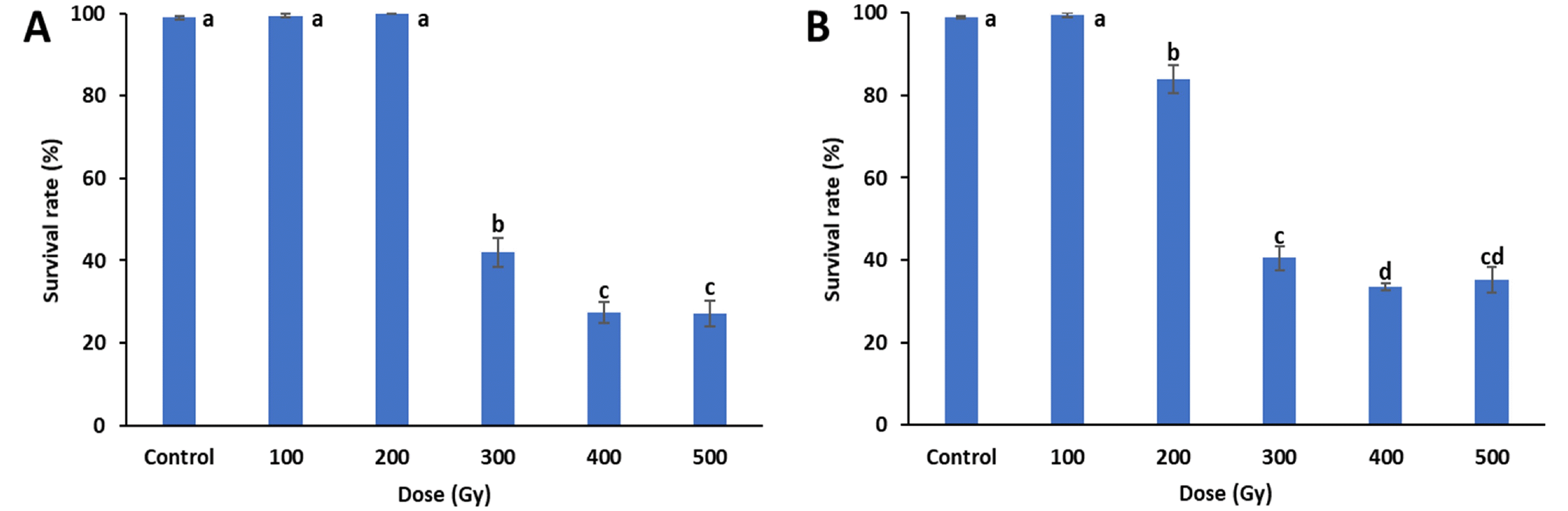
Survival rates were investigated in all treatments 25 days after sowing (
Fig. 1). The survival rates of both treatments decreased with increasing doses. When compared to the control, the proton-beam irradiations showed significant decreases in survival rates at and above 200 Gy, and the survival rates of the gamma-ray irradiations were signifi-cantly reduced at and above 300 Gy. In both treatments, the greatest reductions in survival rates were between 200 to 300 Gy, with gamma-rays decreasing from 100% to 42% and proton-beams decreasing from 84% to 41%. The proton beam and gamma-ray treatments at 500 Gy had survival rates of 35% and 27%, respectively. The lethal dose 50 (LD
50), which is the dose where 50% of the irradiated individuals survive, was between 200 and 300 Gy for both treatments.
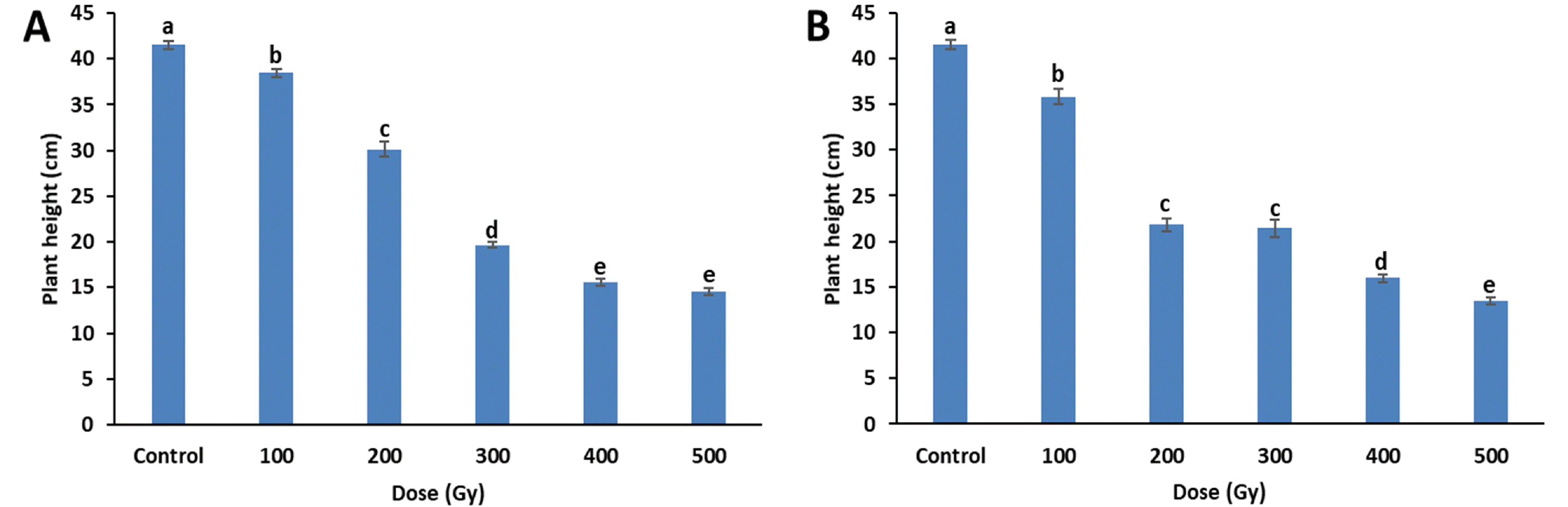
Four weeks after sowing, changes in growth parameters induced by gamma-rays and proton-beams irradiation were measured. When compared to the control, plant height significantly decreased with increasing gamma-ray and proton-beam doses (
Fig. 2). In particular, proton-beams at 200 Gy reduced plant height compared to effects of gamma rays at the same dose. At 500 Gy, plant height decreased by 65.1% (14.5 cm) and 67.7% (13.4 cm) for gamma-rays and proton-beams, respectively, compared to the control (41.5 cm).
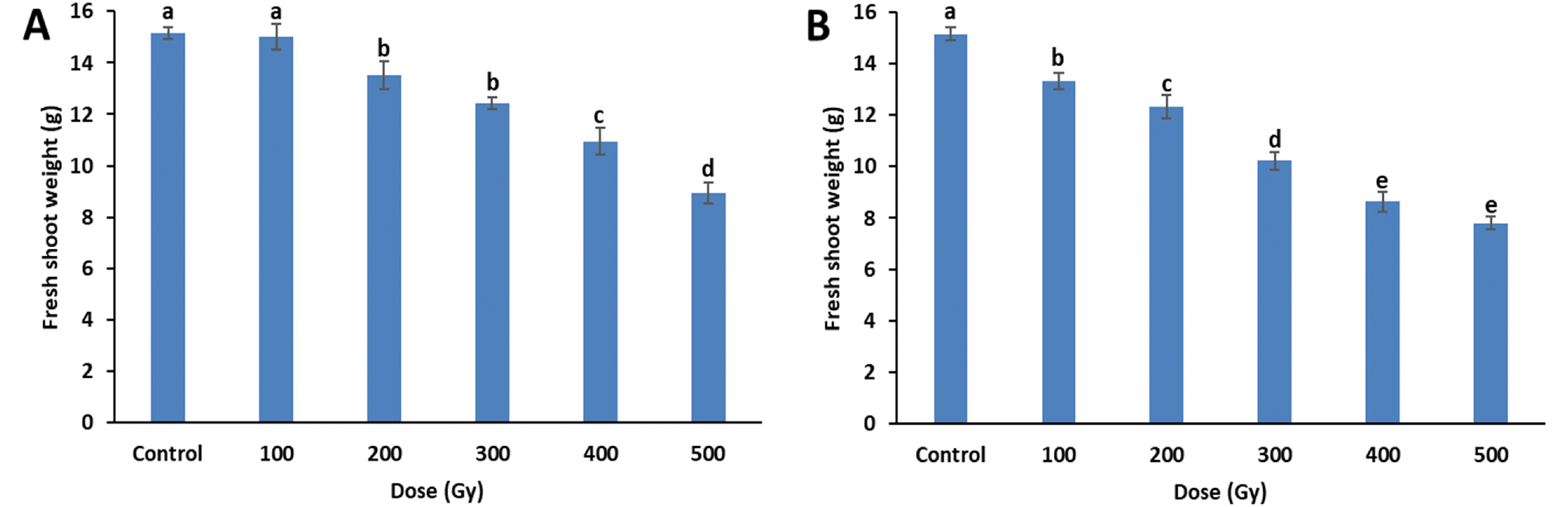
Similar to plant height, fresh shoot weight also decreased with increases in dose, regardless of the source, and the fresh shoot weight decreases were similar for both treat-ments (
Fig. 3). All doses of the proton-beam treatments resulted in lower fresh shoot weight than did the gamma-ray treatments at the same doses. Significant decreases between irradiated and control plants were observed from 100 Gy of proton-beams and 200 Gy of gamma-rays. At 500 Gy, fresh shoot weights for the gamma-ray and proton-beam treatments decreased by 40.8% (9.0 g) and 48.7% (7.8 g), respectively, compared to the control (15.2 g). Re-duction dose 50 (RD
50) for plant height was between 100-200 Gy for proton-beam and 200-300 Gy for gamma-ray treatments. However, the RD
50 of fresh shoot weight was 500 Gy or higher for both proton-beams and gamma-rays.
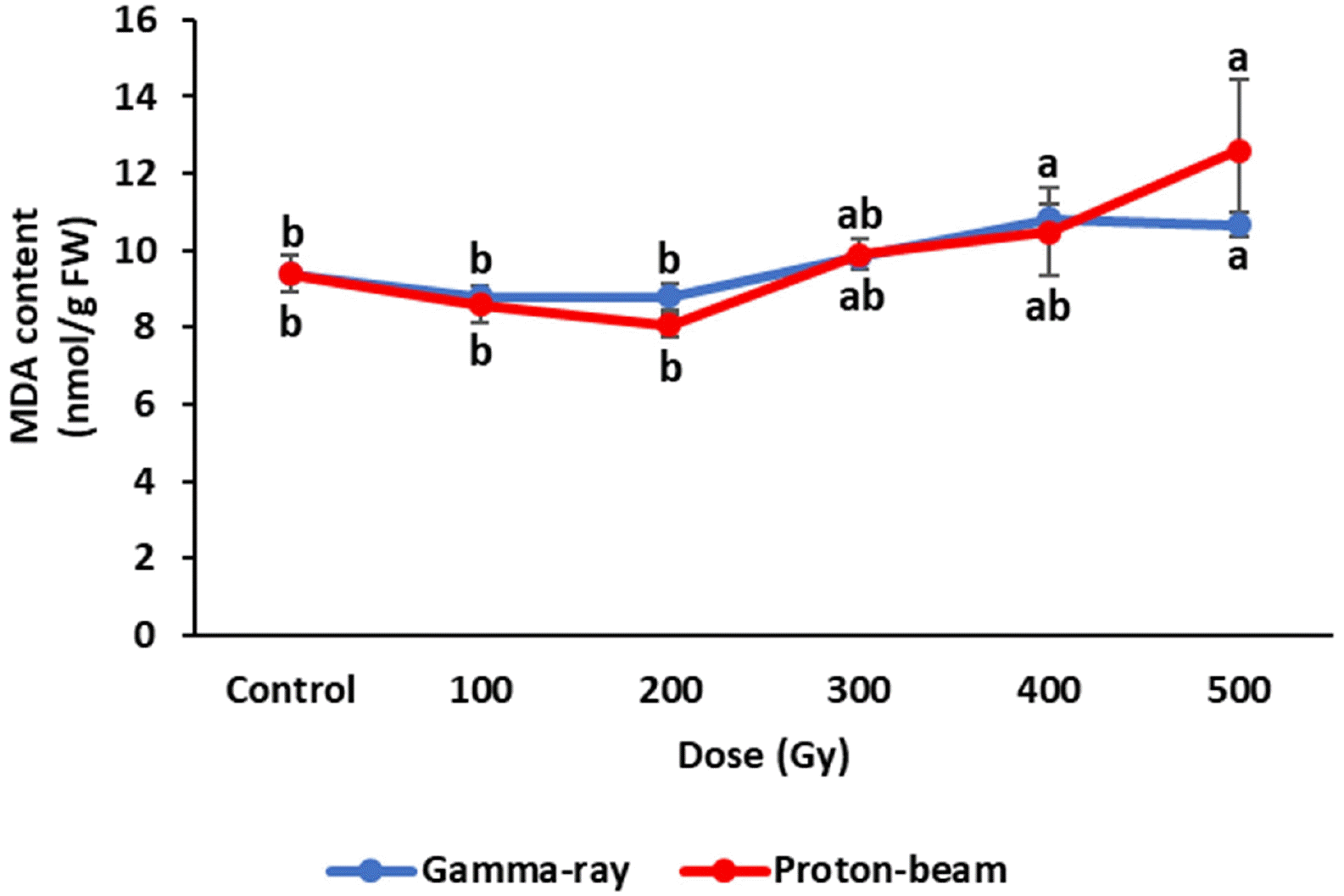
Biochemical responses of cowpea irradiated with gamma rays and proton-beams were compared with control plants four weeks after planting. The plant MDA contents in the gamma-ray and proton-beam treatments showed similar trends (
Fig. 4). MDA contents decreased slightly up to 200 Gy for both radiations compared to the control, but the decreases were not significantly different. However, MDA content increased above 200 Gy. The 500 Gy dose of gamma ray and proton-beam treatments increased MDA content 13.6% and 34.2%, respectively, compared to the control. The MDA content at 500 Gy for proton-beam was higher than that under other treatments.
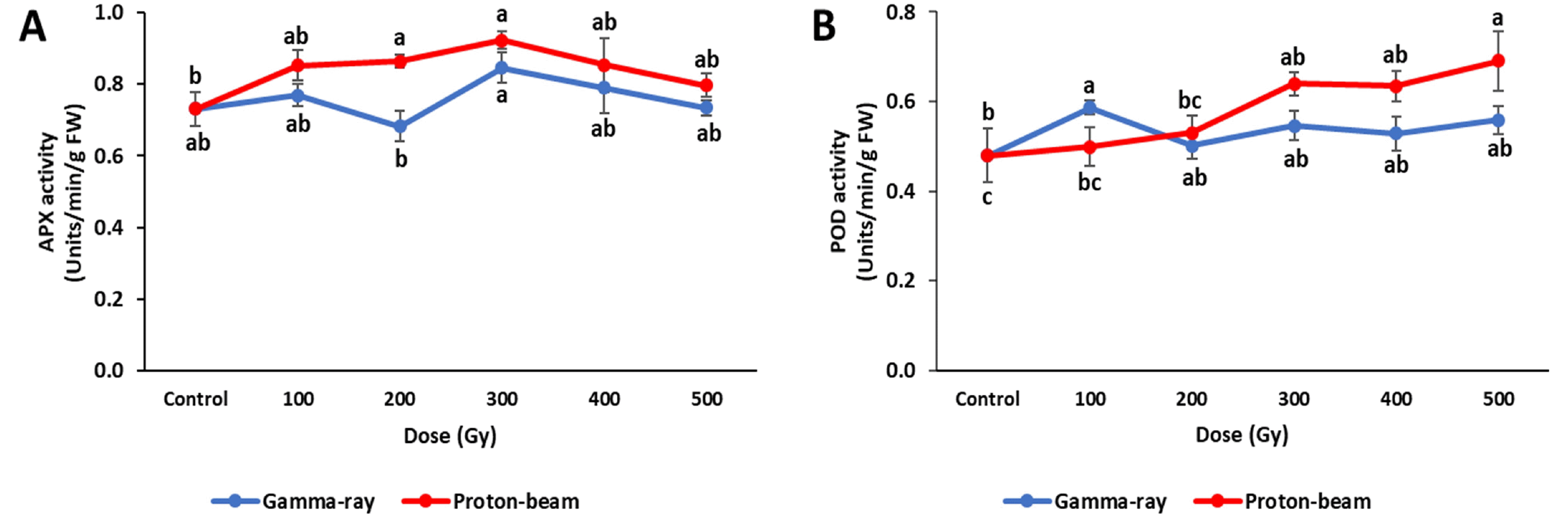
Cowpeas exposed to the different radiations showed variable antioxidant enzyme (APX and POD) activity re-sponses (
Fig. 5). APX activity in the proton-beam treat-ment increased in proportion with the dose up to 300 Gy, but decreased between 400 Gy and 500 Gy. The gamma-ray treatment showed results similar to that under the proton-beam treatment, except for a decrease at 200 Gy. All other gamma-ray and proton-beam doses showed higher APX activities than the control. APX activities at all gamma-ray doses were lower than activities at correspond-ing proton-beam doses.
POD activity in the proton-beam treatments significantly increased as the dose increased, with the highest value observed at 500 Gy (
Fig. 5). However, POD activity in the gamma-ray treatment showed a significant increase only at 100 Gy, with no significant changes observed at the other gamma-ray doses when compared to the control. The proton-beam treatments showed higher POD activities than gamma-ray treatments, except at the 100 Gy dose.
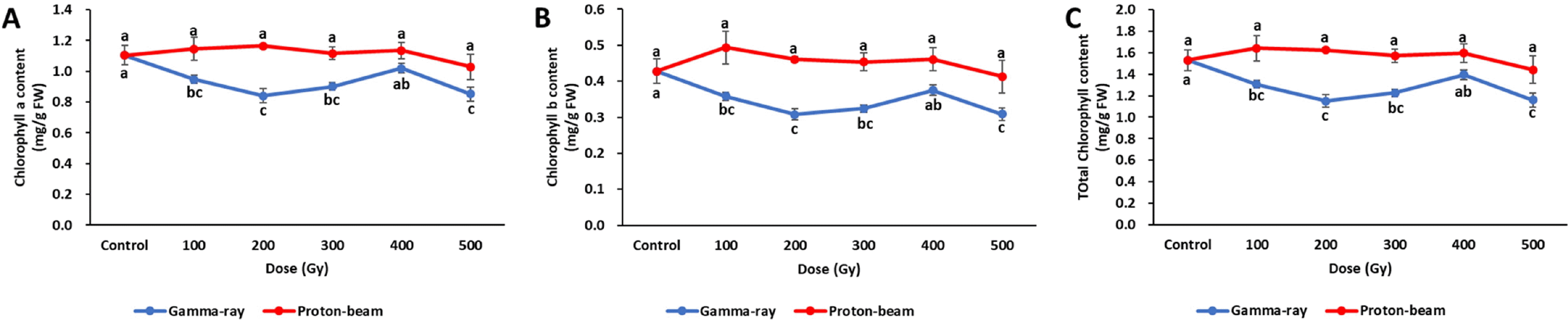
Changes in chlorophyll content by gamma-ray and proton beam irradiation were very different (
Fig. 6). In the gamma ray treatment, chlorophyll a, b, and total chlorophyll con-tents were the highest in the control, and decreased at all doses. Especially low amounts of chlorophyll a, b, and total chlorophyll contents in the gamma-ray treatment were observed at 200 Gy and 500 Gy. In the proton-beam treat-ment, unlike in the gamma-ray treatments, no significant differences were identified between any of the doses and the control. The chlorophyll a, b, and total chlorophyll con-tents for all gamma-ray doses were lower than those at the proton-beam doses.
DISCUSSION
Mutagenesis is very useful for expanding gene pools in plants. However, in addition to having positive effects, ionizing radiation can simultaneously cause negative ef-fects such as a reduction in germination rates, survival rates, growth rates, and fertility of plants. For these reasons, in mutation breeding the mutagenic effectiveness and efficiency should be considered when selecting a desired mutagen and dose (
Verma et al. 2017). Therefore, essential research on radio sensitivity for new mutagens such as proton-beams is needed. Unlike gamma-rays, which is an electromagnetic radiation, proton-beams are an ion beam composed of particles having mass and charge (
Oladosu et al. 2016). A proton-beam has a low LET value compared to other ion-beams and has been proposed as a new mutagen because it shows a different mutant spectrum (
Lee et al. 2015). However, there is little radio sensitivity information about proton-beams. Therefore, this study was conducted to evaluate the morphological and biochemical responses to proton-beam irradiation compared with those to gamma-rays, and to obtain information on LD
50 and RD
50 values for cowpea mutation breeding programs.
We first investigated the effects of both radiations at different doses on cowpea seedling emergence. Five days after sowing, the emergence rates decreased significantly as the dose increased. However, 10 days after sowing all treatments had emergence rates over 90%. Therefore, the final emergence rates were not significantly affected by proton-beam and gamma-ray irradiations, although proton-beam irradiation caused more emergence delays than did gamma-ray irradiation. Survival rates were similar to emer-gence rates and were significantly reduced by 200 Gy of proton-beams and 300 Gy of gamma-rays. Reductions in survival rates have been reported in various plants at high doses of proton-beams, gamma-rays, and carbon-ion beams (
Eun et al. 2007;
Takano et al. 2013;
Im et al. 2017a). In the current study, proton-beam irradiation had more severe effects on survival rates than gamma-ray irradiation, al-though both mutagens had similar LD
50 within a range of 200-300 Gy. Similarly, rice had more reduced LD
50 values from proton-beams (250 Gy) than gamma-rays (300 Gy) (
Yamaguchi et al. 2006;
Kim et al. 2012b). However, soybean had similar LD
50 values (around 300 Gy) between proton-beams and gamma-rays (
Im et al. 2017a;
Kusmiyati et al. 2018).
Plant height and fresh shoot weight of cowpeas irradi-ated with gamma-rays and proton-beams showed a similar decreasing pattern with increasing dose. Ionizing radiation typically decreases various growth parameters in crops regardless of the irradiation source (
Liu et al. 2013;
Im et al. 2017a;
Verma et al. 2017). These effects are due to the cell cycle stopping or pausing during the G2/M phase, reduced cell division, increased peroxide production, and damage to enzymes, growth hormones, and the entire genome (Preuss and Britt 2003; Kumar
et al. 2010;
Lagoda 2012). Studies have found increases in early growth pro-motion and grain yield from low gamma-ray or proton-beam doses in cowpea (
Olasupo et al. 2016), groundnut (
Tshilenge-Lukanda et al. 2013), and rape (
Eun et al. 2007). However, such increases were not found in the present study.
When comparing the two radiation treatments using the same dose, plant height and fresh shoot weight were more severely affected by the proton-beam treatment. Growth inhibition by proton-beams with a higher LET were greater than that by gamma-rays. Similarly, carbon-ion beams and helium-ion beams, which are high LET beams, required the same or lower doses to achieve the same biological effects on survival rate and fertility as gamma-rays in rice (
Yamaguchi et al. 2009). Based on the reduction of plant height, the RD
50 for plant height was between 100-200 Gy for proton-beams and 200-300 Gy for gamma-rays. However, the RD
50 for fresh shoot weight was 500 Gy or higher for both proton-beam and gamma-ray. These results showed that plant height had a more sensitive response to radiation than fresh shoot weight.
Typically, RD
30-RD
50 and LD
40-LD
60 were used to determine the optimal dose of radiation for mutation breed-ing (
Ryu et al. 2016). Based on these criteria, the optimal dose in cowpea was 200 Gy to 300 Gy for both irradiation sources. In addition, the shoulder dose (i.e., the dose at which survival rates start to decrease rapidly in a dose–survival rate curve) leads to efficient mutations with minimal effects on survival (
Yamaguchi et al. 2009;
Jo and Kim 2019). Based on these criteria, the optimal dose for cowpea was 300 Gy and 200 Gy for gamma-rays and proton beams, respectively.
Biochemical responses of cowpea to gamma-ray and proton-beam irradiation were relatively complex compared to plant growth. Non-stressed plants maintain a balance between ROS production and defense systems (
Foyer and Noctor 2005). However, ionizing radiation treatments in-crease the production of ROS in plants (
Kim et al. 2012a). Lipid peroxidation by high levels of ROS damages cell structures and impedes normal physiological functions (i.e. mitotic delays due to defective transport mechanism) (
Montillet et al. 2005;
Ünyayar et al. 2006;
Marcu et al. 2013). In the current study, cellular injury by lipid peroxidation was estimated using MDA content. Gamma-ray and proton beam treatments showed similar MDA tendencies of high radiation doses having higher MDA contents. These results are similar to findings of a study on soybeans irradiated with proton-beans, where MDA contents tended to in-crease with an increasing dose, or showed no significant changes (
Im et al. 2017b). There are two possible explan-ations. First, ROS generation was not increased at irradi-ation levels up to 200 Gy. Alternatively, any ROS increases were offset by a stimulation of the defense system (in-creased antioxidant enzyme activity). However, at doses at or above 300 Gy, the balance between ROS and the defense system was broken due to the defense system being damaged, or the ROS increase was too excessive for the defense system to match. At 500 Gy, the proton-beam treatment showed a higher MDA content than the gamma-ray treat-ment, suggesting that high doses of proton-beams induced more ROS generation than the gamma-rays, which caused more cellular damage.
ROS generated by ionizing radiation can be removed by defense mechanisms such as antioxidant enzymes (
Gill and Tuteja 2010). APX activity had a similar response to both ionizing radiations, but was higher at all radiation doses in the proton-beam treatments than the gamma radiation treat-ments. The increasing APX activity trend with higher doses changed around LD
50 (300 Gy), suggesting that radiation doses over LD
50 can damage the APX-related scavenging system and interfere with any responses in APX activity caused by dose. In addition, APX activity and MDA con-tent had opposite trends that indicate ROS content, which is not eliminated by the reduction of antioxidant enzyme activity, can lead to an increase in lipid peroxidation (MDA content). Another antioxidant enzyme, POD, responded differently to both ionizing radiations. There were no signi-ficant differences between gamma-ray treatments at all doses, except for 100 Gy and the control group. However, POD activities in the proton-beam treatments increased with dose. The responses of the two antioxidant enzymes indicate that the proton-beam irradiation induced more ROS production than did gamma radiation.
In the gamma-ray treatment, chlorophyll a, b, and total chlorophyll contents decreased at all doses when compared to the control. In contrast, no significant differences were found between all proton-beam doses and the control. This study found that the chlorophyll a, b, and total chlorophyll contents of cowpea were more affected by gamma-rays than proton-beams. These findings are consistent with pre-vious studies that reported gamma-irradiated plants had lower chlorophyll content than controls (
Ling et al. 2008;
Marcu et al. 2013).
In conclusions, cowpea cultivar ‘Okdang’ seeds irradi-ated with proton-beams had the same or greater responses in germination, survival, and plant growth than seeds irradiated with gamma-rays. MDA content and antioxidant activity, which indicates the degree of oxidative stress, were higher in the proton-beam treatments than in the gamma-ray treatments. However, the effects of gamma-rays on chlorophyll content appear to be greater than proton-beams. Based on the results in this study, the optimal dose of gamma-rays and proton-beams in cowpea mutation breeding is between 200-300 Gy. The current radio sensitivity study provides highly valuable infor-mation for using proton-beams and gamma-rays in mu-tation breeding.
ACKNOWLEDGEMENTS
This work was supported by Grants from the National Research Foundation of Korea (NRF) (NRF-2018M2A2A 6A05057264), Republic of Korea.
-
CONFLICT OF INTEREST
No potential conflict of interest relevant to this article was reported.
Fig. 1Mean survival rates of cowpea cultivar ‘Okdang’ seeds irradiated with various doses of gamma-rays (A) and proton-beams (B). Within each treatment, mean dose differences are indicated by bars with different letters that were determined by an LSD test at P = 0.05 (n = 3). The error bars indicate the standard error for n = 3 independent experiments.
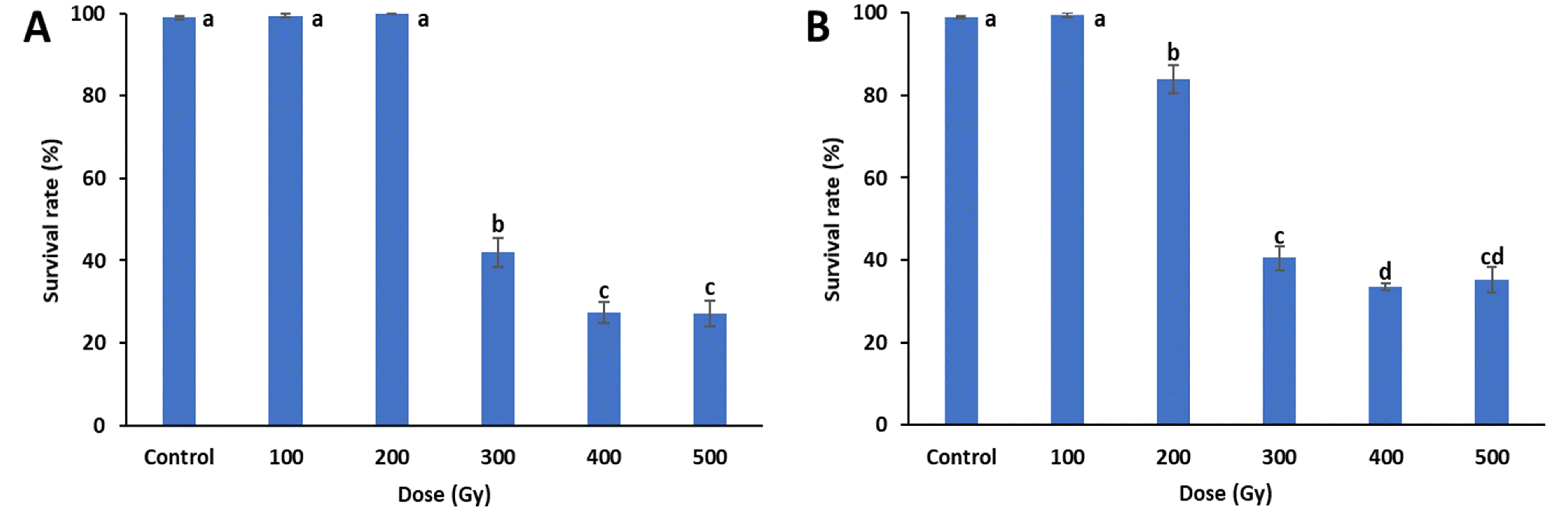
Fig. 2Plant height of cowpea cultivar ‘Okdang’ seeds irradiated with various doses of gamma-rays (A) and proton-beams (B). Within each treatment, mean dose differences are indicated by bars with different letters that were determined by an LSD test at P = 0.05 (n = 3). The error bars indicate the standard error for n = 3 independent experiments.
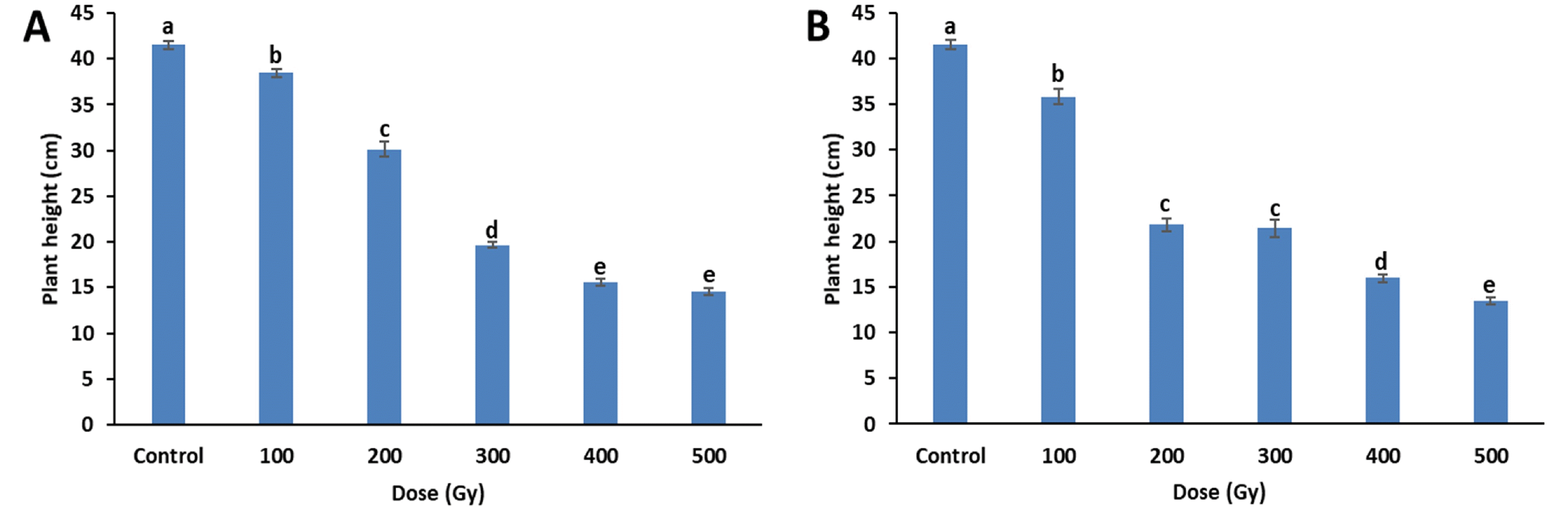
Fig. 3Fresh shoot weight of cowpea cultivar ‘Okdang’ seeds irradiated with various doses of gamma-rays (A) and proton-beams (B). Within each treatment, mean dose differences are indicated by bars with different letters that were determined by an LSD test at P = 0.05 (n = 3). The error bars indicate the standard error for n = 3 independent experiments.
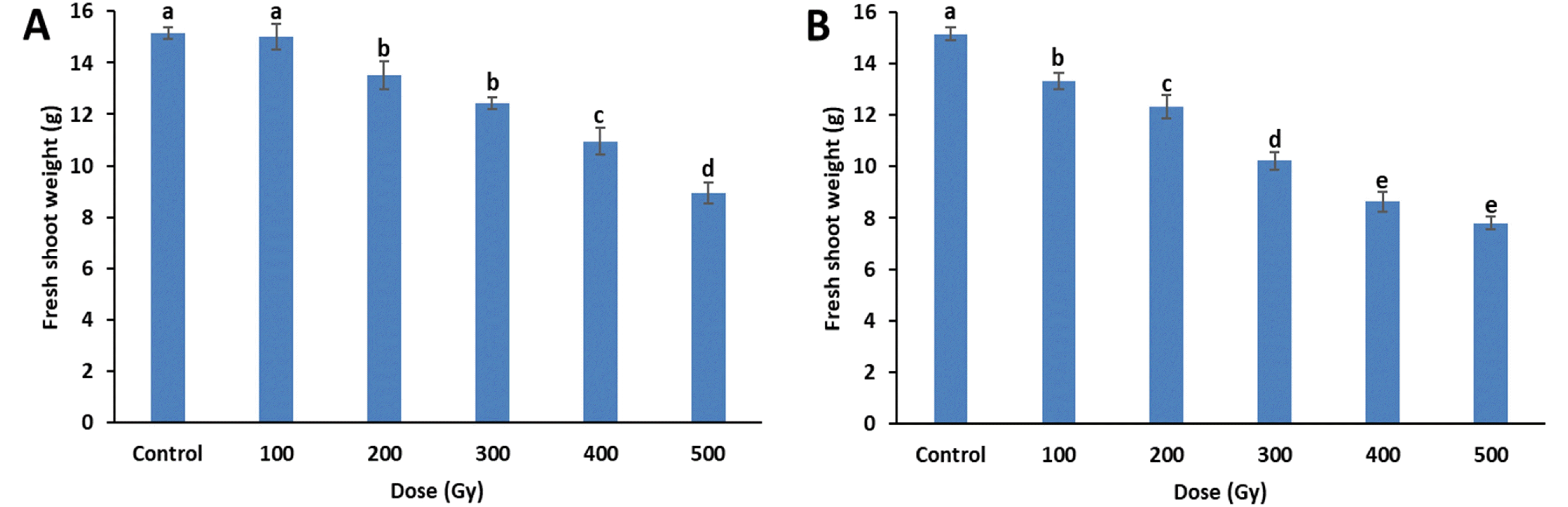
Fig. 4Antioxidant enzyme activities of cowpea cultivar ‘Okdang’ seeds irradiated with various doses of gamma-rays and proton-beams. (A) APX activity (B) POD activity. Mean dose differences are indicated by different letters that were determined by an LSD test at P = 0.05 (n = 3). The error bars indicate the standard error for n = 3 independent experiments.
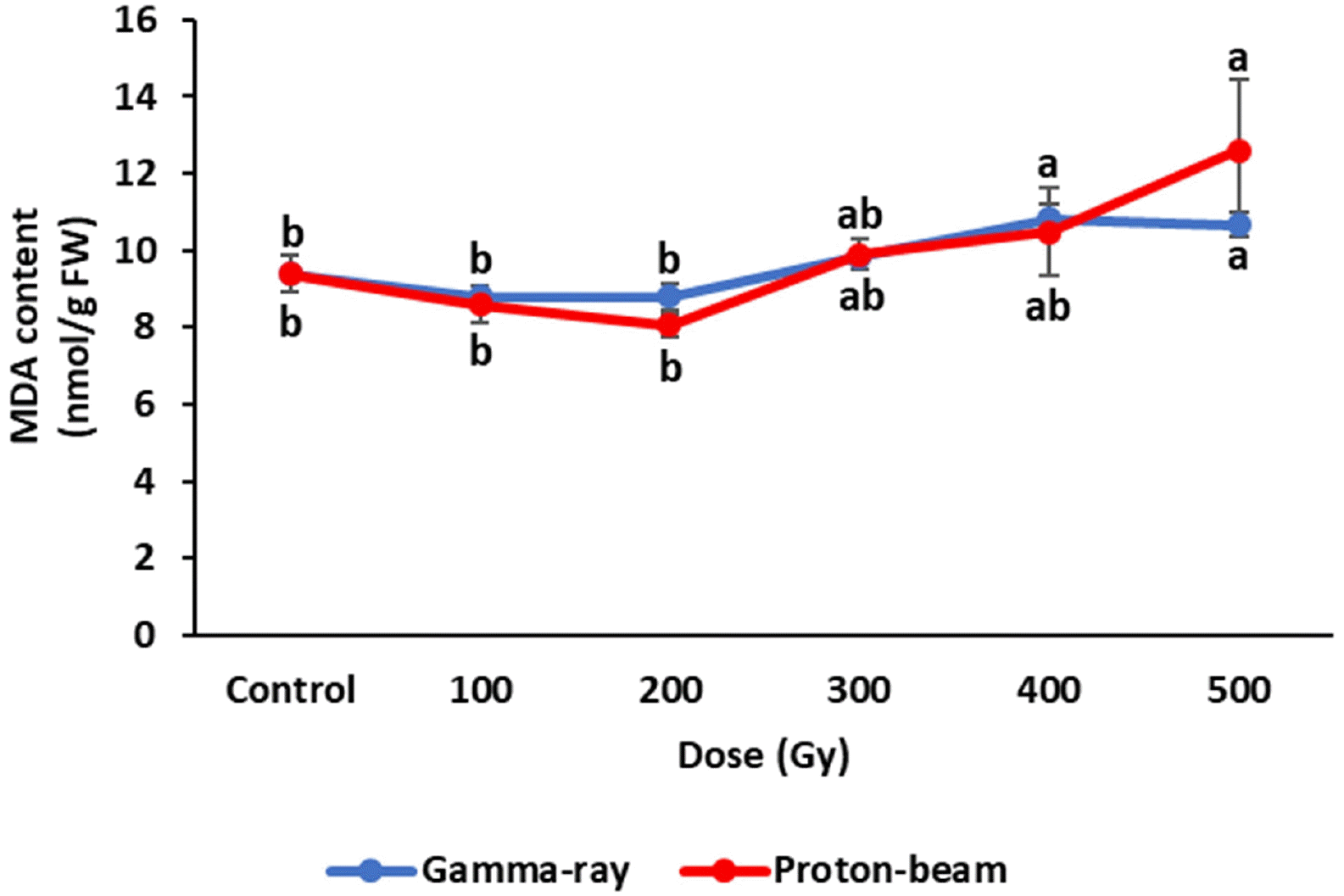
Fig. 5MDA contents of cowpea cultivar ‘Okdang’ seeds irradiated with various doses of gamma-rays and proton-beams. Mean dose differences are indicated by different letters that were determined by an LSD test at P = 0.05 (n = 3). The error bars indicate the standard error for n = 3 independent experiments.
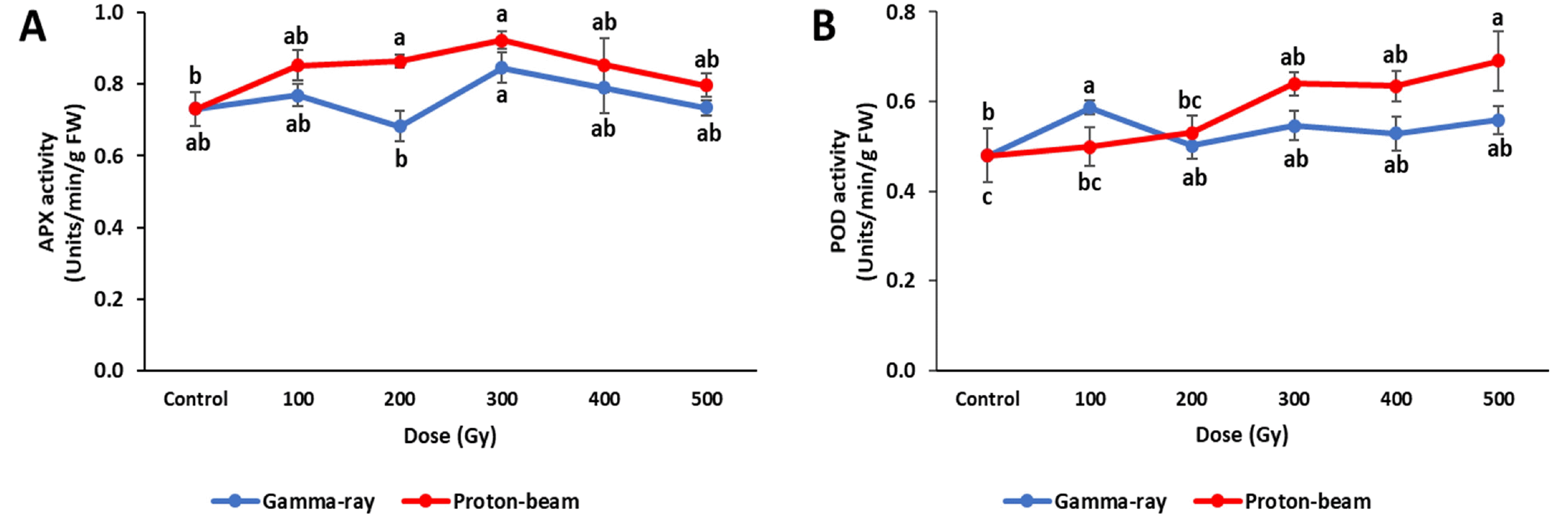
Fig. 6Chlorophyll contents of cowpea cultivar ‘Okdang’ irradiated with various doses of gamma-ray and proton-beam. (A) Chlorophyll A content (B) Chlorophyll B content (C) Total chlorophyll content. Mean dose differences are indi-cated by different letters that were determined by an LSD test at P = 0.05 (n = 3). The error bars indicate the standard error for n = 3 independent experiments.
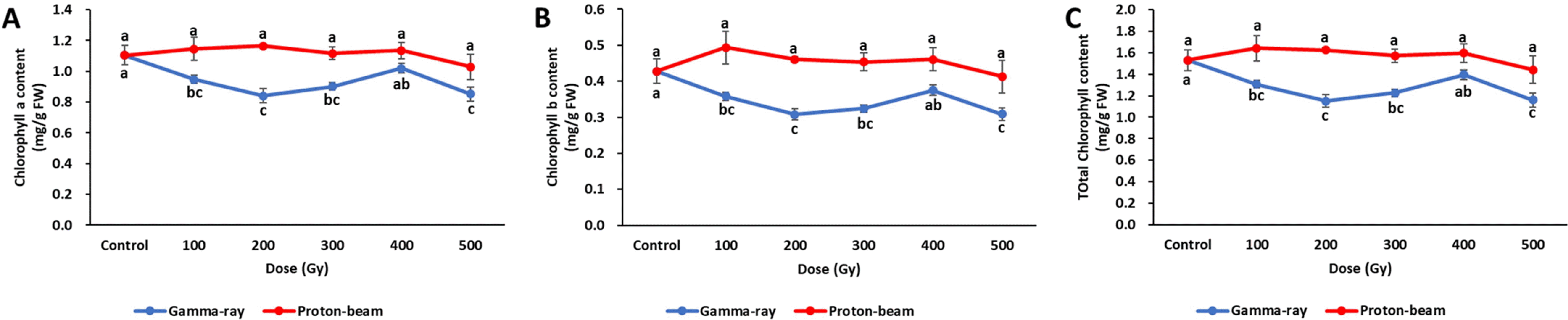
Table 1Mean emergence rates of cowpea cultivar ‘Okdang’ seeds irradiated with various doses of gamma-rays and proton-beams.
Table 1
Dose
(Gy) |
Emergence rate (%) |
|
|
gamma-ray |
|
proton-beam |
|
|
|
5 days |
6 days |
7 days |
8 days |
9 days |
10 days |
5 days |
6 days |
7 days |
8 days |
9 days |
10 days |
|
control |
93.25 a |
95.50 a |
95.50 a |
96.00 a |
96.75 a |
97.25 a |
|
93.25 a |
95.50 a |
95.50 ab |
96.00 ab |
96.75 ab |
97.25 a |
|
100 |
93.00 a |
95.00 ab |
95.50 a |
96.50 a |
96.50 a |
98.50 a |
|
88.00 a |
94.50 a |
98.00 a |
98.00 a |
98.50 a |
98.50 a |
|
200 |
89.50 a |
93.50 ab |
94.50 a |
95.00 a |
95.00 ab |
98.00 a |
|
51.00 b |
84.50 b |
93.00 ab |
95.00 ab |
97.00 ab |
96.50 ab |
|
300 |
40.00 b |
89.50 b |
93.00 a |
93.00 a |
93.50 ab |
94.00 ab |
|
26.00 c |
92.50 a |
96.00 ab |
96.50 ab |
96.50 ab |
96.50 ab |
|
400 |
23.00 c |
80.00 c |
93.00 a |
94.00 a |
94.50 ab |
95.00 ab |
|
12.00 d |
83.50 b |
90.50 b |
91.50 b |
92.00 bc |
92.50 bc |
|
500 |
15.50 d |
44.50 d |
70.00 b |
77.50 b |
90.50 b |
91.00 b |
|
16.00 d |
71.00 c |
82.00 c |
86.00 c |
88.50 c |
90.00 c |
References
- Abe T, Kazama Y, Hirano T. 2015. Ion beam breeding and gene discovery for function analyses using mutants. Nucl. Phys. News. 25: 30-34.
- Ahmad P, Sarwat M, Sharma S. 2008. Reactive oxygen species, antioxidants and signaling in plants. J. Plant Biol.. 51: 167-173.
- Ariharasutharsan G, Parameswari C, Vanniarajan C, Murugan E, Ramessh C. 2019. Radio sensitivity studies in white seeded sesame (Sesamum indicum L.). Electron. J. Plant Breed.. 10: 797-803.
- Bradford MM. 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem.. 72: 248-254.
- Chauhan A, Kumar V, Iyer PR, Vishwakarma G, Nair J, Surendran P, et al. 2019. Effect of proton beam ir-radiation on survival and seedling growth parameters of Indian rice (Oryza sativa L.) variety 'Indira Barani Dhan 1'. Electron. J. Plant Breed.. 10: 490-499.
- Elavarthi S, Martin B. Sunkar R., 2010. Spectrophotometric assays for antioxidant enzymes in plants. editor. Plant Stress Tolerance. Methods in Molecular Biology (Methods and Protocols), Vol. 639. Humana Press. pp. 273-280.
- Esnault MA, Legue F, Chenal C. 2010. Ionizing radiation: advances in plant response. Environ. Exper. Bot.. 68: 231- 237.
- Eun J, Kim J, Lim H, Han S, Choi S, Jang Y. 2007. Effect of proton ion and gamma-ray irradiation on radiosensitivity of M1 seedlings in Brassica napus. Korean J. Hortic. Sci. Technol.. 25: 17-23.
- Foyer CH, Noctor G. 2005. Redox homeostasis and anti-oxidant signaling: a metabolic interface between stress perception and physiological responses. Plant Cell. 17: 1866-1875.
- Frota K, Mendonça S, Saldiva P, Cruz R, Arêas J. 2008. Cholesterol‐lowering properties of whole cowpea seed and its protein isolate in hamsters. J. Food Sci.. 73: H235-H240.
- Garg N, Manchanda G. 2009. ROS generation in plants: boon or bane? Plant Biosyst.. 143: 81-96.
- Gill SS, Tuteja N. 2010. Reactive oxygen species and anti-oxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem.. 48: 909-930.
- Goh EJ, Kim JB, Kim WJ, Ha BK, Kim SH, Kang SY, et al. 2014. Physiological changes and anti-oxidative responses of Arabidopsis plants after acute and chronic γ-irradia-tion. Radiat. Environ. Biophys.. 53: 677-693.
- Im J, Kim WJ, Kim SH, Ha BK. 2017a. Effects of proton beam irradiation on seed germination and growth of soy-bean (Glycine max L. Merr.). J. Korean Phys. Soc.. 71: 752-757.
- Im J, Ko J, Kim HY, Ha BK. 2017b. Biochemical responses of soybean (Glycine max L. Merr.) to proton beam irra-diation. Plant Breed. Biotech.. 5: 97-105.
- Jo YD, Kim JB. 2019. Frequency and spectrum of radiation-induced mutations revealed by whole-genome sequencing analyses of plants. Quantum Beam Sci.. 3: 1-13.
- Kim DK, Choi JG, Kwon OD, Lee KD, Ryu KI. 2018. Cowpea cultivar, 'Okdang', with an intermediate plant habit and erect plant type. Korean J. Breed. Sci.. 50: 319-323.
- Kim DY, Hong MJ, Park CS, Seo YW. 2015. The effects of chronic radiation of gamma ray on protein expression and oxidative stress in Brachypodium distachyon. Int. J. Radiat. Biol.. 91: 407-419.
- Kim SH, Song M, Lee KJ, Hwang SG, Jang CS, Kim JB, et al. 2012a. Genome-wide transcriptome profiling of ROS scavenging and signal transduction pathways in rice (Oryza sativa L.) in response to different types of ionizing radiation. Mol. Biol. Rep.. 39: 11231-11248.
- Kim SK, Park SY, Kim KR, Shin JH, Kim SY, Kim HY, et al. 2012b. Effect of proton beam irradiation on germination, seedling growth, and pasting properties of starch in rice. J. Crop Sci. Biotech.. 15: 305-310.
- Kusmiyati F, Sas M, Herwibawa B. 2018. Mutagenic effects of gamma rays on soybean (Glycine max L.) germination and seedlings. IOP conference series: Earth and environ-mental science. IOP Publishing. Bristol, United Kingdom:
- Lagoda P. 2012. Effects of radiation on living cells and plants. Plant mutation breeding and biotechnology. pp. 123-134.
- Lee YM, Jo YD, Lee HJ, Kim YS, Kim DS, Kim JB, et al. 2015. DNA damage and oxidative stress induced by proton beam in Cymbidium hybrid. Hort. Environ. Bio-technol.. 56: 240-246.
- Ling APK, Chia JY, Hussein S, Harun AR. 2008. Physiolo-gical responses of Citrus sinensis to gamma irradiation. World Appl. Sci. J.. 5: 12-19.
- Liu B, Daryanto S, Wang L, Li Y, Liu Q, Zhao C, et al. 2017. Excessive accumulation of Chinese fir litter inhibits its own seedling emergence and early growth-a greenhouse perspective. Forests. 8: 1-11.
- Liu Q, Wang Z, Zhou L, Qu Y, Lu D, Yu L, et al. 2013. Relationship between plant growth and cytological effect in root apical meristem after exposure of wheat dry seeds to carbon ion beams. Nucl. Instrum. Methods Phys. Res., B. 305: 9-15.
- Marcu D, Damian G, Cosma C, Cristea V. 2013. Gamma radiation effects on seed germination, growth and pig-ment content, and ESR study of induced free radicals in maize (Zea mays). J. Biol. Phys.. 39: 625-634.
- Mba C, Afza R, Bado S, Jain SM. Davey MR., Anthony P., 2010. Induced mutagenesis in plants using physical and chemical agents. editors. Plant cell culture: Essential methods. John Wiley & Sons, Ltd.. Hoboken, New Jersey, U.S.A. pp. 111-130.
- Montillet JL, Chamnongpol S, Rustérucci C, Dat J, Van De Cotte B, Agnel JP, et al. 2005. Fatty acid hydroperoxides and H2O2 in the execution of hypersensitive cell death in tobacco leaves. Plant Physiol.. 138: 1516-1526.
- Nassourou MA, Njintang YN, Noubissié TJB, Nguimbou RM, Bell JM. 2016. Genetics of seed flavonoid content and antioxidant activity in cowpea (Vigna unguiculata L. Walp.). Crop J.. 4: 391-397.
- Ni Z, Kim ED, Ha M, Lackey E, Liu J, Zhang Y, et al. 2009. Altered circadian rhythms regulate growth vigour in hybrids and allopolyploids. Nature. 457: 327-331.
- Oladosu Y, Rafii MY, Abdullah N, Hussin G, Ramli A, Rahim HA, et al. 2016. Principle and application of plant mutagenesis in crop improvement: a review. Biotechnol. Biotechnol. Equip.. 30: 1-16.
- Olasupo FO, Ilori CO, Forster BP, Bado S. 2016. Mutagenic effects of gamma radiation on eight accessions of Cowpea (Vigna unguiculata [L.] Walp.). Am. J. Plant Sci.. 7: 339- 351.
- Pitzschke A, Forzani C, Hirt H. 2006. Reactive oxygen species signaling in plants. Antioxid. Redox Sign.. 8: 1757-1764.
- Ryu J, Kwon SJ, Im SB, Jeong SW, Ahn JW, Kim JB, et al. 2016. Effects of gamma-ray irradiation on radio sensiti-vity in oat (Avena sativa). Korean J. Plant Res.. 29: 128- 135.
- Schmittgen TD, Livak KJ. 2008. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc.. 3: 1101- 1108.
- Sharma P, Jha AB, Dubey RS, Pessarakli M. 2012. Reactive oxygen species, oxidative damage, and antioxidative de-fense mechanism in plants under stressful conditions. J. Bot.. 2012: 1-26.
- Sombié PAED, Compaoré M, Coulibaly AY, Ouédraogo JT, Tignégré J-BDLS, Kiendrébéogo M. 2018. Antioxidant and phytochemical studies of 31 cowpeas (Vigna unguiculata (L. Walp.)) genotypes from Burkina Faso. Foods. 7: 1-9.
- Sreerama YN, Sashikala VB, Pratape VM, Singh V. 2012. Nutrients and antinutrients in cowpea and horse gram flours in comparison to chickpea flour: Evaluation of their flour functionality. Food Chem.. 131: 462-468.
- Takano N, Takahashi Y, Yamamoto M, Teranishi M, Yamaguchi H, Sakamoto AN, et al. 2013. Isolation of a novel UVB-tolerant rice mutant obtained by exposure to carbon-ion beams. J. Radiat. Res.. 54: 637-648.
- Tanaka A, Shikazono N, Hase Y. 2010. Studies on biological effects of ion beams on lethality, molecular nature of mutation, mutation rate, and spectrum of mutation phe-notype for mutation breeding in higher plants. J. Radiat. Res.. 51: 223-233.
- Tshilenge-Lukanda L, Kalonji-Mbuyi A, Nkongolo K, Kizungu R. 2013. Effect of gamma irradiation on morpho-agronomic characteristics of groundnut (Arachis hypogaea L.). Am. J. Plant Sci.. 4: 2186-2192.
- Ünyayar S, Celik A, Çekiç FÖ, Gözel A. 2006. Cadmium-induced genotoxicity, cytotoxicity and lipid peroxidation in Allium sativum and Vicia faba. Mutagenesis. 21: 77-81.
- van Harten AM. 1998. Mutation breeding: theory and prac-tical applications. Cambridge University Press. Cambridge, United Kingdom..
- Van Hoeck A, Horemans N, Nauts R, Van Hees M, Vandenhove H, Blust R. 2017. Lemna minor plants chronically exposed to ionising radiation: RNA-seq analysis indicates a dose rate dependent shift from acclimation to survival strat-egies. Plant Sci.. 257: 84-95.
- Verma AK, Sharma S, Kakani RK, Meena RD, Choudhary S. 2017. Gamma radiation effects seed germination, plant growth and yield attributing characters of fennel (Foeniculum vulgare Mill.). Int. J. Curr. Microbiol. Appl. Sci.. 6: 2448- 2458.
- Wang YC, Qu GZ, Li HY, Wu YJ, Wang C, Liu GF, et al. 2010. Enhanced salt tolerance of transgenic poplar plants expressing a manganese superoxide dismutase from Tamarix androssowii. Mol. Biol. Rep.. 37: 1119-1124.
- Xiong S, Yao X, Li A. 2013. Antioxidant properties of peptide from cowpea seed. Int. J. Food Prop.. 16: 1245-1256.
- Yamaguchi H, Hase Y, Tanaka A, Shikazono N, Degi K, Shimizu A, et al. 2009. Mutagenic effects of ion beam irradiation on rice. Breed. Sci.. 59: 169-177.
- Yamaguchi H, Morishita T, Degi K, Tanaka A, Shikazono N, Hase Y. 2006. Effect of carbon-ion beams irradiation on mutation induction in rice. Plant Mutation Reports. 1: 25-27.
- Yang TC. 1999. Proton radiobiology and uncertainties. Radiat. Meas.. 30: 383-392.
- Yokota Y, Hase Y, Shikazono N, Tanaka A, Inoue M. 2003. LET dependence of lethality of carbon ion irradiation to single tobacco cells. Int. J. Radiat. Biol.. 79: 681-685.
- Zhang L, Xiao S, Li W, Feng W, Li J, Wu Z, et al. 2011. Overexpression of a Harpin-encoding gene hrf1 in rice enhances drought tolerance. J. Exp. Bot.. 62: 4229-4238.
- Zirkle RE, Tobias CA. 1953. Effects of ploidy and linear energy transfer on radiobiological survival curves. Arch. Biochem. Biophys.. 47: 282-306.