Abstract
Pre-harvest sprouting (PHS) is one of the important traits that not only cause serious economic issues but also lead to reduction in grain quality and yield in rice (Oryza sativa L.). To analyze the quantitative trait loci (QTLs) for PHS tolerance, we evaluated PHS, seed dormancy (SD), and low-temperature germination (LTG) of 88 F2:3 populations and their parental lines. Genotypic analysis was performed by using 441 single nucleotide polymorphisms (SNPs) detected from re-sequencing data. Seed dormancy (SD) and low-temperature germination (LTG) were identified to exhibit a positive correlation with PHS. Under the field condition, two major QTLs for PHS, qPHS1-1FC and qPHS1-2FC were identified on chromosome 1. Under the growth chamber condition, qPHS1-1GC and qPHS1-2GC had the same regions on chromosome 1. QTLs of SD and LTG (qSD1-1, qSD1-2, qLTG1-1, and qLTG1-2) had the same regions; these results suggested that candidate QTLs demonstrate pleiotropy about PHS, SD, and LTG. The major QTLs detected in this study are hypothesized to provide an important resource for molecular breeding and gain a better understanding of the genetics of traits in rice.
-
Key words: Pre-harvest sprouting, Whole genome resequencing, Recombinant inbred lines, Rice, QTL
INTRODUCTION
Pre-harvest sprouting (PHS) of rice (
Oryza sativa L.) is one of the most important traits because of serious economic issues that have led to a significant reduction in grain quality and grain yield. Seed dormancy (SD) prevents seedling emergence in the wrong season or place in wild species, in which the lack of dormancy results in PHS, the phenomenon of seed germination of the rice panicle or cereal grains before harvest. When matured rice panicles are not harvested in time or harvested rice grains are not dried immediately after harvesting, and rice cultivars with high PHS are subjected to germination by frequent rainfall and high temperatures (
Gubler et al. 2005;
Chono et al. 2006;
Wan et al. 2006;
Tejakhod and Ellis 2018). The weather is changing in Korea due to typhoons and unexpectedly high temperatures and heavy rains are more frequent during the rice maturing stage that causes an increase in PHS (
Lee et al. 2016).
Seed germination is very important for the success of crop production (
Almansouri et al. 2001). Among the associate traits about seed germination, the effects of PHS, SD, and low-temperature germination (LTG) are extremely complex, involving various physical and biochemical quality. These traits are expressed as quantitative traits under complex genetic control. The germination associated genes were reported to be related to abscisic acid (ABA) and gibberellic acid (GA) that are major signaling molecules involved in germination induction. ABA has demonstrated that endospermic sugars act as an essential energy source for seed germination and determine seed dormancy and germination by affecting ABA signaling (
Du et al. 2018). GA promotes germination, which requires not only initiating the activity of the embryo, but also breaking the physical barrier of the seed coat surrounding the embryo (
Debeaujon and Koornneef 2000). Several studies have considered SD as the major component of PHS (
Li and Foley 1997;
Gu et al. 2003;
Chen et al. 2008;
Fang et al. 2008;
Gu et al. 2011;
Graeber et al. 2012;
Ye et al. 2015).
Improving PHS tolerance for rice is one of the major breeding targets (
Zhang et al. 2014). To overcome PHS in rice under unpredictable weather changes, many genes and alleles that are associated with PHS must be identified. In an attempt to face the challenge to incorporate PHS tolerance into commercial rice cultivars, herein, we conducted QTL analysis with the F
2:3 population derived from the cross between high-quality japonica varieties. It is hypo-thesized that the QTLs detected in this study can be used directly in commercial rice breeding programs.
MATERIALS AND METHODS
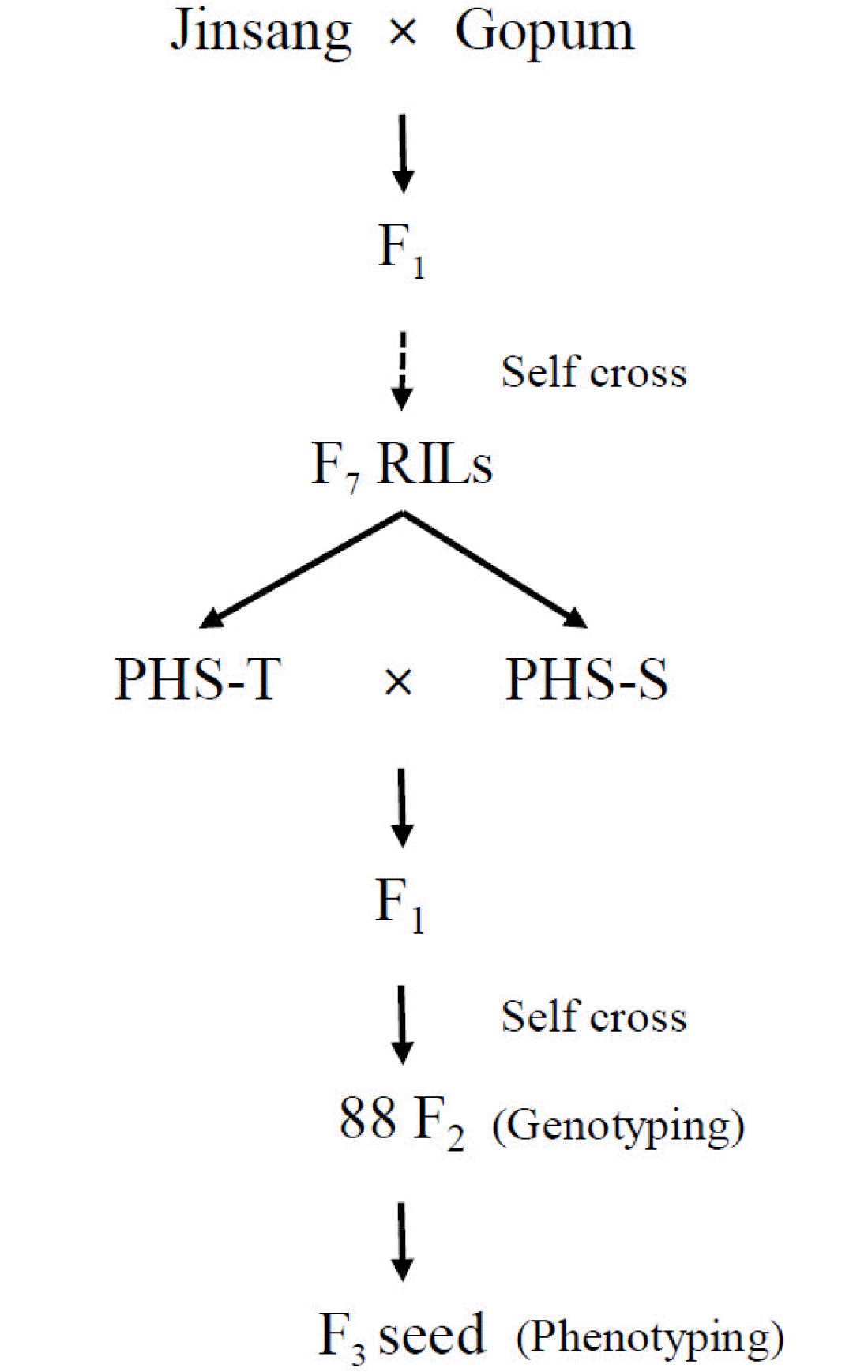
Plant materials
PHS tolerance line (PHS-T) and PHS susceptible line (PHS-S) were selected from the RILs derived from a cross between ‘Jinsang’, a parent with high-quality rice (
Hong et al. 2014) and ‘Gopum’ with high-quality rice but susceptible to PHS (
Kang et al. 2018). For QTL analysis, the 88 F
2 population was developed from a cross between PHS-T and PHS-S. PHS-T, PHS-S, and 88 F
2 plants were grown in the experimental field at Pusan National University in 2018 (
Fig. 1).
On 45 days after heading, three panicles from parent and 88 F
2 population were evaluated for PHS tolerance under the chamber condition. The harvested panicles of each line were wrapped in paper towels and placed in water at 100% relative humidity in the chamber at 20℃ for 7 days (
Gao et al. 2008). The three panicles of each plant were evaluated for PHS tolerance under field condition at 52 days after heading (
Supplementary Table S1). Each panicle from chamber and field conditions were evaluated for PHS by checking the germination rate. All statistical analyses were conducted using the Statistical Analysis System (SAS) software version 9.4 for Windows.
To evaluate SD and LTG, high-quality seeds were selected by the removal of shriveled and unfilled seeds. Fifty seeds with two times from each plant harvested at 45 days after heading were immediately placed in a petri dish (diameter 9 cm) with two sheets of moistened filter paper. The seeds were germinated under two conditions for the germination test in a growth chamber at 13℃ for 15 days and 28℃ for 7 days, respectively. Seed dormancy was evaluated from the germination rate at 28℃. The germi-nation rate of each selected individual was scored as a percentage of germinated seeds in the total numbers of seeds (
Lin et al. 1998;
Gu et al. 2003;
Cruz and Milach 2004;
Gu et al. 2011;
Hyun et al. 2017).
Total genomic DNA was extracted from young leaves of PHS-T, PHS-S, and 88 F
2 lines, using modified cetyl trimethyl ammonium bromide (CTAB) protocol (
Porebski et al. 1997). Whole-genome re-sequencing of the materials was performed on an MGISEQ-2000 platform (MGI, Shenzhen, China). The raw SNPs were filtered out by alignment with the Nipponbare reference genome (IRGSP-1.0) by BWA and GATK tools. To ensure high-confidence SNP markers for genetic map construction, all raw SNPs identified between the parental lines were filtered as follows: depth at each SNP in both the parental lines > 10 and mapping quality of reads > 20. We further selected SNPs which were 100% homozygous alleles in each parental line. The final candidate SNPs were used for the geno-typing of the F
2 population. To ensure high quality, reads of the F
2 population were filtered as follows: read depth > 10 and mapping quality of reads > 20. SNPs that had > 50% missing or ambiguous data were excluded. Subsequently, selected SNPs were filtered such that the distance between SNPs was 100K bp; the remaining high-quality data were used in the mapping.
Linkage maps were constructed from the genotype data by using version 4.1 of QTL IciMapping (
Meng et al. 2015). Genetic distances were estimated by using the software’s Kosambi map function (
Kosambi 1944). QTL IciMapping was also used with the phenotype data for 4 traits for QTL analysis. The inclusive composite interval mapping for additive QTL (ICIM-ADD) and inclusive composite interval mapping for epistatic QTL (ICIM-EPI), along with other default parameters, were used. The LOD threshold for ICIM-ADD and ICIM-EPI were set by using permutation 1000 and manual LOD value 5, respectively.
RESULTS
Phenotypic analysis of preharvest sprouting
The PHS rate of PHS-T, PHS-S, and F
2:3 populations were surveyed by involving three panicles under growth chamber and field conditions. F
2:3 population showed obvious segregation of tolerance and susceptibility, while agricultural traits such as plant height, heading date, culm length, and panicle length demonstrated uniformity in F
2 and F
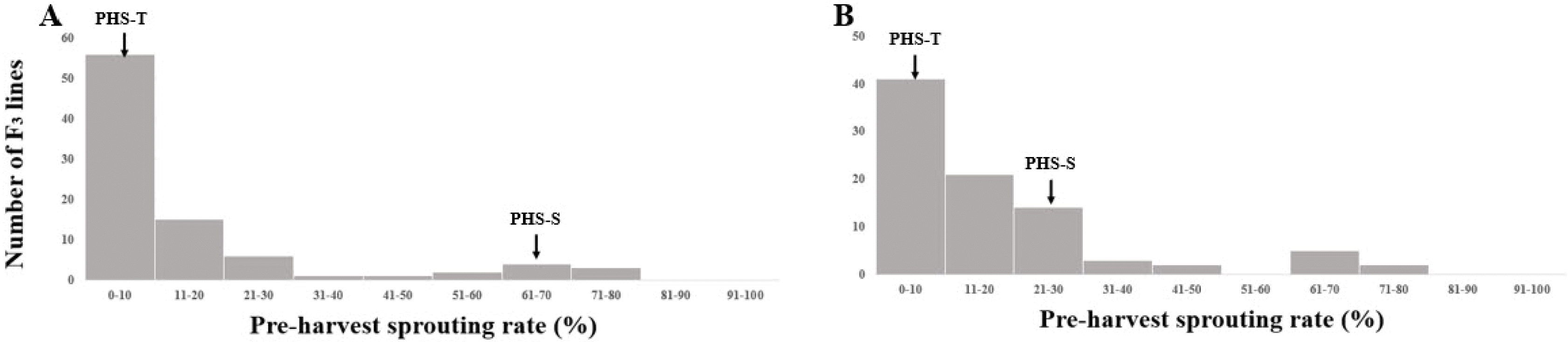
3 populations. The PHS rate of PHS-T displayed lower than 10% and PHS-S displayed 70.2% and 36.1% under growth chamber and field conditions, respectively (
Fig. 2). PHS rate of the F
3 population had a mean of 16.9% with a range from 0.6% to 79.8% under chamber condition, which had a mean of 13.7% with a range from 0% to 77.9% under field condition (
Fig. 3).
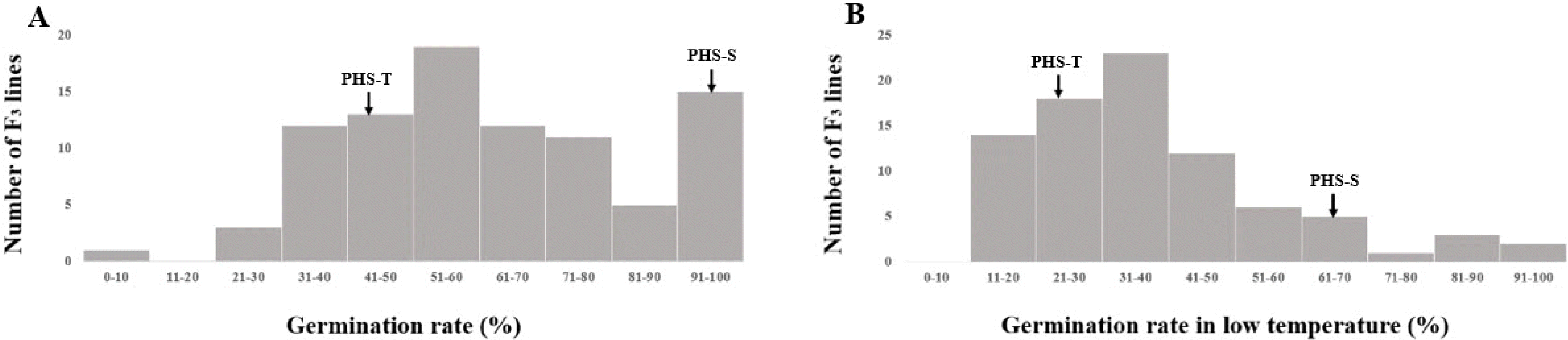
The germination rate at 28℃ of two parents, and F
2:3 populations were surveyed by fifty seeds per panicle. The germination rate at 28℃ of PHS-T and PHS-S were 41.44% and 98.61%, respectively. The germination rate of the F
3 population had a mean of 62.49% with a range from 9.17% to 100%. The LTG rate of PHS-T indicated 25.67% and PHS-S resulted in 69.46%. LTG rate of the F
3 population had a mean of 38.01% with a range from 7.49% to 95.19% (
Fig. 4).
A high variability was observed among the F
2:3 populations for PHS, SD, and LTG traits (
Table 1). There was significant correlation between each PHS and SD (
r = 0.64,
r = 0.57,
P-value < 0.01). Also, there was a significant correlation between each PHS and LTG (
r = 0.77,
r = 0.77,
P-value < 0.01). SD trait showed a significant correlation with LTG trait (
r = 0.70,
P-value < 0.01).
A total of 90 accessions (PHS-T, PHS-S, and 88 F
2:3) were sequenced using MGISEQ-2000 to identify polymorphic SNP position between PHS-T and PHS-S; 20,568 SNPs showing polymorphism were identified (
Supplementary Fig. S1). To construct a linkage map, these SNPs were filtered so that the distance between SNPs was 100 kb, and 567 SNPs were selected from total SNPs. 376 distinct positions of 567 SNPs were mapped on 17 linkage groups on 12 chromosomes. The linkage map was 991.8 cM in size with an average interval of 2.9 cM (
Fig. 5).
1 QTL was identified in the same SNP markers position under each condition. Under the field condition, 1 QTL (
qPHS1FC) was detected on chromosome 1 with LOD score of 35.8, thereby accounting for PVE of 89.0%. Under the chamber condition also, 1 QTL (
qPHS1GC) was detected at identical positions with LOD scores of 24.0, thereby accounting for PVE of 86.1% (
Table 2).
The QTLs of SD and LTG were detected in the F
2:3 populations, 1 QTLs of each trait was also identified in the same SNP markers position as PHS QTLs. In the SD trait, 1 QTL (
qSD1) was detected on chromosome 1 with LOD score of 8.6, thereby accounting for PVE of 35.5%. In the LTG trait, 1 QTL (
qLTG1) was detected at identical positions with LOD score of 13.7, thereby accounting for PVE of 49.6% (
Table 3).
Total 45 and 17 digenic epistatic QTLs for PHS were identified under the field condition and the chamber condition. Among those QTLs, 2 and 7 epistatic QTLs under field condition and chamber condition contained
qPHS1FC and
qPHS1GC, respectively. For SD and LTG, 3 and 4 epistatic interactions were detected, respectively. The 1 epistatic interaction of LTG contains
qLTG1, however
qSD1 was not involved in any epistatic interaction for SD (
Supplementary Table S2 and Fig. S2).
DISCUSSION
PHS is one of the essential traits for yield and quality of the grain, which is closely associated with seed dormancy, inhibits seed germination under prevailing high humidity and temperature conditions (
Bewley et al. 2013;
Rodríguez et al. 2015). Seed dormancy is a complex agronomic trait that is influenced not only environmental factors such as temperature and humidity but also endogenous hormones such as ABA, and GA (
Du et al. 2018). Therefore, to identify QTLs highly related to the genetic control of PHS tolerance, we performed evaluation under field and chamber conditions using the selected two inbred lines (PHS-T and PHS-S) in F
7 RIL and F
2:3 populations.
For the construction of the linkage map, after a series of filtering of 20,568 SNPs, a total of 567 SNPs was found suitable to build the genetic map. 567 SNPs were mapped to 376 distinct positions, but the patterns of classification revealed genetic diversity on chromosome 9. The presence of wide structural gaps was identified between the patterns (
Supplementary Fig. S1). In the previous study, regions of pattern with wide structural gaps on chromosome 9 were reported to have low levels of polymorphism in the Japonica group (
Santos et al. 2020).
The reported QTLs have been detected for PHS tolerance on chromosome 1.
qSD-1,
Sdr6,
qSD1, and
qDEG1 are located on the short arm of chromosome 1, and
qSD1.1 and
qSnd-1 are co-localized on the long arm of chromosome 1 (
Miura et al. 2002;
Gu et al. 2006;
Li et al. 2011;
Lu et al. 2011;
Marzougui et al. 2012;
Wang et al. 2014). The
qPHS1FC and
qPHS1GC were detected in this study at the marker interval ch01-0.040-ch01-0.053 (400 kb to 530 kb) on chromosome 1,
qSD1 and
qLTG1 region overlapped on the same position. The
Sdr6, the closest reported QTL compare to QTLs identified in this study, was detected in the interval between RM7278 and RM3425 (1,791 kb to 5,114 kb) on chromosome 1 using forty-four chromosome segment substitution lines (CSSLs) derived from a cross between Koshihikari and Nona Bokra (
Marzougui et al. 2012). The
PHS9 on chromosome 1 in rice regulated pre-harvesting sprouting by controlling the reactive oxygen species (ROS) signaling and ABA signaling through interaction with
OsGAP (
Xu et al. 2019). To clone the
PHS9, a mapping population was generated by crossing
phs9-D with Kasalath, and 53 bp deletions were detected as causal variation in
LOC_Os01g13950. However, there was no difference between PHS-T and PHS-S in this variation (data not shown). The region of QTLs detected in this study was not reported for PHS traits yet. In addition, all QTLs identified in this study showed large additive effect and PVE. This implies this QTLs are suitable to apply in marker assist breeding program.
PHS leads to a reduction in grain quality and grain yield in rice. Thus, developing a cultivar with PHS tolerance is one of the important goals in rice breeding. In this study, we used the F2 and F3 populations that were developed from selected PHS-T line and PHS-S line in the RILs to detect QTL of PHS-related phenotypes. We identified one novel major QTL for PHS tolerance under the two environments. It is hypothesized that the results of this study can be used as important resources for molecular breeding and to gain a better understanding of the genetics of the genetic traits in rice.
SUPPLEMENTARY MATERIALS
ACKNOWLEDGEMENTS
This work was supported for 2 years by Pusan National University.
Fig. 1Process of development of the plant materials for the current study.
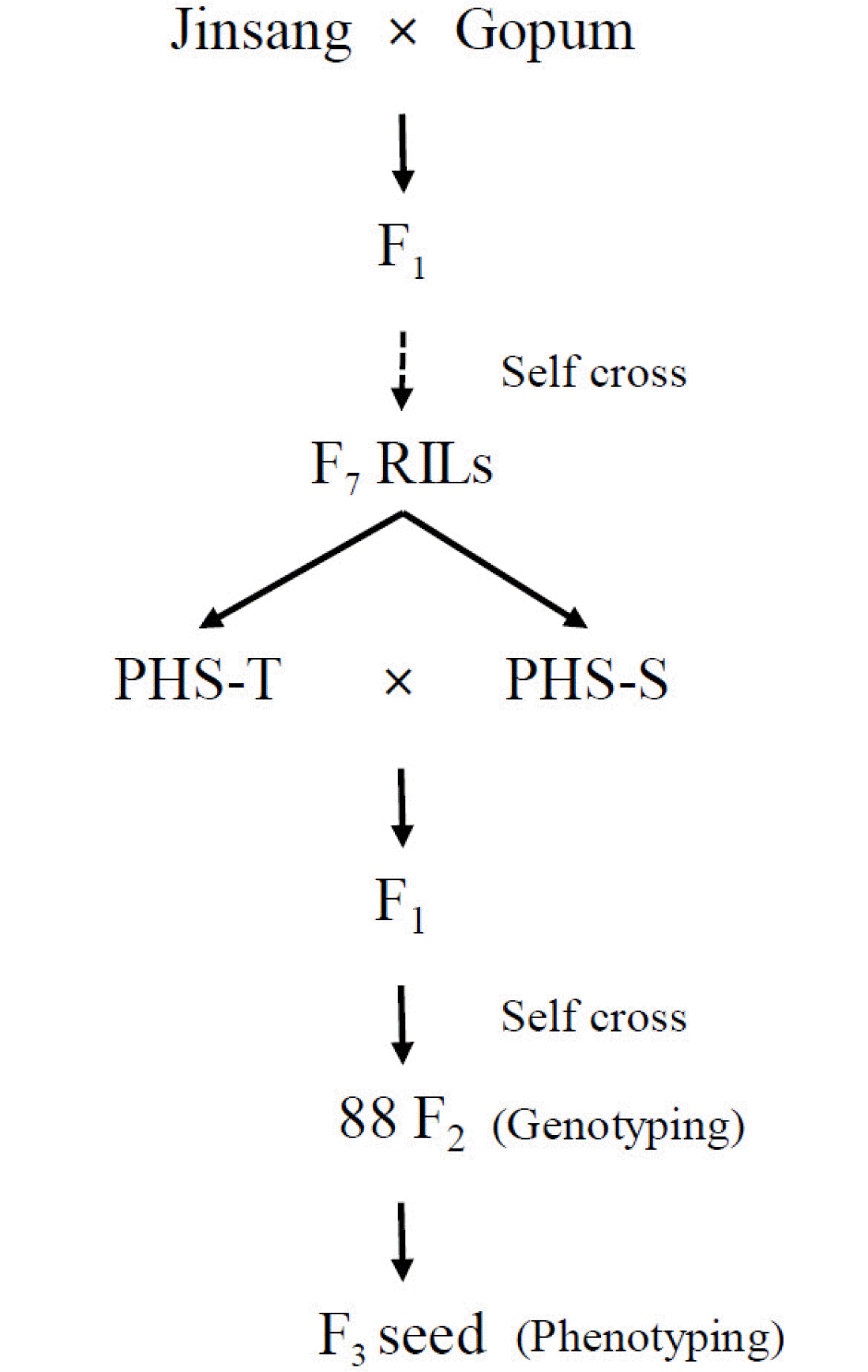
Fig. 2Experimental results of PHS-T and PHS-S under chamber and field conditions. (A) Pre-harvest sprouting under growth chamber condition. (B) Pre-harvest sprouting under field condition.

Fig. 3Frequency distributions of germination rate at 28℃ (A) and low-temperature germination rate (B) in parental cultivars (PHS-T, PHS-S) and F3 population.
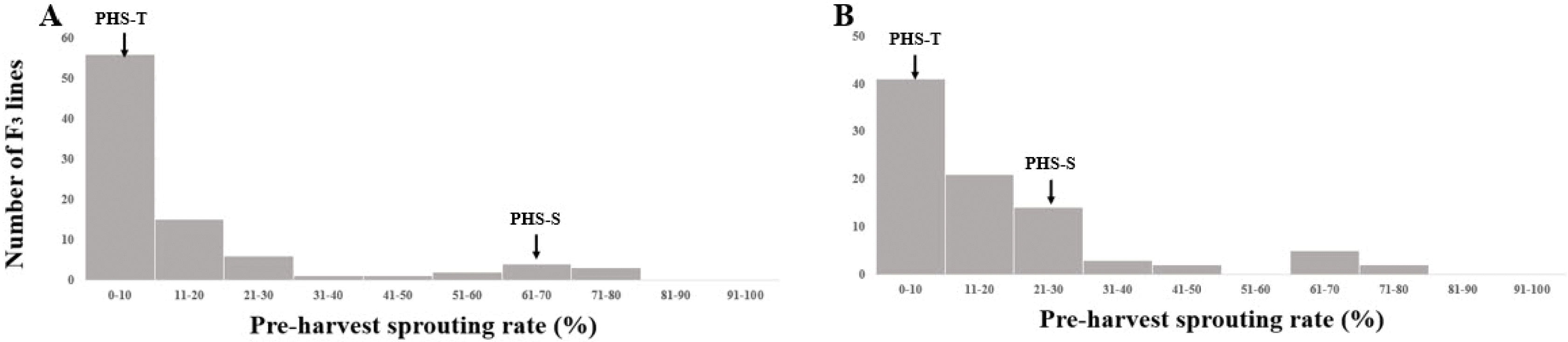
Fig. 4Frequency distributions of preharvest sprouting rates in parental cultivars (PHS-T, PHS-S), and F3 population under the growth chamber (A) and field conditions (B).
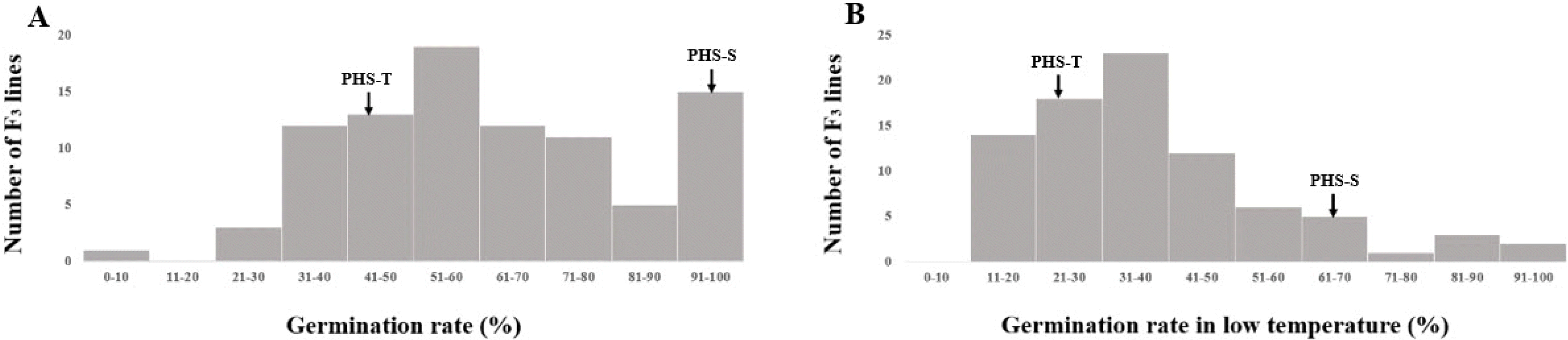
Fig. 5Linkage maps for preharvest sprouting tolerance, seed dormancy, and low-temperature germination in 88 F2 population. Markers are indicated on the right and genetic distances (cM).

Table 1Pearson’s correlation of preharvest sprouting under the field and growth chamber conditions, seed dormancy, and low-temperature germination using 88 F3 population.
Table 1
|
Trait |
Correlations |
|
|
PHS (Field)z)
|
PHS (Chamber) |
SD |
|
PHS (Chamber) |
0.82** |
|
|
|
SD |
0.64** |
0.57** |
|
|
LTG |
0.77** |
0.77** |
0.70** |
Table 2QTLs of preharvest sprouting identified using the F3 population under field and growth chamber conditions.
Table 2
|
Trait |
QTLs |
Linkage group |
Marker interval |
QTL peak location (cM) |
LODz)
|
PVEy) (%) |
Additive effectx)
|
|
PHS (Field) |
qPHS1FC
|
1 |
ch01-0.040/ch01-0.053 |
25.0 |
35.8 |
89.0 |
31.0 |
|
PHS (Chamber) |
qPHS1GC
|
1 |
ch01-0.040/ch01-0.053 |
25.0 |
24.0 |
86.1 |
28.4 |
Table 3QTLs of seed dormancy and low-temperature germination were identified using the F3 population under growth chamber condition.
Table 3
|
Trait |
QTLs |
Linkage group |
Marker interval |
QTL peak location (cM) |
LODz)
|
PVEy) (%) |
Additive effectx)
|
|
Seed dormancy |
qSD1
|
1 |
ch01-0.040/ch01-0.053 |
25.0 |
8.6 |
35.5 |
17.9 |
|
Low temperature germination |
qLTG1
|
1 |
ch01-0.040/ch01-0.053 |
25.0 |
13.7 |
49.6 |
20.6 |
References
- Almansouri M, Kinet JM, Lutts S. 2001. Effect of salt and osmotic stresses on germination in durum wheat (Triticum durum Desf.). Plant Soil. 231: 243-254.
- Bewley JD, Bradford KJ, Hilhorst HWM, Nonogaki H. 2013. Seeds: physiology of development, germination and dormancy. Springer. New York, U. S. A:
- Cai HW, Morishima H. 2000. Genomic regions affecting seed shattering and seed dormancy in rice. Theor. Appl. Genet.. 100: 840-846.
- Chen CX, Cai SB, Bai GH. 2008. A major QTL controlling seed dormancy and preharvest sprouting resistance on chromosome 4A in a Chinese wheat landrace. Mol. Breed.. 21: 351-358.
- Cheon KS, Won YJ, Jeong YM, Lee YY, Kang DY, Oh J, et al. 2020. QTL mapping for preharvest sprouting resistance in japonica rice varieties utilizing genome resequencing. Mol. Genet. Genom.. 295: 1129-1140.
- Chono M, Honda I, Shinoda S, Kushiro T, Kamiya Y, Nambara E, et al. 2006. Field studies on the regulation of abscisic acid content and germinability during grain development of barley: Molecular and chemical analysis of preharvest sprouting. J. Exp. Bot.. 57: 2421-2434.
- Cruz RPD, Milach SCK. 2004. Cold tolerance at the germination stage of rice: methods of evaluation and characterization of genotypes. Sci. Agric.. 61: 1-8.
- Debeaujon I, Koornneef M. 2000. Gibberellin requirement for Arabidopsis seed germination is determined both by testa characteristics and embryonic abscisic acid. Plant Physiol.. 122: 415-424.
- Du L, Xu F, Fang J, Gao SP, Tang JY, Fang S, et al. 2018. Endosperm sugar accumulation caused by mutation of PHS8/ISA1 leads to preharvest sprouting in rice. Plant J.. 95: 545-556.
- Fang J, Chai C, Qian Q, Li C, Tang J, Sun L. 2008. Mutations of genes in synthesis of the carotenoid precursors of ABA lead to preharvest sprouting and photo-oxidation in rice. Plant J.. 54: 177-189.
- Gao FY, Ren GJ, Lu XJ, Sun SX, Li HJ, Gao YM, et al. 2008. QTL analysis for resistance to preharvest sprouting in rice (Oryza sativa). Plant Breed.. 127: 268-273.
- Graeber K, Nakabayashi K, Miatton E, Leubner-Metzger G, Soppe WJ. 2012. Molecular mechanisms of seed dormancy. Plant Cell Environ.. 35: 1769-1786.
- Gu XY, Chen ZX, Foley ME. 2003. Inheritance of seed dormancy in weedy rice. Crop Sci.. 43: 835-843.
- Gu XY, Kianian SF, Foley ME. 2004. Multiple loci and epistases control genetic variation for seed dormancy in weedy rice (Oryza sativa). Genetics. 166: 1503-1516.
- Gu XY, Kianian SF, Foley ME. 2006. Isolation of three dormancy QTLs as Mendelian factors in rice. Heredity. 96: 93-99.
- Gu XY, Foley ME, Horvath DP, Anderson JV, Feng J, Zhang L, et al. 2011. Association between seed dormancy and pericarp color is controlled by a pleiotropic gene that regulates abscisic acid and flavonoid synthesis in weedy red rice. Genetics. 189: 1515-1524.
- Gubler F, Millar AA, Jacobsen JV. 2005. Dormancy release, ABA and preharvest sprouting. Curr. Opin. Plant Biol.. 8: 183-187.
- Hong WJ, Yoon MY, Ra WH, Park JW, Min SK, Kwon SW, et al. 2014. A new variety 'Jinsang' with lodging tolerant and low amylose content. Korean J. Breed. Sci.. 46: 103-107.
- Hori K, Sugimoto K, Nonoue Y, Ono N, Matsubara K, Yamanouchi U, et al. 2010. Detection of quantitative trait loci controlling preharvest sprouting resistance by using backcrossed populations of japonica rice cultivars. Theor. Appl. Genet.. 120: 1547-1557.
- Hyun DY, Oh M, Choi YM, Lee S, Lee MC, Oh S. 2017. Morphological and molecular evaluation for germinability in rice varieties under low-temperature and anaerobic conditions. J. Crop Sci. Biotechnol.. 20: 21-27.
- Kang S, Shon J, Kim HS, Kim SJ, Choi JS, Park JH, et al. 2018. Analysis of genetic variation in preharvest sprouting at different cumulative temperatures after heading of Rice. Korean J. Crop Sci.. 63: 8-17.
- Kosambi DD. 1944. Caste and class in India. Sci. Soc.. 8: 243-249.
- Lee O, Park Y, Kim ES, Kim S. 2016. Projection of Korean probable maximum precipitation under future climate change scenarios. Adv. Meteorol.. 2016: 1-16.
- Li B, Foley ME. 1997. Genetic and molecular control of seed dormancy. Trends Plant Sci.. 2: 384-389.
- Li W, Xu L, Bai X, Xing Y. 2011. Quantitative trait loci for seed dormancy in rice. Euphytica. 178: 427-435.
- Lin SY, Sasaki T, Yano M. 1998. Mapping quantitative trait loci controlling seed dormancy and heading date in rice, Oryza sativa L., using backcross inbred lines. Theor. Appl. Genet.. 96: 997-1003.
- Lu B, Xie K, Yang C, Wang S, Liu X, Zhang L. 2011. Mapping two major effect grain dormancy QTL in rice. Mol. Breed.. 28: 453-462.
- Marzougui S, Sugimoto K, Yamanouchi U, Shimono M, Hoshino T, Hori K. 2012. Mapping and characterization of seed dormancy QTLs using chromosome segment substitution lines in rice. Theor. Appl. Genet.. 124: 893-902.
- Meng L, Li H, Zhang L, Wang J. 2015. QTL IciMapping: integrated software for genetic linkage map construction and quantitative trait locus mapping in biparental populations. Crop J.. 3: 269-283.
- Miura K, Lin S, Yano M, Nagamine T. 2002. Mapping quantitative trait loci controlling seed longevity in rice (Oryza sativa L.). Theor. Appl. Genet.. 104: 981-986.
- Mizuno Y, Yamanouchi U, Hoshino T, Nonoue Y, Nagata K, Fukuoka S, et al. 2018. Genetic dissection of preharvest sprouting resistance in an upland rice cultivar. Breed. Sci.. 68: 200-209.
- Porebski S, Bailey LG, Baum BR. 1997. Modification of a CTAB DNA extraction protocol for plants containing high polysaccharide and polyphenol components. Plant Mol. Biol. Rep.. 15: 8-15.
- Rodríguez M, Barrero JM, Corbineau F, Gubler F, Benech-Arnold RL. 2015. Dormancy in cereals (not too much, not so little): about the mechanisms behind this trait. Seed Sci. Res.. 25: 99-119.
- Santos JG, Billot C, Chebotarov D, Droc G, Lorieux M, McNally K, et al. 2020. Multiple streams of genetic diversity in Japonica rice. BioRxiv.
- Sugimoto K, Takeuchi Y, Ebana K, Miyao A, Hirochika H, Hara N, et al. 2010. Molecular cloning of Sdr4, a regulator involved in seed dormancy and domestication of rice. Proc. Natl. Acad. Sci. U.S.A.. 107: 5792-5797.
- Tejakhod S, Ellis RH. 2018. Effect of simulated flooding during rice seed development and maturation on subsequent seed quality. Seed Sci. Res.. 28: 72-81.
- Wan JM, Jiang L, Tang JY, Wang CM, Hou MY, Jing W, et al. 2006. Genetic dissection of the seed dormancy trait in cultivated rice (Oryza sativa L.). Plant Sci.. 170: 786-792.
- Wang L, Cheng J, Lai Y, Du W, Huang X, Wang Z, et al. 2014. Identification of QTLs with additive, epistatic and QTL × development interaction effects for seed dormancy in rice. Planta. 239: 411-420.
- Xu F, Tang J, Gao S, Cheng X, Du L, Chu C. 2019. Control of rice preharvest sprouting by glutaredoxin-mediated abscisic acid signaling. Plant J.. 100: 1036-1051.
- Ye H, Feng J, Zhang L, Zhang J, Mispan MS, Cao Z, et al. 2015. Map-based cloning of seed dormancy1-2 identified a gibberellin synthesis gene regulating the development of endosperm-imposed dormancy in rice. Plant Physiol.. 169: 2152-2165.
- Zhang Y, Miao X, Xia X, He Z. 2014. Cloning of seed dormancy genes (TaSdr) associated with tolerance to preharvest sprouting in common wheat and development of a functional marker. Theor. Appl. Genet.. 127: 855-866.