Abstract
The anaerobic germination 1 (AG1) locus, which confers tolerance to the anaerobic germination of rice seed, has been previously identified and a gel-based InDel DNA marker developed for marker-assisted selection (MAS). However, there is a need for marker development for high-throughput genotyping in order to increase the breeding efficiency. Single nucleotide polymorphisms (SNPs)-based Kompetitive Allele Specific PCR (KASP) markers enable high-accuracy and high-throughput analyses for the genotyping of AG1 locus. In the present study, we developed one AG1 InDel-specific and four flanking KASP markers, which can be used as the foreground and recombination markers, respectively, for the AG1 locus. Of those, three KASP markers developed were validated with Dasan-AG1 (BC3F2) segregating lines; the AG1 InDel-specific KASP marker was 100% consistent with the existing AG1 gel-based marker, while the two flanking markers showed 70.3 and 66.7 percent consistency to the AG1 gel-based marker, respectively. In addition, the five KASP markers developed were further used to genotype 78 Korean and 95 foreign rice varieties. The genotyping results with 172 varieties revealed that most of the Korean varieties possessed the AG1 locus; however, a wide variation was observed in the foreign varieties for AG1 locus. Hence, the KASP markers developed for AG1 locus show promise as useful tools for expediting the breeding of varieties tolerant to anaerobic germination stress via high-throughput genotyping.
-
Key words: Anaerobic germination, AG1, Abiotic tolerance, Gel-based marker, KASP marker, InDel, Single-nucleotide polymorphism
Direct seeding of rice (
Oryza sativa L.) is gaining popularity among farmers owing to its low cost and reduced labor intensity (
Pandey et al. 2002). However, the rice seeds in the direct seeding fields are submerged and suffer from hypoxia or anoxia, leading to poor seedling establishment. Rice is extremely sensitive to anoxia during germination and seedling growth, resulting in anaerobic germination (AG) being the main limiting factor for the wide adoption of direct seeding methods (
Hsu and Tung 2015). Hence, the breeding of AG tolerant rice varieties could be an effective method for resolving the limitation of the direct seeding of rice (
Yang et al. 2019).
The gene loci for tolerance to anaerobic germination have been identified mainly from indica or weed rice. The quantitative trait locus (QTL)
qAG-9-2 on chromosome 9 (
Angaji et al. 2010) was derived from Khao Hlan On (KHO), a weed rice plant in Myanmar, and accounted for 33.5% of the phenotypic variation related to anaerobic germination tolerance (
Angaji et al. 2010). The other major QTLs for AG tolerance were identified on chromosome 7:
qAG7.1, derived from the native Ma-Zhan Red variety in China and accounting for 31.7% of the phenotypic variation (
Septiningsih et al. 2013), and
qAG7 which is derived from Nanhi, belonging to the aus-type rice, and accounts for 22.3% of the phenotypic variation (
Baltazar et al. 2014). Some minor QTLs for AG tolerance showing relatively low phenotypic variation were also identified;
qAG1,
qAG2, and
qAG7 on chromosomes 1, 2 and 7 were identified from the Japanese variety Kinmaze (
Jiang et al. 2004), and
qAG-5 and
qAG-11 on chromosomes 5 and 11 were identified from USSR5 (
Jiang et al. 2006). A recent study has identified a novel QTL
qAG2.1 on chromosome 2 in the japonica variety Gihobyeo, accounting for 9.19% of the phenotypic variation (
Kim et al. 2019).
Of the numerous AG tolerance loci that have been identified, only
qAG-9-2 QTL has been fine-mapped and cloned as
OsTPP7, which encodes a trehalose-6-phosphate phosphatase (TPP) (
Kretzschmar et al. 2015).
OsTPP7 (LOC_Os09g20390) is located in the 21 kb InDel region, which was identified by comparing IR64 and KHO and by fine-mapping for
qAG-9-2 in about 50 kb (
Kretzschmar et al. 2015). The functional analysis suggested that OsTPP7 is involved in enhancing starch mobilization to promote embryo germination and coleoptile elongation. Rice has 13 TPP-like genes, some of which have been reported to turn trehalose-6-phosphate (T6P) to trehalose (
Shima et al. 2007;
Ge et al. 2008). Therefore, OsTPP7 activates the anaerobic germination tolerance by modulating local T6P/sucrose ratios (
Lunn et al. 2014;
Yadav et al. 2014;
Kretzschmar et al. 2015).
DNA markers closely linked to target gene/QTL are a useful tool for transferring the target loci into elite breeding lines, and the process is referred to as marker-assisted selection (MAS). This approach is favored when multiple QTLs control the phenotypic variability of one trait under selection. A gel-based DNA marker for anaerobic germination was developed based on the 21-kb InDel region of
qAG-9-2 (
Kretzschmar et al. 2015). However, a gel-based marker is low-throughput, time-consuming and labor intensive as it often requires multiple rounds of PCR and gel electrophoresis (
Ramkumar et al. 2015). KASP is one of the uniplex SNP genotyping platforms, being extensively used in massive SNP genotyping studies and markers-assisted breeding due to its low cost, high throughput, and high accuracy (
Semagn et al. 2014). KASP genotyping can be performed in 96-, 384-, and 1,536-well plates, enabling a much higher throughput compared to the gel-based method.
Hence, in this study, we developed a KASP marker targeting the InDel region of AG1 locus located in qAG-9-2 QTL. We also developed additional KASP markers targeting SNPs located in the flanking region of AG1 locus; these flanking markers can be used as recombination markers which enable efficient selection of segregating lines with recombination events in the flanking regions. Further profiling of domestic and foreign rice varieties was also performed with the KASP markers developed in the study.
MATERIALS AND METHODS
Plant materials
Rice varieties tolerant (KHO, IR64-AG1) and susceptible (IR64, IR64-Sub1A) to anaerobic germination stress were used to develop the AG1-specific KASP markers. IR64-AG1 (KHO allele) and IR64-Sub1A (FR13A allele), which harbor AG1 and Sub1 loci, respectively, in IR64 background were previously developed and imported from the IRRI through a material transfer agreement. Twenty-seven BC3F2 lines used for the validation of the three KASP markers were developed from a cross between the IR64-AG1 and Dasan varieties, in which Dasan was used as a recurrent parent.
Analysis of gel-based markers for AG1
The primers for the gel-based
AG1 InDel marker (DFR marker with three primers; DFR_F2, DFR_R2 and DFR_LB2) were previously designed based on the 21-kb InDel region-specific markers for
qAG-9-2 QTL (
Kretzschmar et al. 2015). The total genomic DNA was extracted from leaf samples according to the cetyltrimethyl ammonium bromide (CTAB) method (Murray and Thomson 1980). The PCR amplification of the gel-based
AG1 InDel marker was performed by initial denaturation at 96℃ for 4 minutes, followed by 35 cycles of denaturation at 94℃ for 30 seconds, annealing at 55℃ for 30 seconds, and extension at 72℃ for 1 minute, followed by a final extension at 72℃ for 4 minutes. The PCR products were separated by electrophoresis on 2% agarose gel and visualized using the gel-imager (Davinch-Gel
TMImaging System).
KASP marker assays for SNPs/InDels were performed based on the manual of LGC Genomics (
http://www.lgcgenomics.com). The primers for the developed markers have been listed in
Table 1. KASP markers assay was performed in a 96-well format and set up as 10 mL reactions with 4.86 mL of DNA template, 0.14 mL of assay mix (containing a common primer and two or three different allele specific primers), and 5 mL of master mix (containing fluorescence resonant energy transfer and Taq polymerase). Amplification was started at 94℃ for 15 minutes, followed by 10 cycles at 94℃ for 20 seconds and at 61℃ for 60 seconds (touchdown to 55℃, ‒1℃ per cycle), followed by 26 cycles at 94℃ for 20 seconds, 55℃ for 60 seconds, and a read step at 37℃ for 1 minute. After amplification, the fluorescence signals from the end PCR products were read with the CFX Maestro software (Bio-Rad, U.S.A.) using the recommended excitation and emission values.
RESULTS
Development of InDel/SNP-specific KASP markers for AG1 locus
AG1 locus, representing the 21-kb InDel region located in
qAG9-2 QTL (
Angaji et al. 2010), is one of the major genomic loci determining anaerobic germination tolerance in rice. The development of a gel-based marker targeting the InDel region of
AG1 has been previously reported (
Kretzschmar et al. 2015;
Fig. 1A). Since the KASP marker system enables accurate and high-throughput genotyping and thereby accelerates the efficiency of MAS, we developed KASP markers targeting this
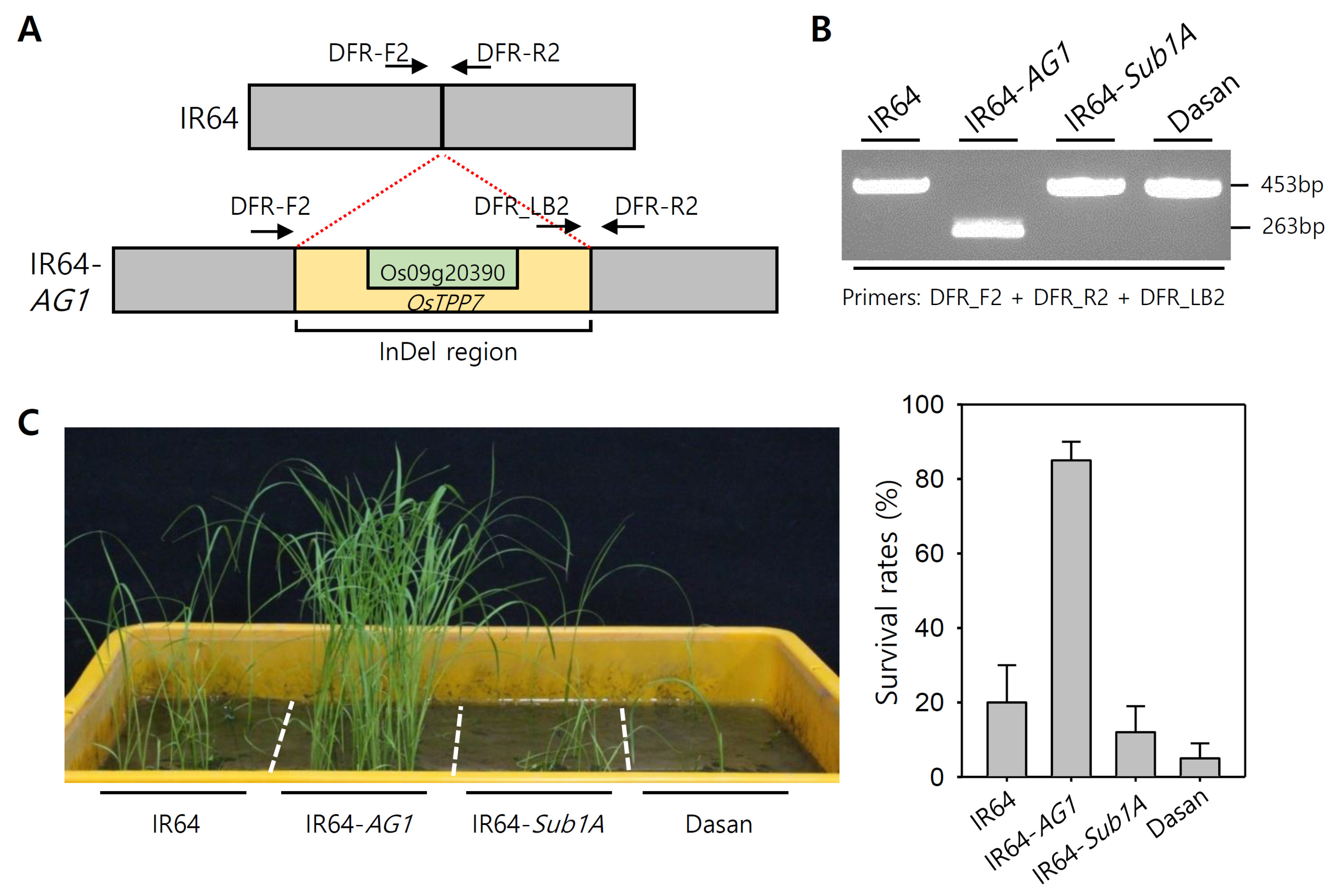
AG1 locus as a replacement for the gel-based marker. Using the gel-based markers, we first genotyped four control varieties including IR64-
AG1, a backcross introgression line derived from a cross between IR64 and KHO. IR64-
Sub1A, an introgressed line harboring
Sub1A in the IR64 background genotype, was also used as a control variety for phenotyping and genotyping. This was done since the AG and submergence tolerance traits are quite similar in terms of tolerance for excessive water stress, but different in their tolerance mechanisms. The genotyping of the control varieties by the existing gel-based markers showed that only IR64-
AG1 harbored
AG1 insertion fragment, but deletion band patterns were observed in the other three control varieties (
Fig. 1B). PCR amplification using the set of three primers (
AG1 gel-based marker) produced 263 bp and 453 bp bands for
AG1 insertion and deletion, respectively. This method failed with DFR-F2 and DFR-R2 primers for
AG1 insertion, since the expected PCR product would be more than 20 kb in size; hence PCR amplification occurred only between DFR-LB2 and DFR-R2. The phenotypic screening of the control varieties under submergence conditions during the period of seedling establishment revealed that only the IR64-
AG1 variety showed a tolerant phenotype; the other varieties had a survival rate of less than 20% (
Fig. 1D).
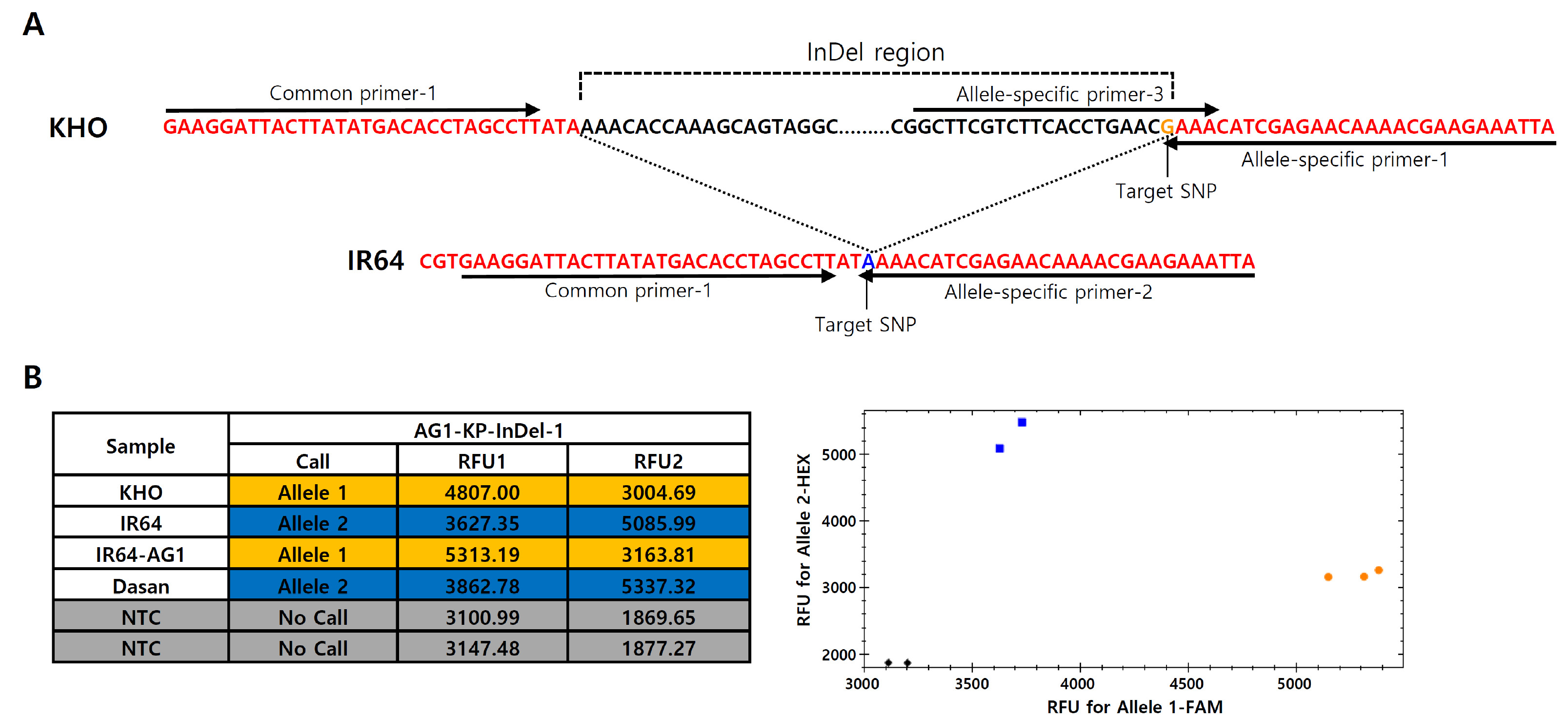
In order to develop KASP markers for the
AG1 InDel region, amplicon sequencing was performed by PCR for amplifying the target region with the gel-based DFR marker (primers; DFR_F2, DFR_R2 and DFR_LB2). The nucleotide sequences obtained from IR64 and KHO were aligned using NCBI blast (BLAST: Basic Local Alignment Search Tool) to observe sequence variation between the two genotypes (
Fig. 2A). Based on the sequence variation, allele specific KASP primers targeting SNP (G for KHO and A for IR64 alleles) were designed at the ends of the InDel region. For the development of codominant markers distinguishing the SNP (G/A), allele-specific primer-3 hybridizing on the InDel fragment was also designed, constituting four primers in one set named
AG1-KP-InDel-1 (
Fig. 2A). The
AG1-KP-InDel-1 marker thus obtained was validated by genotyping of the control varieties. In the KASP marker analysis, the scatter plot of the two fluorescence signals were clearly separated into tolerant and sensitive alleles; KHO and IR64-
AG1 alleles were shown as tolerant alleles and the other varieties were shown as susceptible genotypes (
Fig. 2B).
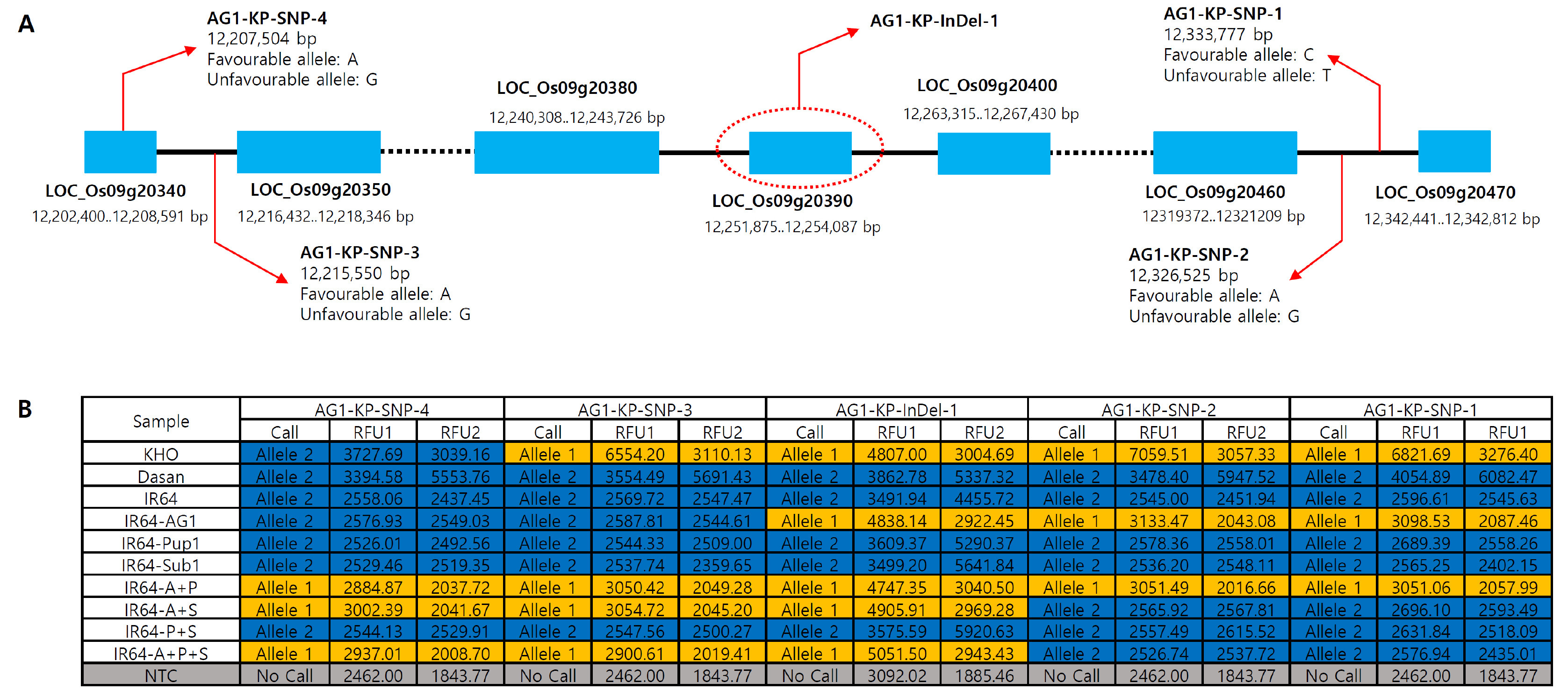
In addition to the development of the foreground KASP marker targeting the
AG1 InDel locus, we also developed flanking markers that can be used to select the progenies with recombination events around the
AG1 locus in the segregating population. Candidate SNP information was obtained from the previous SNP database analyzed by the comparison of whole genome sequences of KHO and IR64 (
Platten et al. 2019). A total of 4 SNP positions on both sides of
AG1 locus located within a physical distance of about 140 kb (12,202,400-12,342,441 bp) were chosen and primers were designed. Of the four flanking KASP markers, two markers (
AG1-KP-SNP-1 and
AG1-KP-SNP-2) were successfully validated with the control varieties, showing KHO allele in IR64-
AG1 and IR64 allele in IR64 and Dasan. However, KASP assay with the other two markers (
AG1-KP-SNP-3 and
AG1-KP-SNP-4) showed only IR64 allele in both IR64-
AG1 and IR64 (
Fig. 3B). Thus, we tested more breeding lines including multi-QTLs pyramiding lines. Notably, the two flanking markers showed KHO allele in IR64-
AG1-Pup1, IR64-
AG1-Sub1 and IR64-
AG1-Pup1-Sub1 lines which harbor
AG1 locus, indicating that the genomic segment size of
AG1 locus are various among
AG1-harboring breeding lines (
Fig. 3B). Conclusively, the four KASP markers were successfully validated with the control varieties and multiple breeding lines (
Fig. 3B).
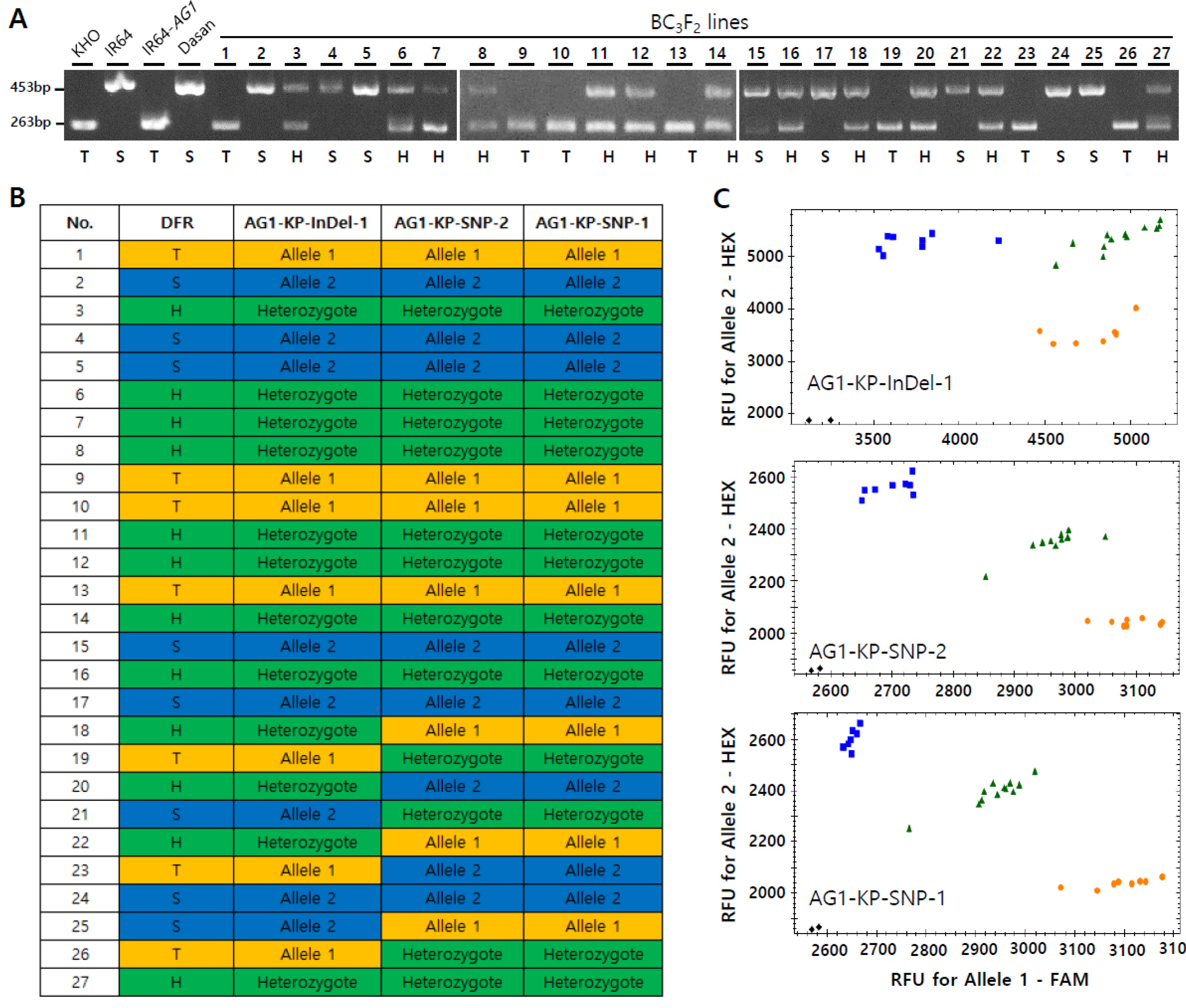
To validate the KASP markers developed in the current study, we performed genotyping of the segregating BC
3F
2 lines obtained from the cross between the IR64-
AG1 and Dasan varieties, wherein Dasan was used as a recurrent parent. In order to analyze the consistency of the markers, 27 BC
3F
2 lines and the control panel varieties were genotyped with the gel-based DFR marker as well as the three KASP markers (
AG1-KP-InDel-1,
AG1-KP-SNP-1 and
AG1-KP-SNP-2) (
Fig. 4); the other two flanking markers (
AG1-KP-SNP-3 and
AG1-KP-SNP-4) were not used for genotyping of the BC
3F
2 lines because they were not polymorphic between IR64-
AG1 and IR64 (
Fig. 3B). The genotyping result showed that allelic patterns were well segregated with all three markers in the segregating lines (
Fig. 4). The percentage of consistency in the allele types genotyped by the DFR marker was 100, 70.3 and 66.7 with those of
AG1-KP-InDel-1,
AG1-KP-SNP-1 and
AG1-KP-SNP-2, respectively (
Table 2). The Cohen’s kappa coefficient, which is a statistical measure of inter-rater reliability or agreement (
Landis and Koch 1977), was calculated for the analysis of statistical agreement among the markers. The agreement of the allele types by the DFR marker with
AG1-KP-InDel-1,
AG1-KP-SNP-1 and
AG1-KP-SNP-2, was 1.0, 0.54 and 0.48, respectively, indicating ‘Very good’ with the InDel marker and ‘Moderate’ with the two flanking markers (
Table 2). The result demonstrates that the two flanking markers showing ‘moderate agreement’ with the gel-based marker can be used as recombinant markers in marker-assisted backcross.
For further validation of all five KASP markers, we utilized them to profile 172 Korean/foreign varieties for
AG1 locus. All the varieties, except for two, were categorized into one of the homozygous types (tolerance or susceptible) (
Table 3). The profile results of 172 domestic and foreign varieties by KASP assays revealed that, except for four varieties (Milyang23, Dasan, Sobi, Mokyang), most of the Korean varieties tested harbored the KHO type allele for
AG1 flanking locus, while the foreign variety population showed a large variation (
Table 3). Interestingly, all of the Korean varieties profiled in our study showed the same allele types (tolerant or susceptible) for the four KASP markers (
AG1-KP-SNP-3,
AG1-KP-InDel-1,
AG1-KP-SNP-2 and
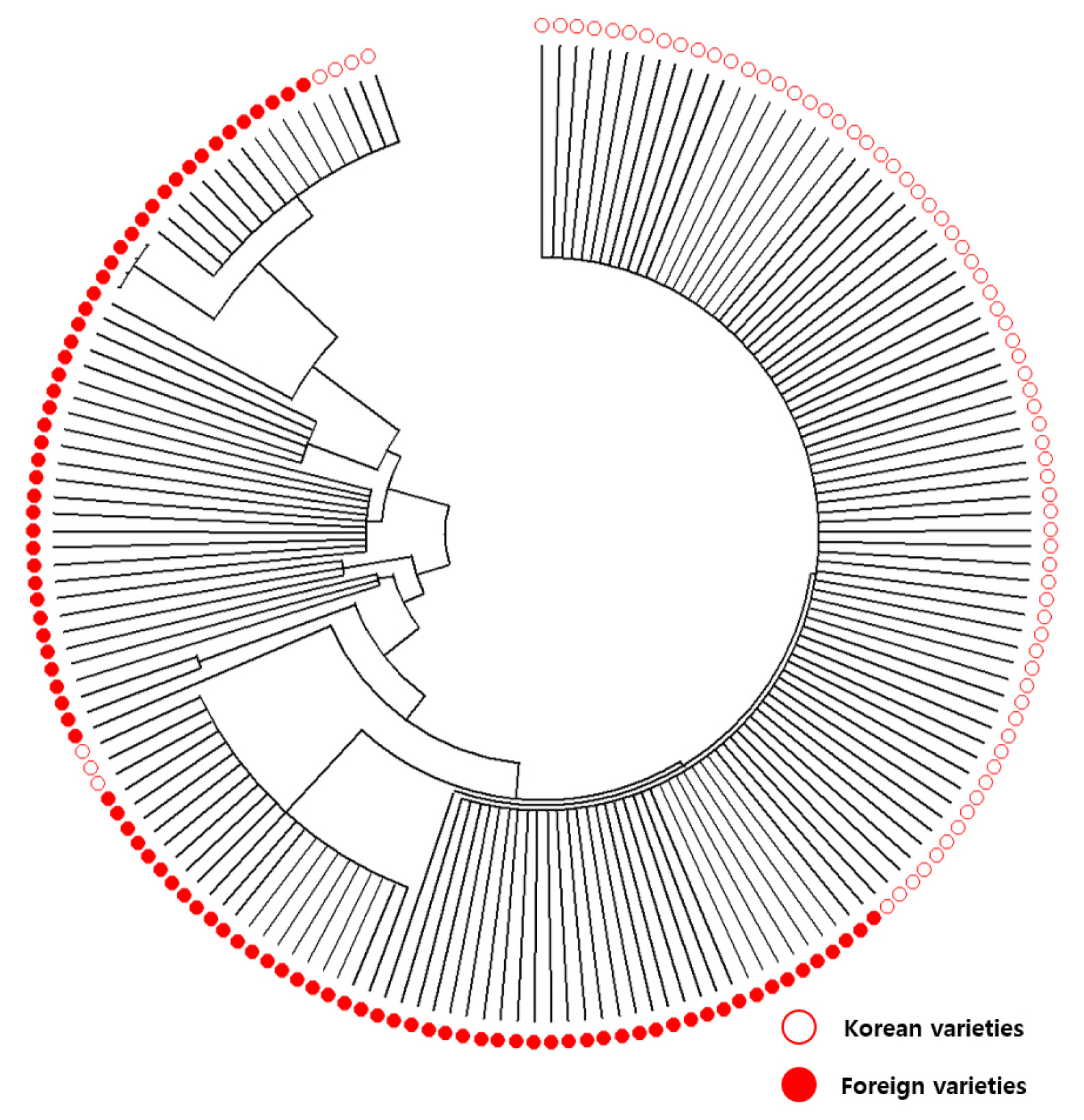
AG1-KP-SNP-1) except four varieties, while foreign varieties possessed various types of alleles, susceptible and tolerant, for the five markers. Phylogenetic analysis with Neighbor joining tree for the profiling of the 172 rice varieties revealed that Korean and foreign varieties clearly separated in
AG1 genotype (
Fig. 5), indicating the genetic constitution of Korean and foreign varieties are different on
AG1 locus.
DISCUSSION
AG is a useful trait, particularly in direct-seeded rice fields. The development of molecular DNA markers would increase the efficiency of AG trait introgression to the breeding lines. However, compared to the SNP-based KASP marker, the traditional DNA markers such as STS and SSR markers are low throughput, time consuming and not user-friendly. Hence, we developed KASP markers for the AG1 locus and the flanking region of AG1, which could be used for foreground and background selection, respectively. The KASP markers thus developed were validated by genotyping the segregating breeding lines and further used to perform profiling of domestic and foreign rice varieties for AG1 locus in the KASP assay.
The replacement of existing gel-based markers with SNP-based KASP markers would be a good strategy for increasing the selection efficiency by high-throughput genotyping.
Zhao et al. (2016) developed KASP markers for
AhFAD2 gene, which regulated the desaturase activity of fatty acids, replacing the existing CAPS marker.
Tan et al. (2017) also developed KASP markers for the greenbug resistance gene
Gb7 and the Hessian fly resistance gene
H32 in wheat by replacing the existing gel-based markers; these KASP markers were validated with various populations. In our previous study, we had developed the gene-specific KASP markers for submergence tolerance
Sub1 by replacing the existing gel-based markers, AEX1 and GnS2 (
Moon et al. 2019). It should be noted that the replaced KASP markers must be accurate and consistent with the genotyping result of the existing markers, if developed based on the same SNPs. As expected, the KASP marker (
AG1-KP-InDel-1) developed in this study showed 100% consistency with the existing gel-based marker. Thus, this KASP marker could be used as a foreground marker targeting
AG1 locus for high-throughput analysis in the breeding process.
In addition to the foreground markers which can be used to select genomic segments containing the target gene/QTLs (foreground selection), the linked flanking markers could also be used for the selection of segregating progenies with the target loci and recombination events between the target locus and linked flanking markers (recombinant selection). In plant breeding, backcrossing is used to introgress favorable traits from a donor plant into an elite variety (recurrent parent). Backcrossing is repeated with the recurrent parent until most of the genomic segments derived from the donor are eliminated. However, the donor segments that might harbor unfavorable traits can remain in the vicinity of the target locus, even after many backcrossing generations. Recombinant marker assays can help minimize this linkage drag during the selection process (
Frisch et al. 1999).
Tanksley et al. (1989) demonstrated that the reconstruction of the recurrent parent genotype usually requires more than six generations, while only three generations are required in the marker-assisted backcross. In order to develop recombinant markers of
AG1 locus, we designed the primers for KASP markers based on the SNPs located at a physical distance of about 70-90 kb from the
AG1 locus. The marker validation analysis with 27 BC
3F
2 lines revealed that the allele types genotyped by
AG1-KP-InDel-1 marker showed 70.3 and 66.7% consistency with those of
AG1-KP-SNP-1 and
AG1-KP-SNP-2, respectively (
Table 2). This finding points to the occurrence of active recombination between these markers. The other two flanking markers (
AG1-KP-SNP-3 and
AG1-KP-SNP-4) also showed the occurrence of recombination from
AG1-KP-InDel position (
Table 3). Thus, the InDel KASP marker and the other four flanking KASP markers would serve as useful tools for high-throughput selection as replacement and recombination markers, respectively, during the breeding of
AG tolerant rice varieties.
The five KASP markers developed in this study were used for the profiling of 172 rice varieties for AG1 locus. The profiling analysis revealed that, with the exception of four varieties (Milyang23, Dasan, Sobi, Mokyang), all the 73 Korean varieties possessed tolerant genotype harboring KHO allele for AG1 locus, while 95 foreign varieties showed a large variation in the AG1 allele type. Since most of the Korean varieties are japonica subspecies, this result shows that the tolerant allele type is broadly immanent in japonica varieties. Further investigation is needed not only for the AG1 InDel, but also for the effect of gene-specific allele types within the InDel region on AG tolerance.
ACKNOWLEDGEMENTS
This work was supported by the Next-Generation BioGreen 21 Program grant (Plant Molecular Breeding Center, No. PJ01319601), Rural Development Administration, Republic of Korea.
Fig. 1Genotyping and phenotyping of IR64-
AG1 plants with control plants. (A) Schematic diagram of InDel region in
AG1 locus. The yellow color represents the InDel region located in
AG1 locus and the light green color represents the region of
OsTPP7 (Os09g20390). The arrows indicate the primer positions of the existing gel-based marker (DFR marker,
Kretzschmar et al. 2015). (B) PCR amplification of control varieties with existing
AG1 gel-based marker. (C) Phenotypic screening and survival rates of the control plants for anaerobic germination.
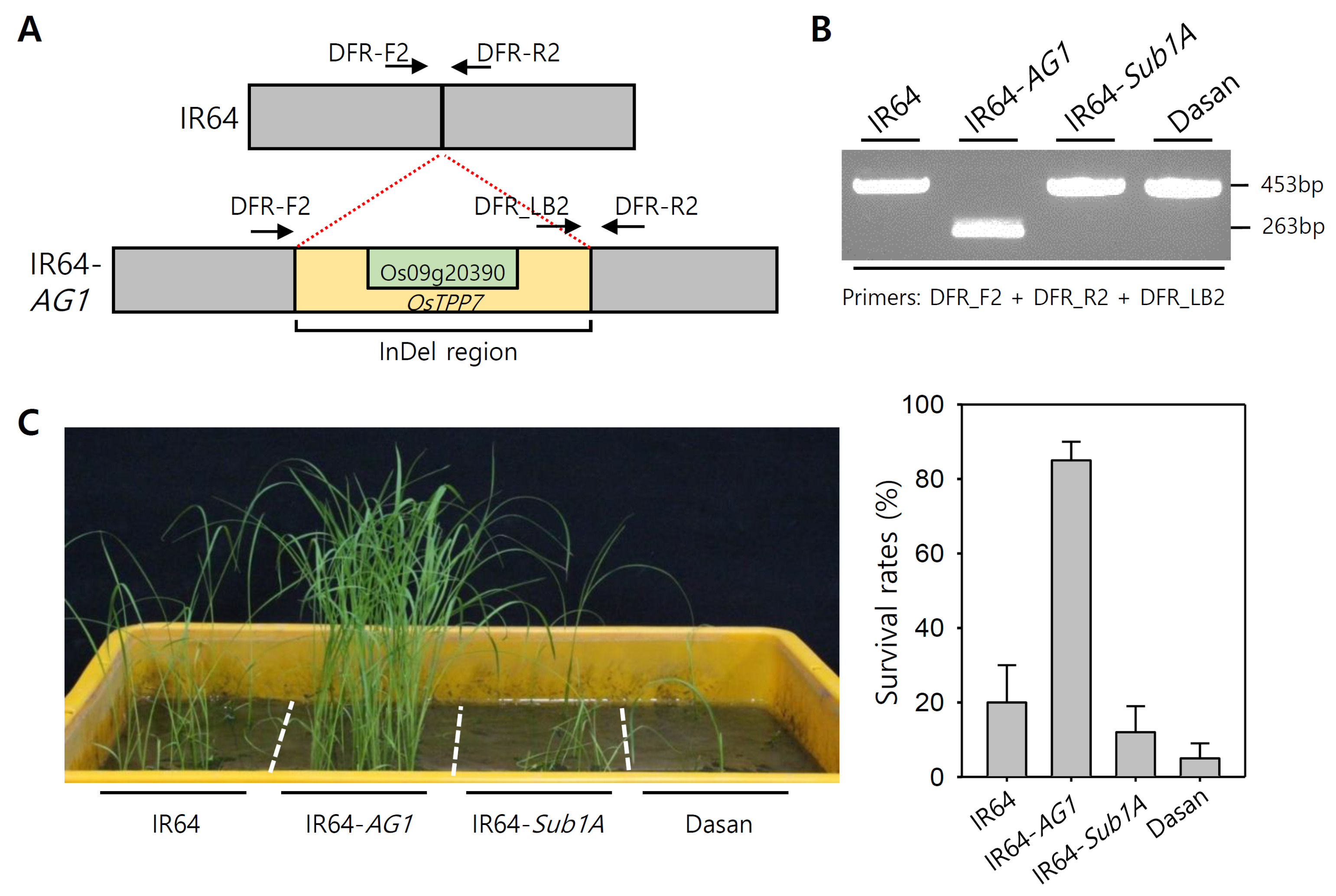
Fig. 2Primer design of AG1-InDel region for KASP marker development and genotyping of the control varieties. (A) Primer design of AG1-KP-InDel-1 marker in AG1-InDel region. Arrows indicate the location and direction of the primer sequences. One common primer (Common-primer-1) and three allele-specific primers (Allele-specific primer-1, Allele-specific primer-2 and Allele-specific primer-3) targeting the SNP position (G/A) were designed. (B) Genotyping results of the control varieties by the developed AG1-InDel specific KASP marker (left table) and the scatter plot of the KHO and IR64 type alleles (right panel). Allele 1 and Allele 2 are tolerance (KHO) and susceptible (IR64) types, respectively. RFU1 and RFU2 represent FAM and HEX fluorescent values, respectively (NTC = non template control).
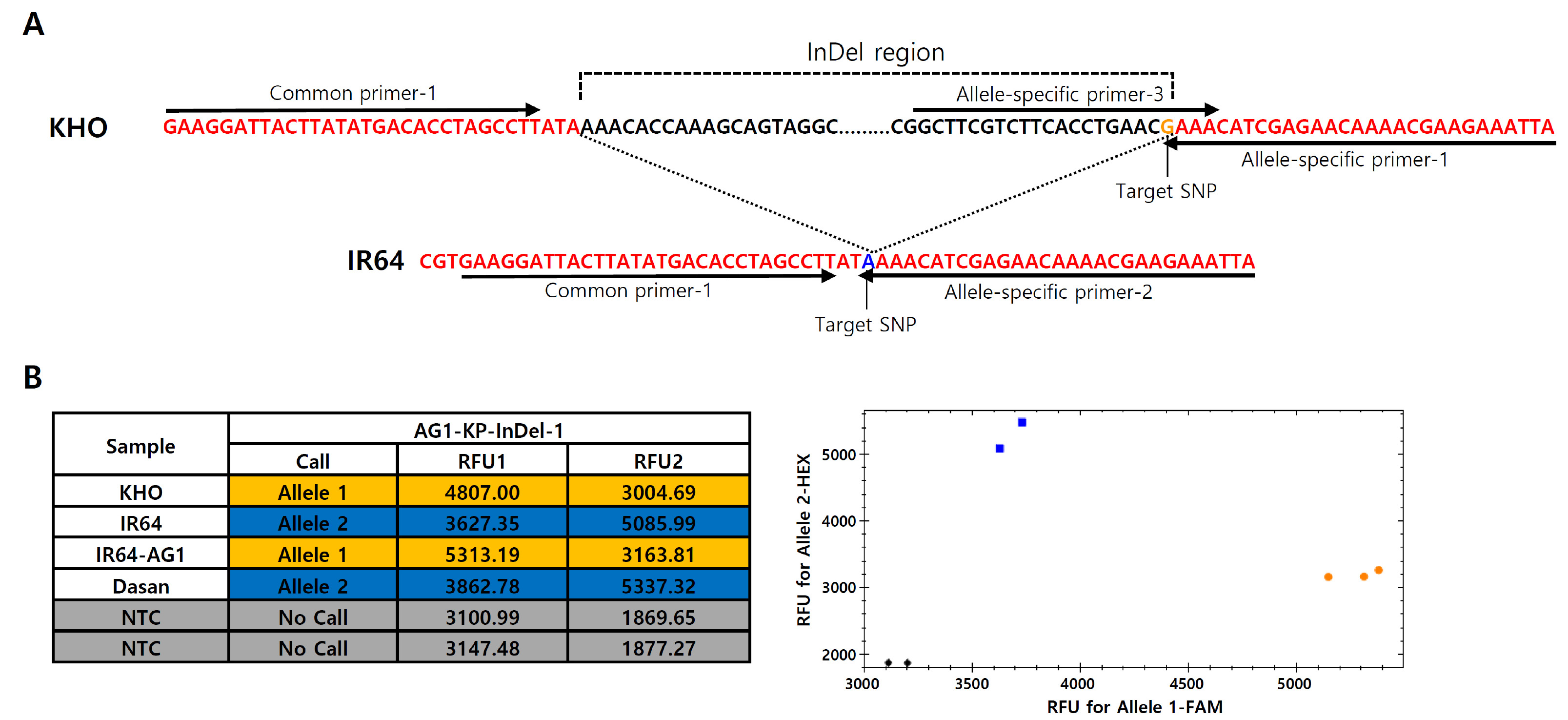
Fig. 3Genomic location of the four AG1 flanking KASP markers and validation of the markers with the control varieties. (A) Genomic position of the AG1 InDel and four flanking SNPs that were used to develop the flanking KASP markers, AG1-KP-SNP-1, AG1-KP-SNP-2, AG1-KP-SNP-3 and AG1-KP-SNP-4 markers. Red arrows represent the position of the KASP markers on chromosome 9. (B) Genotyping results of the control panel by the KASP markers. IR64-A, P and S represent IR64 harboring AG1, Pup1 and Sub1, respectively.
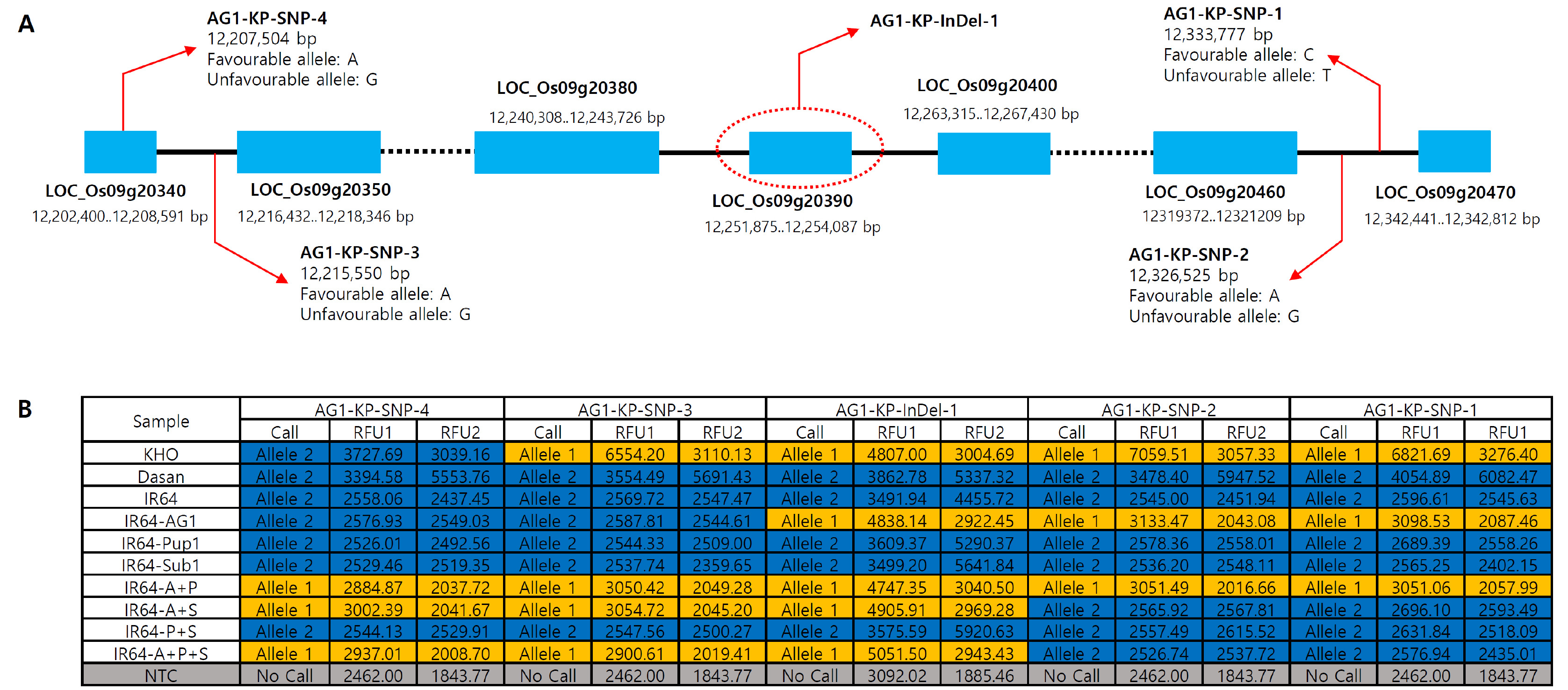
Fig. 4Validation of the existing gel-based DFR marker and three developed KASP markers using BC3F2 segregating lines. (A) Genotyping of 27 BC3F2 segregating lines with the DFR marker. (B) Genotyping of 27 BC3F2 segregating lines with the developed KASP markers. (C) The scatter plot of the KHO, heterozygote and IR64 type alleles. Orange, blue and green dots are allele 1 (tolerant), allele 2 (sensitive) and heterozygote types, respectively. T, tolerant type; S, sensitive type; H, heterozygote.
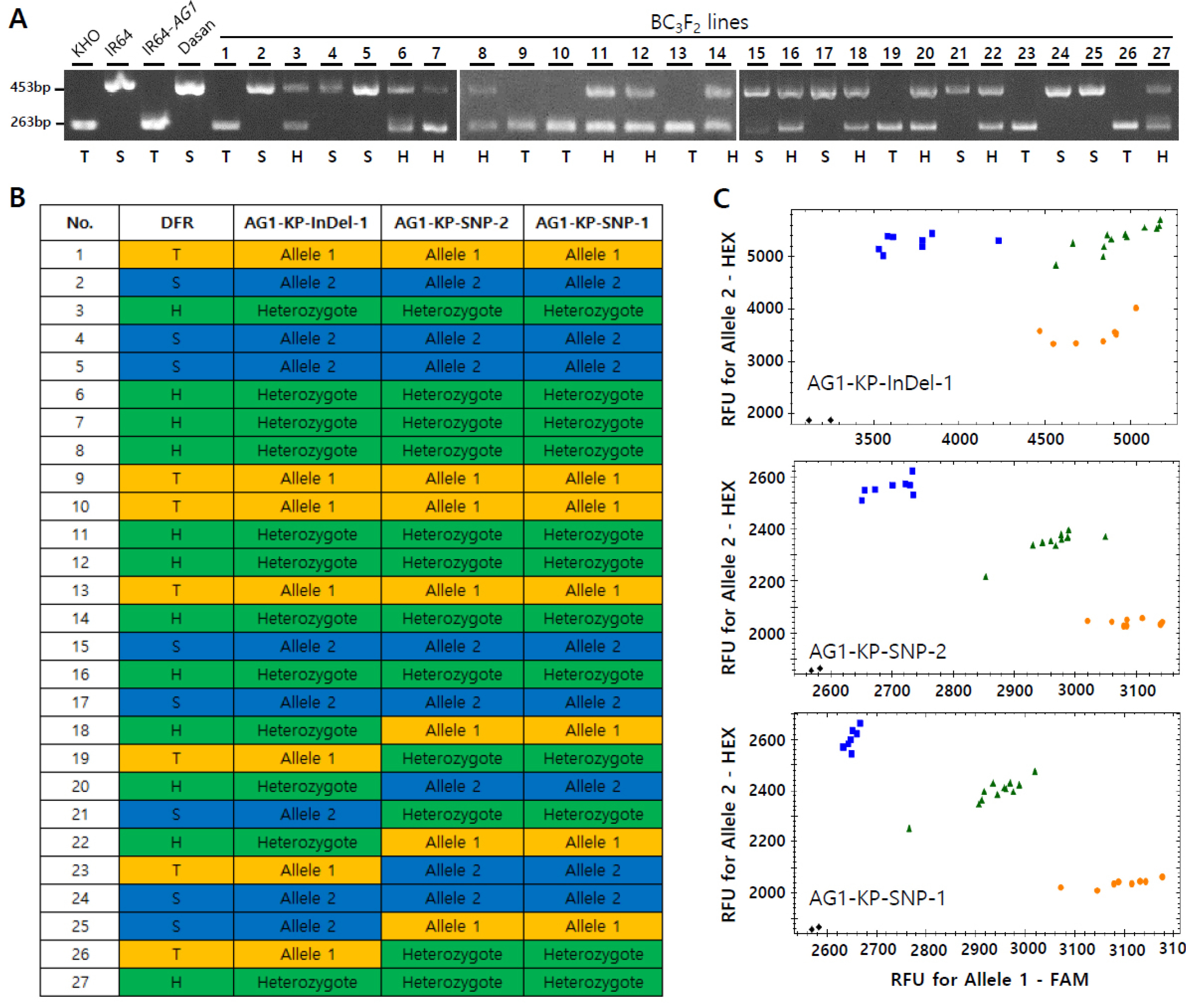
Fig. 5Neighbor joining tree for the profiling of the 172 rice varieties using the five KASP markers (AG1-KP-SNP-1, AG1-KP-SNP-2, AG1-KP-InDel-1, AG1-KP-SNP-3 and AG1-KP-SNP-4). Hollow and solid circles represent Korean and foreign varieties, res-pectively.
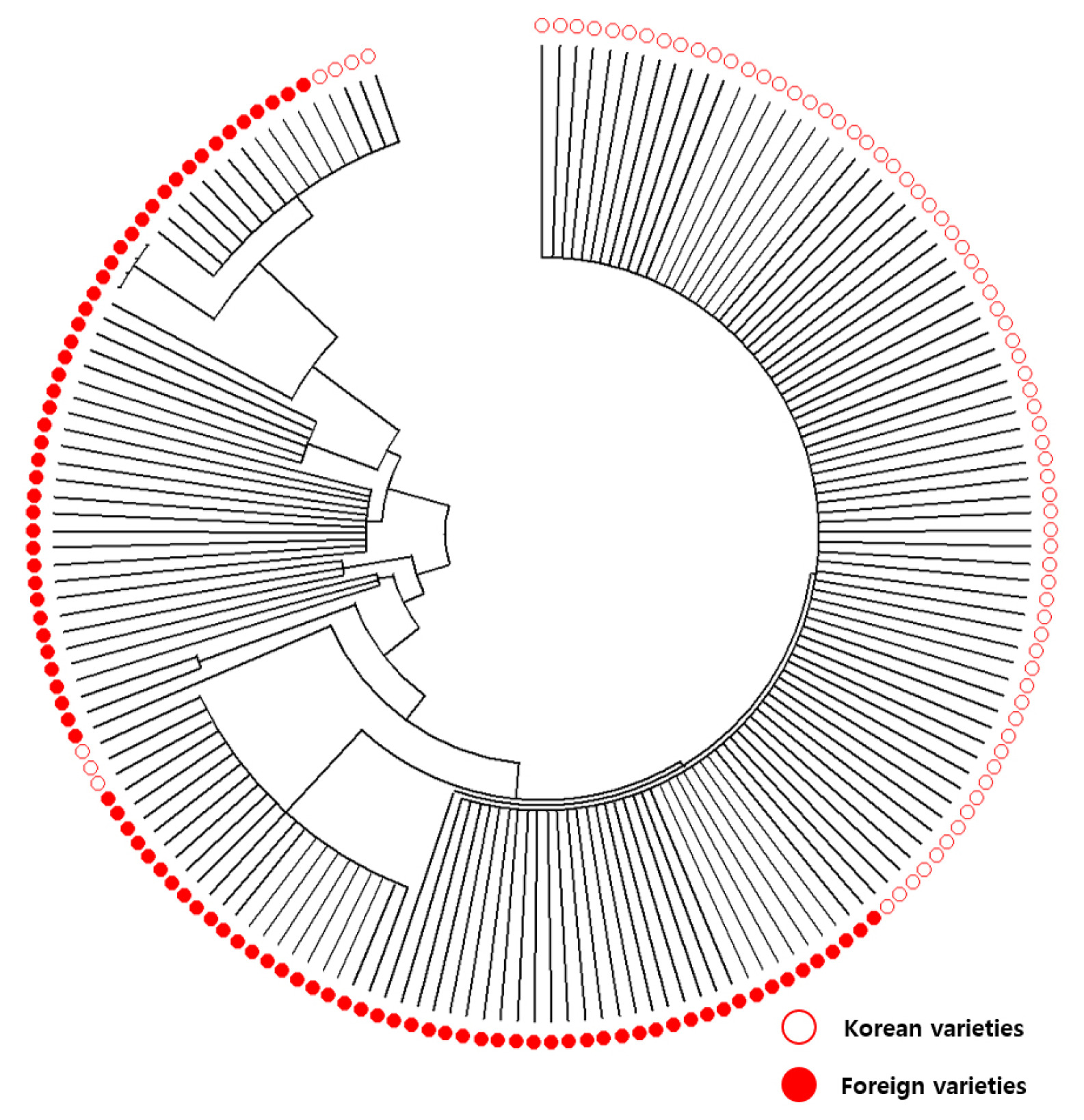
Table 1The list of the primers used in this study.
Table 1
|
Gel-based marker |
Primer name |
Sequence (5ʹ-3ʹ) |
|
DFR |
DFR_F2 |
CCACCATGATGTAGTTCAGTTGTGAAC |
|
DFR_R2 |
CACCGTTAAAATCGGCCGTTAG |
|
DFR_LB2 |
CGGCTTCGTCTTCACCTGAAC |
|
|
KASP marker
|
Primer name
|
Sequence (5ʹ-3ʹ)
|
|
|
AG1-KP-InDel-1 |
Allele-specific primer-1 |
TAATTTCTTCGTTTTGTTCTCGATGTTTC |
|
Allele-specific primer-2 |
AATTTCTTCGTTTTGTTCTCGATGTTTT |
|
Allele-specific primer-3 |
GGCTTCGTCTTCACCTGAACGAAA |
|
Common-primer-1 |
GAAGGATTACTTATATGACACCTAGCCTT |
|
AG1-KP-SNP-1 |
Allele-specific primer |
ACGTTACAATGGTTTGAGTATATGG |
|
Allele-specific primer |
GTCTACGTTACAATGGTTTGAGTATATGA |
|
Common-primer |
TTGTAAGGGTAAGGGTGGGACCTAT |
|
AG1-KP-SNP-2 |
Allele-specific primer |
GAGACGGAGAAGACGGAGAAG |
|
Allele-specific primer |
GGAGACGGAGAAGACGGAGAAA |
|
Common-primer |
CCATCACCGCCAAGAAGTCATCTTA |
Table 2Agreement of the markers calculated by Cohen’s kappa method.
Table 2
|
Markers |
% of consistency with the DFR marker |
Cohen’s Kappa coefficient (k) |
Strength of agreementz)
|
|
AG1-KP-InDel-1 |
100 |
1.00 |
Very good |
|
AG1-KP-SNP-1 |
70.3 |
0.54 |
Moderate |
|
AG1-KP-SNP-2 |
66.7 |
0.48 |
Moderate |
Table 3Profiling of 172 domestic and foreign varieties using developed KASP markers in this study.
Table 3
|
No |
Variety |
Ori |
Marker name |
No |
Variety |
Ori |
Marker name |
|
|
|
M1 |
M2 |
M3 |
M4 |
M5 |
M1 |
M2 |
M3 |
M4 |
M5 |
|
1 |
APO |
FR |
S |
T |
T |
S |
S |
87 |
IR29 |
FR |
T |
T |
T |
T |
T |
|
2 |
Akitakomachi |
FR |
S |
T |
T |
T |
T |
88 |
IR8 |
FR |
S |
S |
S |
S |
S |
|
3 |
MS11 |
FR |
S |
T |
T |
T |
T |
89 |
IR56 |
FR |
S |
S |
S |
S |
S |
|
4 |
Nipponbare |
FR |
S |
T |
T |
T |
T |
90 |
IR36 |
FR |
S |
T |
T |
S |
S |
|
5 |
Koshihikari |
FR |
S |
T |
T |
T |
T |
91 |
IR24 |
FR |
S |
T |
T |
S |
S |
|
6 |
Hitomebore |
FR |
S |
T |
T |
T |
T |
92 |
IR64 |
FR |
S |
S |
S |
S |
S |
|
7 |
Azucena |
FR |
S |
T |
T |
T |
T |
93 |
IR72 |
FR |
S |
T |
T |
T |
T |
|
8 |
Ausboro |
FR |
S |
T |
T |
S |
S |
94 |
IR49830 |
FR |
T |
S |
S |
S |
S |
|
9 |
Aus299 |
FR |
S |
T |
T |
S |
S |
95 |
Chucheongbyeo |
KO |
S |
T |
T |
T |
T |
|
10 |
ARC11544 |
FR |
S |
T |
T |
T |
T |
96 |
Yeonghojinmi |
KO |
S |
T |
T |
T |
T |
|
11 |
Aswina |
FR |
T |
T |
T |
S |
S |
97 |
Junam |
KO |
S |
T |
T |
T |
T |
|
12 |
Aus261 |
FR |
S |
T |
T |
T |
T |
98 |
Ilmi |
KO |
S |
T |
T |
T |
T |
|
13 |
CO39 |
FR |
S |
T |
T |
S |
S |
99 |
Nampyeong |
KO |
S |
T |
T |
T |
T |
|
14 |
Chahora 144 |
FR |
S |
T |
T |
T |
T |
100 |
Samkwang |
KO |
S |
T |
T |
T |
T |
|
15 |
BR11 |
FR |
S |
T |
T |
S |
S |
101 |
Ilpum |
KO |
S |
T |
T |
T |
T |
|
16 |
Basmati 370 |
FR |
S |
T |
T |
S |
S |
102 |
Hopum |
KO |
S |
T |
T |
T |
T |
|
17 |
Basmati bahar |
FR |
T |
T |
T |
T |
T |
103 |
Haiami |
KO |
S |
T |
T |
T |
T |
|
18 |
Binadhan10 |
FR |
T |
T |
T |
S |
S |
104 |
Dongjin 1 |
KO |
S |
T |
T |
T |
T |
|
19 |
Fedearroz50 |
FR |
T |
T |
T |
T |
T |
105 |
Sindongjin |
KO |
S |
T |
T |
T |
T |
|
20 |
Dular |
FR |
T |
T |
T |
T |
T |
106 |
Dongjin |
KO |
S |
T |
T |
T |
T |
|
21 |
Dom sofid |
FR |
S |
T |
T |
T |
S |
107 |
Gopum |
KO |
S |
T |
T |
T |
T |
|
22 |
ColombiaXXI |
FR |
S |
S |
S |
S |
S |
108 |
Seonong 1 |
KO |
S |
T |
T |
T |
T |
|
23 |
Crdhan 405 |
FR |
S |
T |
T |
NC |
NC |
109 |
Seonong 2 |
KO |
S |
T |
T |
T |
T |
|
24 |
Cypress |
FR |
S |
T |
T |
T |
T |
110 |
Seonong 5 |
KO |
S |
T |
T |
T |
T |
|
25 |
Kalarata 1-24 |
FR |
T |
T |
T |
T |
T |
111 |
Seonong 4 |
KO |
S |
T |
T |
T |
T |
|
26 |
IRBB66 |
FR |
S |
T |
T |
S |
S |
112 |
Seonong 3 |
KO |
S |
T |
T |
T |
T |
|
27 |
Inia Tacuri |
FR |
S |
T |
T |
T |
T |
113 |
Seonong 6 |
KO |
S |
T |
T |
T |
T |
|
28 |
FL478 |
FR |
T |
T |
T |
T |
T |
114 |
Seonong 7 |
KO |
S |
T |
T |
T |
T |
|
29 |
Giza 178 |
FR |
T |
T |
T |
T |
T |
115 |
Seonong 8 |
KO |
S |
T |
T |
T |
T |
|
30 |
IAC 165 |
FR |
S |
T |
T |
T |
T |
116 |
Seonong 11 |
KO |
S |
T |
T |
T |
T |
|
31 |
Komboka |
FR |
T |
T |
T |
S |
S |
117 |
Seonong 10 |
KO |
S |
T |
T |
T |
T |
|
32 |
Kharsu 80A |
FR |
T |
T |
T |
T |
S |
118 |
Seonong 9 |
KO |
S |
T |
T |
T |
T |
|
33 |
Kasalath |
FR |
S |
T |
T |
S |
S |
119 |
Seonong 12 |
KO |
S |
T |
T |
T |
T |
|
34 |
Khaiyan |
FR |
S |
NC |
T |
NC |
NC |
120 |
Seonong 13 |
KO |
S |
T |
T |
T |
T |
|
35 |
Khao Lhan On |
FR |
S |
T |
T |
T |
T |
121 |
Seonong 14 |
KO |
S |
T |
T |
T |
T |
|
36 |
Minghui 63 |
FR |
T |
T |
T |
T |
T |
122 |
Seonong 17 |
KO |
S |
T |
T |
T |
T |
|
37 |
MatatAG1 |
FR |
T |
T |
T |
T |
T |
123 |
Seonong 16 |
KO |
S |
T |
T |
T |
T |
|
38 |
Makassane |
FR |
S |
T |
T |
T |
T |
124 |
Seonong 15 |
KO |
S |
T |
T |
T |
T |
|
39 |
Lijiangxin-tuanheigu |
FR |
S |
T |
T |
T |
T |
125 |
Seonong 18 |
KO |
S |
T |
T |
T |
T |
|
40 |
M202 |
FR |
S |
T |
T |
T |
T |
126 |
Seonong 19 |
KO |
S |
T |
T |
T |
T |
|
41 |
Madabaru |
FR |
T |
T |
T |
T |
S |
127 |
Seonong 20 |
KO |
S |
T |
T |
T |
T |
|
42 |
Phka Rumdoul |
FR |
T |
T |
T |
T |
T |
128 |
Hwayong |
KO |
S |
T |
T |
T |
T |
|
43 |
Osgovka |
FR |
S |
T |
T |
T |
T |
129 |
Chilbo |
KO |
S |
T |
T |
T |
T |
|
44 |
NSIC RC 238 |
FR |
T |
S |
S |
S |
S |
130 |
Seonong 21 |
KO |
S |
T |
T |
T |
T |
|
45 |
Moroberekan |
FR |
S |
T |
T |
T |
T |
131 |
Odae |
KO |
S |
T |
T |
T |
T |
|
46 |
N22 (heat) |
FR |
T |
T |
T |
T |
T |
132 |
Unkwang |
KO |
S |
T |
T |
T |
T |
|
47 |
NSIC RC 222 |
FR |
T |
S |
S |
S |
S |
133 |
Obong |
KO |
S |
T |
T |
T |
T |
|
48 |
Rinaldo bersani |
FR |
S |
T |
T |
T |
T |
134 |
Milyang 23 |
KO |
S |
S |
S |
S |
S |
|
49 |
Rayada |
FR |
T |
T |
T |
S |
S |
135 |
Jinmi |
KO |
S |
T |
T |
T |
T |
|
50 |
PSB RC 82 |
FR |
S |
T |
S |
S |
S |
136 |
Hwacheongbyeo |
KO |
S |
T |
T |
T |
T |
|
51 |
Sabri |
FR |
S |
T |
T |
T |
T |
137 |
Dasan |
KO |
S |
S |
S |
S |
S |
|
52 |
Saducho |
FR |
S |
T |
T |
S |
S |
138 |
Hanmaeum |
KO |
S |
T |
T |
T |
T |
|
53 |
Samba Mahsuri |
FR |
S |
S |
S |
S |
S |
139 |
Boramchan |
KO |
S |
T |
T |
T |
T |
|
54 |
Taducan |
FR |
T |
T |
T |
S |
S |
140 |
Milkyqueen |
KO |
S |
T |
T |
T |
T |
|
55 |
Swarna |
FR |
S |
T |
T |
T |
S |
141 |
Bakjinju |
KO |
S |
T |
T |
T |
T |
|
56 |
San-Huang-Zhau No2 |
FR |
T |
T |
T |
T |
T |
142 |
Boseogchal |
KO |
S |
T |
T |
T |
T |
|
57 |
Tai |
FR |
T |
S |
S |
S |
S |
143 |
Dongjinchal |
KO |
S |
T |
T |
T |
T |
|
58 |
Tainung 67 |
FR |
S |
T |
T |
T |
T |
144 |
Baegokchal |
KO |
S |
T |
T |
T |
T |
|
59 |
Tong 88-7 |
FR |
S |
T |
T |
T |
T |
145 |
Aromi |
KO |
S |
T |
T |
T |
T |
|
60 |
Vandana |
FR |
S |
T |
T |
T |
T |
146 |
Heugkwang |
KO |
S |
T |
T |
T |
T |
|
61 |
Utri Merah |
FR |
T |
T |
T |
T |
T |
147 |
Daerip 1 |
KO |
S |
T |
T |
T |
T |
|
62 |
Vuninzara |
FR |
S |
S |
S |
S |
S |
148 |
Geunnun |
KO |
S |
T |
T |
T |
T |
|
63 |
WAB 56-125 |
FR |
T |
T |
T |
T |
T |
149 |
Jeogjinju |
KO |
S |
T |
T |
T |
T |
|
64 |
Way Rarem |
FR |
S |
T |
T |
S |
T |
150 |
Miho |
KO |
S |
T |
T |
T |
T |
|
65 |
Kusahonami |
FR |
S |
T |
T |
T |
T |
151 |
Sobi |
KO |
S |
S |
S |
S |
S |
|
66 |
Phka Rumduol |
FR |
T |
T |
T |
T |
T |
152 |
Shinongheugchal |
KO |
S |
T |
T |
T |
T |
|
67 |
G Zhenshan 97B |
FR |
T |
T |
T |
T |
T |
153 |
Mipum |
KO |
S |
T |
T |
T |
T |
|
68 |
Hoshiaoba |
FR |
S |
T |
T |
T |
T |
154 |
Haedamssal |
KO |
S |
T |
T |
T |
T |
|
69 |
Be Ko Aoba |
FR |
S |
T |
T |
T |
T |
155 |
Joami |
KO |
S |
T |
T |
T |
T |
|
70 |
Hukuhibiki |
FR |
S |
T |
T |
T |
T |
156 |
Hongjinju |
KO |
S |
T |
T |
T |
T |
|
71 |
TR22183 |
FR |
S |
T |
T |
T |
T |
157 |
Heugjinju |
KO |
S |
T |
T |
T |
T |
|
72 |
Inia tacuari |
FR |
S |
T |
T |
T |
T |
158 |
Saenuri |
KO |
S |
T |
T |
T |
T |
|
73 |
Adday sel |
FR |
T |
T |
T |
S |
S |
159 |
Jeogjinjuchal |
KO |
S |
T |
T |
T |
T |
|
74 |
Nona Bokra |
FR |
T |
T |
T |
T |
S |
160 |
Seilmi |
KO |
S |
T |
T |
T |
T |
|
75 |
A69-1 |
FR |
S |
S |
S |
S |
S |
161 |
Cheongwoo |
KO |
T |
T |
T |
T |
T |
|
76 |
Hasawi |
FR |
T |
T |
T |
T |
T |
162 |
Yeongwoo |
KO |
T |
T |
T |
T |
T |
|
77 |
IR64-SPIKE |
FR |
S |
S |
S |
S |
S |
163 |
Jonong |
KO |
S |
T |
T |
T |
T |
|
78 |
IR64-Saltol |
FR |
S |
S |
S |
S |
S |
164 |
Nokwoo |
KO |
S |
T |
T |
T |
T |
|
79 |
Pokkali |
FR |
T |
T |
T |
T |
T |
165 |
Mokyang |
KO |
S |
S |
S |
S |
S |
|
80 |
IR64-EMF |
FR |
S |
S |
S |
S |
S |
166 |
Nokyang |
KO |
S |
T |
T |
T |
T |
|
81 |
IR64-AG1 |
FR |
S |
S |
T |
T |
T |
167 |
Sanggol |
KO |
S |
T |
T |
T |
T |
|
82 |
IR64-Pup1 |
FR |
S |
S |
S |
S |
S |
168 |
Hanareum 2 |
KO |
S |
T |
T |
T |
T |
|
83 |
IR64-HTSF4.1(73) |
FR |
T |
T |
T |
T |
T |
169 |
Mogwoo |
KO |
T |
T |
T |
T |
T |
|
84 |
IR64-DTY2.2+DTY4.1 |
FR |
S |
S |
S |
S |
S |
170 |
Namwon1 |
KO |
S |
T |
T |
T |
T |
|
85 |
IR64-Sub1 |
FR |
S |
S |
S |
S |
S |
171 |
Jinbubyeo |
KO |
S |
T |
T |
T |
T |
|
86 |
IR28 |
FR |
T |
T |
T |
T |
T |
172 |
Koryeong13 |
KO |
S |
T |
T |
T |
S |
References
- Altman DG. 1990. Practical statistics for medical research. CRC press. Boca Raton, FL, U. S. A:
- Angaji SA, Septiningsih EM, Mackill DJ, Ismail AM. 2010. QTLs associated with tolerance of flooding during germination in rice (Oryza sativa). Euphytica. 172: 159-168.
- Baltazar MD, Ignacio JCI, Thomson MJ, Ismail AM, Mendioro MS, Septiningsih EM. 2014. QTL mapping for tolerance of anaerobic germination from IR64 and the aus landrace Nanhi using SNP genotyping. Euphytica. 197: 251-260.
- Frisch M, Bohn M, Melchinger AE. 1999. Comparison of selection strategies for marker-assisted backcrossing of a gene. Crop Sci.. 39: 1295-1301.
- Ge LF, Chao DY, Shi M, Zhu MZ, Gao JP, Lin HX. 2008. Overexpression of the trehalose-6-phosphate phosphatase gene OsTPP1 confers stress tolerance in rice and results in the activation of stress responsive genes. Planta. 228: 191-201.
- Hsu SK, Tung CW. 2015. Genetic mapping of anaerobic germination-associated QTLs controlling coleoptile elongation in rice. Rice (N. Y.). 8: 38
- Jiang L, Hou M, Wang C, Wan J. 2004. Quantitative trait loci and epistatic analysis of seed anoxia germinability in rice (Oryza sativa L.). Rice Sci.. 11: 238-244.
- Jiang L, Liu S, Hou M, Tang J, Chen L, Zhai H, et al. 2006. Analysis of QTLs for seed low temperature germinability and anoxia germinability in rice (Oryza sativa L.). Field Crops Res.. 98: 68-75.
- Kim JH, Mo YJ, Ha SK, Kim WJ, Kim BK, Jeung JU, et al. 2019. QTL analysis for tolerance to anaerobic germination in japonica rice (Oryza sativa L.). Korean J. Breed. Sci.. 51: 415-423.
- Kretzschmar T, Pelayo MAF, Trijatmiko KR, Gabunada LFM, Alam R, Jimenez R, et al. 2015. A trehalose-6-phosphate phosphatase enhances anaerobic germination tolerance in rice. Nat. Plants. 1: 1-5.
- Landis JR, Koch GG. 1977. The measurement of observer agreement for categorical data. Biometrics. 33: 159-174.
- Lunn JE, Delorge I, Figueroa CM, Dijck PV, Stitt M. 2014. Trehalose metabolism in plants. Plant J.. 79: 544-567.
- Moon JH, Son D, Lee JW, Yoo SC. 2019. Development of Kompetitive Allele Specific PCR markers for submergence tolerant gene Sub1 in rice. Plant Breed. Biotech.. 7: 62-66.
- Murray MG, Thompson WF. 1980. Rapid isolation of high molecular weight plant DNA. Nucleic Acids Res.. 8: 4321-4326.
- Pandey S, Mortimer M, Wade L, Tuong TP, Lopez K, Hardy B. 2002. Direct seeding: research strategies and opportunities. In: Proceedings of the international workshop on direct seeding in Asia rice systems: strategic research issues and opportunities; 25-28 January 2000; Bangkok, Thailand. International Rice Research Institute. Los Banos, Philippines, pp 383.
- Platten JD, Cobb JN, Zantua RE. 2019. Criteria for evaluating molecular markers: Comprehensive quality metrics to improve marker-assisted selection. PLoS One. 14: e0210529
- Ramkumar G, Prahalada GD, Hechanova SL, Vinarao R, Jena KK. 2015. Development and validation of SNP-based functional codominant markers for two major disease resistance genes in rice (O. sativa L.). Mol. Breed.. 35: 1-11.
- Semagn K, Babu R, Hearne S, Olsen M. 2014. Single nucleotide polymorphism genotyping using Kompetitive Allele Specific PCR (KASP): overview of the technology and its application in crop improvement. Mol. Breed.. 33: 1-14.
- Septiningsih EM, Ignacio JCI, Sendon PMD, Sanchez DL, Ismail AM, Mackill DJ. 2013. QTL mapping and confirmation for tolerance of anaerobic conditions during germination derived from the rice landrace Ma-Zhan Red. Theor. Appl. Genet.. 126: 1357-1366.
- Shima S, Matsui H, Tahara S, Imai R. 2007. Biochemical characterization of rice trehalose-6-phosphate phosphatases supports distinctive functions of these plant enzymes. FEBS J.. 274: 1192-1201.
- Tan CT, Yu H, Yang Y, Xu X, Chen M, Rudd JC, et al. 2017. Development and validation of KASP markers for the greenbug resistance gene Gb7 and the Hessian fly resistance gene H32 in wheat. Theor. Appl. Genet.. 130: 1867-1884.
- Tanksley SD, Young ND, Patterson AH, Bonierbale MW. 1989. RFLP mapping in plant breeding: new tools for an old science. Nat. Biotechnol.. 7: 257-264.
- Yadav UP, Ivakov A, Feil R, Duan GY, Walther D, Giavalisco P, et al. 2014. The sucrose-trehalose 6-phosphate (Tre6P) nexus: specificity and mechanisms of sucrose signalling by Tre6P. J. Exp. Bot.. 65: 1051-1068.
- Yang J, Sun K, Li D, Luo L, Liu Y, Huang M, et al. 2019. Identification of stable QTLs and candidate genes involved in anaerobic germination tolerance in rice via high-density genetic mapping and RNA-Seq. BMC Genomics. 20: 1-15.
- Zhao S, Li A, Li C, Xia H, Zhao C, Zhang Y, et al. 2016. Development and application of KASP marker for high throughput detection of AhFAD2 mutation in peanut. Electron. J. Biotechnol.. 25: 9-12.