Abstract
Xanthomonas campestris pv. incanae (Xci) is the causal agent of bacterial blight disease in ornamental crucifers. We compared the whole genomes of closely related Xanthomonas campestris pv. campestris, incanae, raphani and four other species of Xanthomonas following comparative genomics approach. We found 82 singletons out of 4024 Xci genes upon comparison. Out of 82 singletons, top 10 were selected for designing Xci specific marker. Five primers; XCI_1F/R, XCI_2F/R, XCI_3F/R, XCI_5F/R and XCI_6F/R produced amplicons of 495 bp, 503 bp, 612 bp, 665 bp and 468 bp, respectively expected to detect Xanthomonas campestris pv. incanae (Xci). In conclusion, five effective markers were developed for the detection of Xci pathogen by whole genome alignment, which could be served as effective tools in seed quarantine.
-
Key words: Bacterial blight, X. campestris pv. incanae, Comparative genomics, Molecular markers
INTRODUCTION
Xanthomonas campestris is a gram negative rod-shaped bacterium, which infects a wide range of host plants. It has been classified into three pathovars (Pv.) namely
campestris,
raphani and
incanae based on host specificity (
Fargier and Manceau 2007;
Roux et al. 2015). The race structures of
Xanthomonas campestris pv.
campestric (
Xcc) and
Xanthomonas campestris pv.
raphani (
Xcr) were confirmed through gene-for-gene model by determining relationship between host plant and pathogen. So far, 11
Xcc and three
Xcr pathogenic races have been identified (
Fargier and Manceau 2007;
Vicente and Holub 2013). Till now, race structure of
Xci remains unreported (
Vicente et al. 2002;
Fargier and Manceau 2007).
Bacterial blight is a major disease of ornamental crops, such as stock (
Matthiola incana), wallflower (
Erysimum cheiri) and
candytuft (
Iberis umbellata) (
Pirone 1978;
Vicente et al. 2001) of which are economically important in Holland, Italy, Japan, China, Australia and Israel (
Ecker et al. 1995).
Xci causes bacterial blight is strictly host-specific compared to the pathovars of
Xcc and
Xcr.
Xci is either a seed- or soil-borne pathogen, which penetrates through wound or leaf hydathodes, and invades the vascular system of the flowering plant (
Kendeick and Baker 1942;
Williams 1980). The disease is readily get transmitted among healthy plants through water splash. The leaves show yellowing and wilting upon transmission (
Fargier and Manceau 2007;
Cruz et al. 2017). Initially symptoms arise on the main stem and lateral branches of leaf and gradually reaches leaf petioles and seed peduncles (
Rahimian and Okhovatian 1989). The disease may cause 90% of total yield loss in ornamental crops (
Minardi et al. 1988). Therefore, disease management is crucial to check the crop loss. Early diagnosis of the disease and its causal agent is vital for effective disease management. Although molecular techniques, like genomic fingerprinting (repetitive PCR) helps to distinguish
X. campestris isolates at pathovar level (
Vicente et al. 2006;
Parkinson et al. 2009). Such technique requires several repetitions to obtain accuriate results. Recently, Multi-Locus Sequence Typing (MLST) has been used to determine the evolutionary relationship of different
Xcc populations (
Fargier et al. 2011;
Cruz et al. 2018;
Laala et al. 2021). But, MLST requires systemic steps such as data collection, data analysis and finally multi-loci sequence analysis. It is expensive, time consuming, need skilled personnel and sometimes unspeculated results could make ambiguity in distinguishing the bacterial strains (
Zhang et al. 2004). Therefore, there is a need to adopt a new or an improved older technique for the robust detection of
Xci in host plants.
Comparative genomic approach represents direct comparison of the collective genomes of a particular group of organisms either across the genus or species level. It reveals the similarities, differences and evolutionary patterns among the diverse organisms at genomic level (
Abby and Daubin 2007;
Sivashankari and Shanmughavel 2007). This approach was successful in understanding the evolutionary mechanism of
Bacillus licheniformis, the comparison of homologous genes among
Drosophila melanogaster and
Homo sapiens (
Rey et al. 2004). The term ‘pangenomics’ was introduced in the study of collective gene content of an organisms from the same genus or species (
Garrigues et al. 2013). Pangenome analysis focusses on the functional distribution of genes into core genome (genes are common in all individuals), dispensable genome (genes are absent in at least one individual) or singletons (genes are found in only one individual) (
Tettelin et al. 2005;
Mira et al. 2010). Pangenome is significant in an evolutionary context (
Reno et al. 2009) but it could also be used in a broader genomic aspect (
Reinhardt et al. 2009). The importance of pangenome analysis is accomplished herein to identify genes or regions for developing
Xci-specific marker.
To date, no molecular marker has been reported to detect
Xci strain. PCR (polymerase chain reaction) techniques are rapid, sensitive, reliable and economical in the specific identification of the pathogens (
Berg et al. 2005;
Kałużna et al. 2014). Utilizing PCR-based molecular marker might be a precise method in the detection of
Xci as like the previously reported
Xcc (
Rubel et al. 2019b),
Xcc-race specific (
Rubel et al. 2017;
Afrin et al. 2018) and
Xcr specific markers (
Rubel et al. 2019a). In the
Xci, type III effector (T3E) proteins are very essential for it’s virulence (
Roux et al. 2015) to the host plants, which has been focused in this study for marker development. So far, the genomes of two
Xci strains (CFBP 1606R and CFBP 2527R) are available and used in present study. The aim of this study is to develop an
Xci-specific molecular marker to recognize them from other bacterial species. In this regard, the complete genomes of
Xci (CFBP1606R and CFBP2527R) was compared across
Xcc (B100, ATCC33913 CFBP5817, 8004 and CN17),
Xcr (756C) and other
Xanthomonas genomes using EDGAR 3.0 (Electronic Data Gathering, Analysis, and Retrieval) (
https://edgar3.computational.bio.uni-giessen.de/cgi-bin/edgar_login.cgi) to harvest singleton genes from
Xci genome for developing pathovar-specific markers.
Xci-specific markers could be facilitated to the farmers in identifying the
Xci strain within a short time for taking appropriate control measures against bacterial blight disease in ornamental crucifers.
MATERIALS AND METHODS
Culture of bacterial strains
A total of 18 diverse bacterial strains was collected from different sources and used in this study (
Supplementary Table S1). Bacterial strains were grown on King’s B media at 30°C for 48 hours. All bacterial strains were stored at –80°C in a liquid medium with 50% glycerol for further uses.
Bacterial DNA was extracted from cultured bacterial cells using DNeasy Plant Mini Kit (QIAGEN, Hilden, Germany) following the manufacturer’s instruction. The quality of isolated DNA was checked using 1.2% agarose gel electrophoresis at 100 V and quantified using a NanoDropND-1000 spectrophotometer (NanoDrop, Wilmington, DE, USA).
Comparative genome analysis of Xcc, Xcr and Xci
Comparative genome analysis was done by using EDGAR 3.0 (
Blom et al. 2009;
Blom et al. 2016;
Dieckmann et al. 2021) to understand the similarities and variations among the bacterial species. The genomes of
Xcc (B100, ATCC33913, CFBP5817, CN14, CN15, CN16, 8009),
Xcr (756C),
Xci (CFBP2527R, CFBP1606R),
X. axonopodis pv.
campestris,
X. euvesicatoria,
X. axonopodis pv.
diffenbachiae (LMG695),
X. citri pv.
glycines (CFBP2526) were collected from NCBI (National Center for Biotechnology Information) and used for comparative genome analysis (
Table 1).
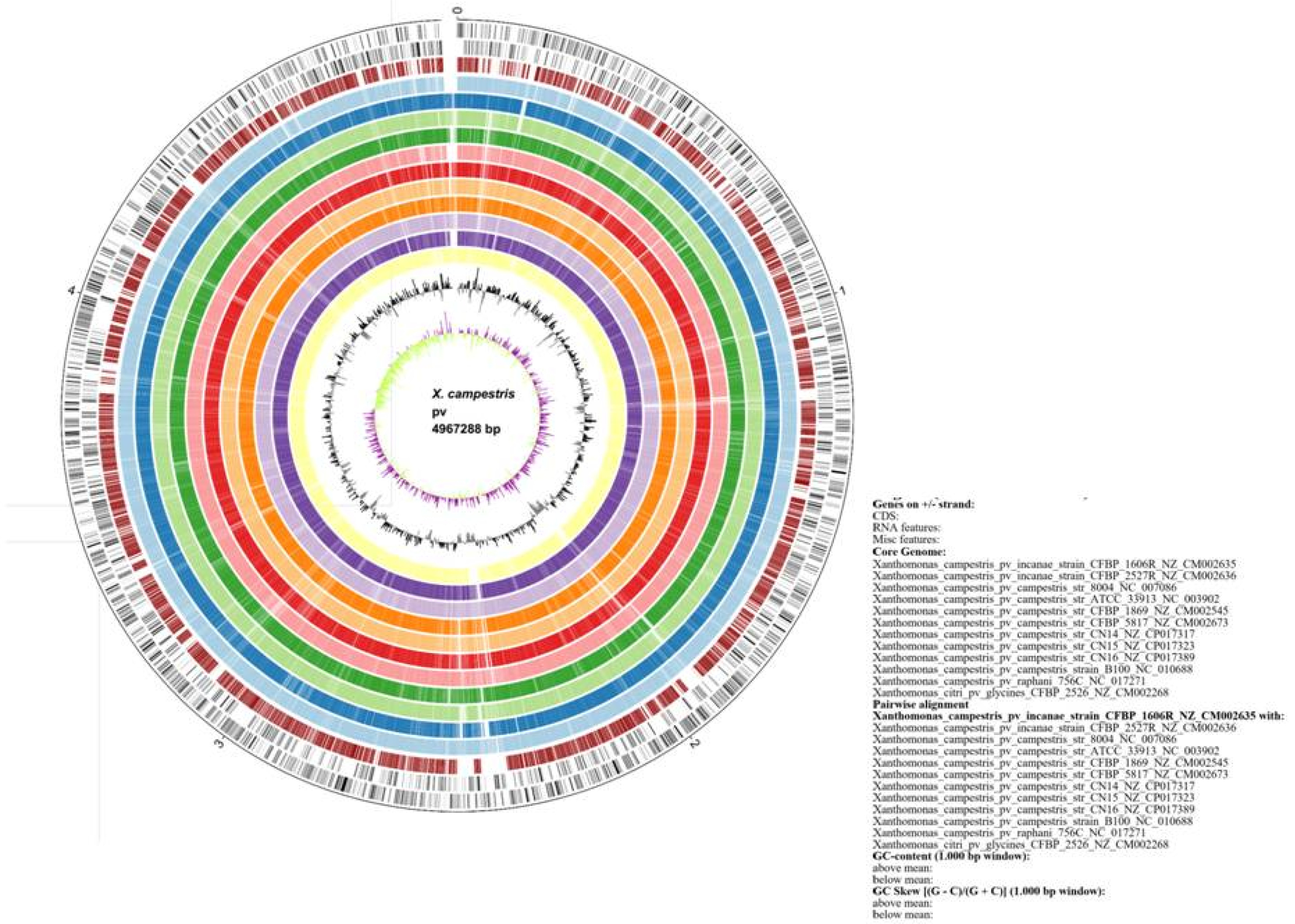
The EDGAR tool produced information on core genome, dispensable genome and singletons; which were considered for marker development. In addition, genome plot and the gene cluster (
Fig. 1) representation were also obtained from the EDGAR tool.
Xci specific singleton genes with a T3E were selected for designing primers. Primers were designed from ten randomly selected singletons genes using primer3plus (
http://www.bioinformatics.nl/cgi-bin/primer3plus/primer3plus.cgi). The uniqueness of
Xci-specific genes was confirmed by using the Xcc-BLAST tool (
http://210.110. 86.160/lab/home.html) and BLASTn of NCBI (
www.ncbi. nlm.nih.gov). Total elven primers were designed, among them one primer pair was designed from each of the gene except XCICFBP2527_RS21720, from which two primers were designed due to large gene size. The prefix XCI_1F/R, XCI_2F/R, XCI_3F/R, XCI_4F/R, XCI_5F/R, XCI_6F/R, XCI_7F/R, XCI_8F/R, XCI_9F/R, XCI_10F/ R and XCI_11F/R were denoted for the 11 primers designed from randomly selected 10 singleton genes of
Xci strain (
Table 2).
PCR validation for specific detection of Xci
Specificity of the designed primer was determined by the PCR assay using extracted DNA from strain of each of the
Xci and
Xcr, eight strains from
Xcc, four strains from
Xanthomonas and four other test bacteria (
Supplementary Table S1). PCR was carried out with 20 µL reaction mixtures contained 1.0 µL of forward and 1.0 µL reverse primers (10 pmol), 8.0 µL Emerald PCR master mix (Takara, Shiga, Japan), 9.0 µL ultra-pure water and 1.0 µL DNA (70 ng). The PCR condition was fixed as; the initial denaturation at 95°C for 5 minutes followed by 25 cycles of denaturation at 95°C for 30 seconds with annealing temperatures varied from 57°C to 68°C for 40 seconds and extension at 72°C for 45 seconds and final extension at 72°C for 5 minutes. The amplified PCR products were separated by electrophoresis using 8 µL of PCR products in 1.4% agarose gel dissolved in 1x TBE buffer and stained with HIQ Blue Mango (20,000X) (bioD, Seoul, South Korea) and run for 40 minutes with 100 V before visualizing under UV light (302 nm).
The bacterial DNA sample was diluted sequentially in ten fold increments using sterile double distilled water up to a dilution of 10-9 and fixed the DNA concentration ranged from 7 ng/µL to 0.00000007 ng/µL for detection of the sensitivity level of the well amplified Xci specific primers. From each of the diluted samples, 1 µL DNA was used as a source of template DNA with the similar PCR mixture as well as conditions were described previously.
RESULTS
Xci specific genes from genome comparison
Comparative genomic approach was implied to identify the genetic similarities and variations among the diverse
Xanthomonas subspecies and their strains. The overall description of the bacterial strains; such as genome size (bp), genes per genome/strain, protein per genome/strain and NCBI taxonomic ID are enlisted in the
Table 1. The complete genomes of closely related strains of
Xcc,
Xcr and
Xci were analysed by comparative genomics and a total of 4,024 genes were identifed in the
Xci strain
(
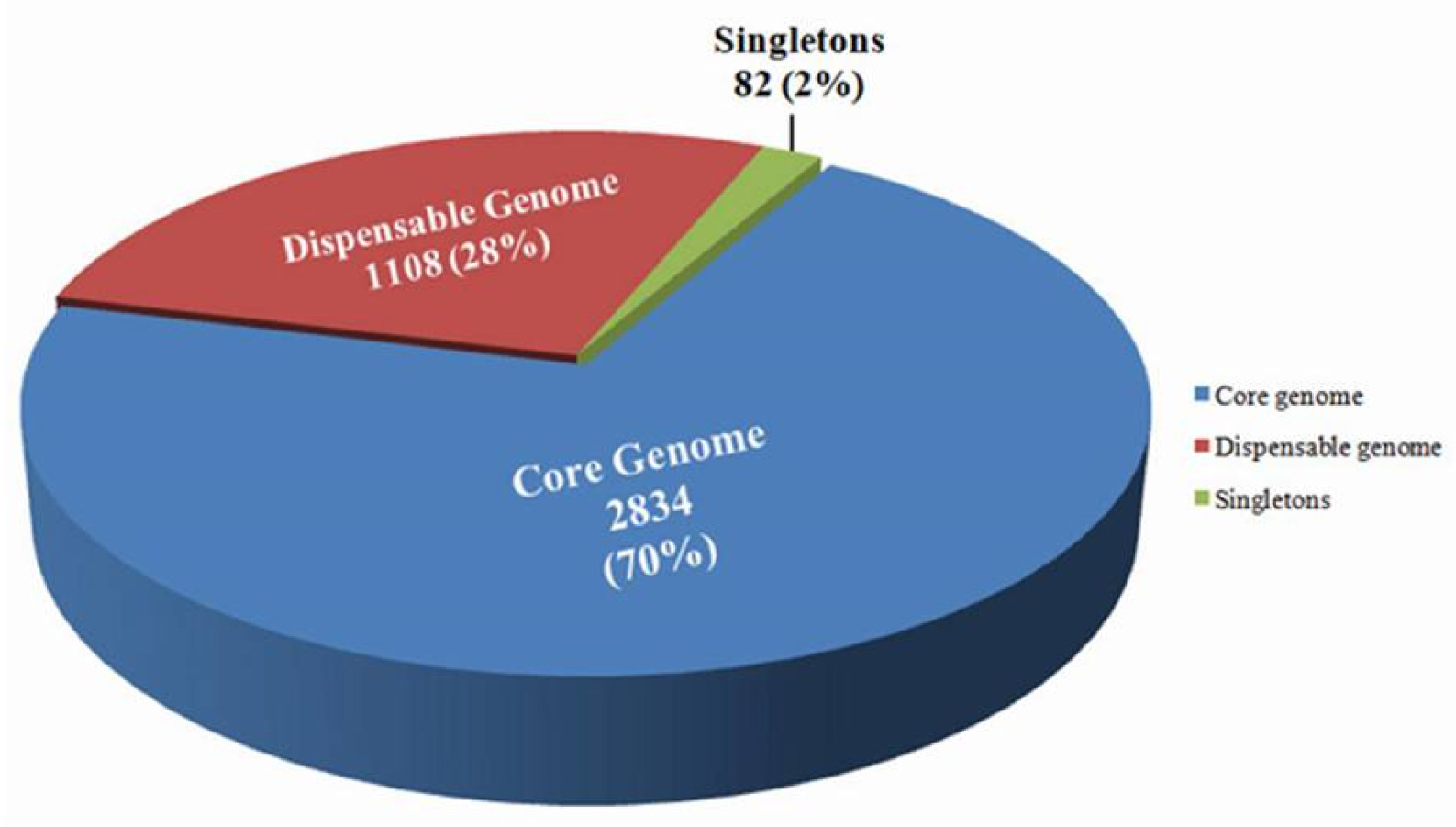
Fig. 2). Comparative genome analysis conferred the segregation of the genomes into three major categories: 1) core genome, 2) dispensable genome and 3) singletons. The core genome consisted of 70% of the total gene (2,834) which are conserved across the genome of the bacterial strains, dispensable or flexible genome contained 1,108 genes which covered about 28% of the total number of genes and the remaining 2% comprised 82 singleton genes, which were found only in
Xci genome (
Fig. 2). These 82 singleton genes were retrieved from EDGAR tool and confirmed the
Xci specificity by
Xcc-BLAST tool and BLASTn of NCBI. Out of 82 singleton genes, only 10 genes were selected randomly for
Xci-specific primer designing. Among the selected genes, only
Xci specific four genes encoded hypothetical proteins and one gene encoded T3E protein (
Table 3).
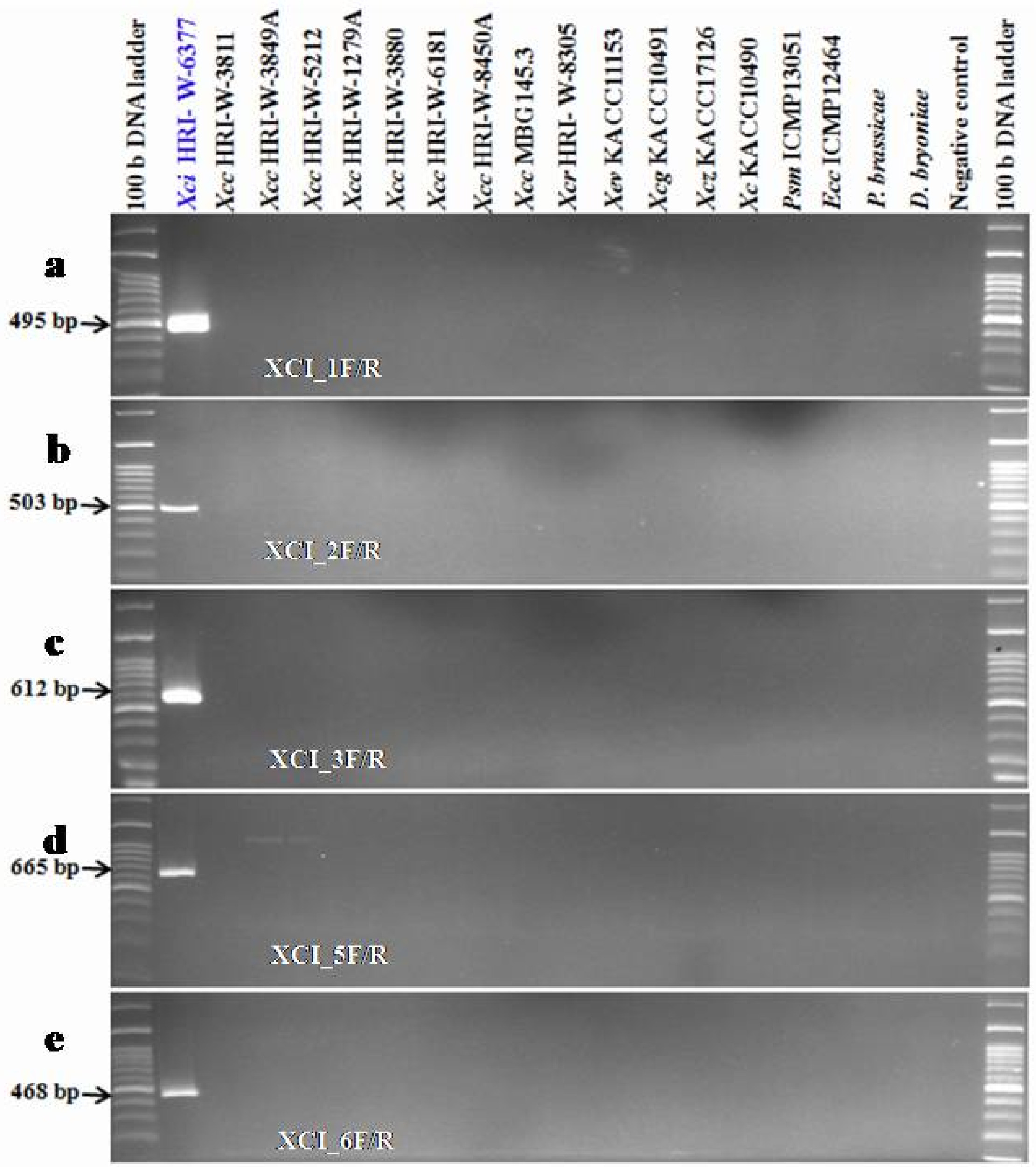
Out of eleven primers, five primer pairs; XCI_1F/R, XCI_2F/R, XCI_3F/R, XCI_5F/R and XCI_6F/R were amplified with expected PCR amplicons of 495 bp, 503 bp, 612 bp, 665 bp and 468 bp, respectively in the Xci population
(
Fig. 3,
Supplementary Table S1). Those amplifications were completely absent in other
Xanthomonas spp. such as
X. campestris pv.
campestris,
X. campestris pv.
raphani,
X. euvesicatoria,
X. axonopodis pv.
glycines,
X. campestris pv.
zinniae,
X. campestris or in
P. syringae pv.
maculicola,
Erwinia carotovora subsp.
carotovora,
Plasmodiophora brassicae and
Didymella bryoniae. These results confirmed that 5 primer sets are specific to
Xci only. The primer sets, XCI_1F/R, XCI_2F/R, XCI_3F/R and XCI_5F/R covered the gene loci of hypothetical proteins;
XCICFBP2527_RS16430,
XCICFBP2527_RS13555,
XCICFBP2527_RS21720 and
XCICFBP2527_RS03935, respectively. Primer set XCI_6F/R tagged T3E protein encoding locus
XCICFBP2527_RS21980. The remaining 6 primers neither produced target amplicons nor even amplified and concluded as redundant primers of
Xci-specific detection.
We identified the orthologous genes and their genomic neighborhood by EDGAR tool. In addition, we identified only the 5 orthologs genes of
Xci rather than any other pathovars of
Xanthomonas genomes. Aparant locations and orientations of the 5 selected genes on
Xanthomonas genome are shown in the
Table 3. Among those genes, two genes are located on positive strand (+) (
XCICFBP2527_ RS13555 and
XCICFBP2527_RS21720) and other three on negative strand (-) (
XCICFBP2527_RS16430,
XCICFB P2527_RS03935 and
XCICFBP2527_RS21980) (
Table 3).
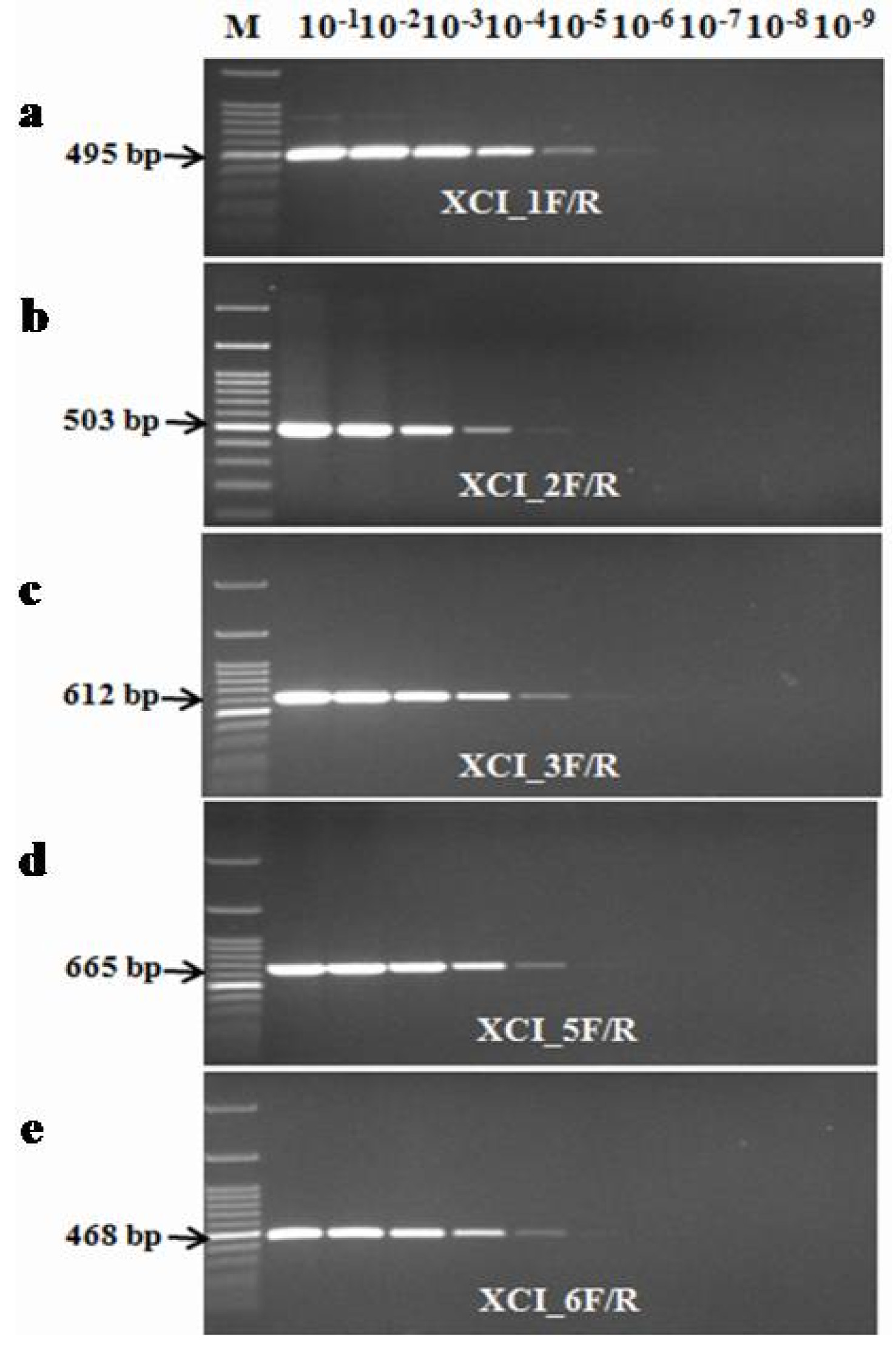
A PCR sensitivity test for Xci specific primers was performed using genomic DNA of Xci strain (HRI-W-6377). The sensitivity of the developed primers was checked using a ten-fold dilution series of genomic DNA. Five primer sets; XCI_1F/R, XCI_2F/R, XCI_3F/R, XCI_5F/R and XCI_6F/R gave effective sensitivity in PCR assay. The expected PCR product was amplified with each primer up to sixth dilution (0.00007 ng/µL) with the exception of one primer pair XCI_2F/R, which resulted amplification up to fifth dilution (0.0007 ng/µL)
(
Fig. 4) and indicated detection limit by PCR.
DISCUSSION
Comparative genomics is an easy and accessible approach, which provides an opportunity for in-depth analysis of intr-aspecies diversity by using the available genome sequences of different strains in the public data bases. Pangenome of bacterial populations has been used since 10 years ago for comparative genomics; currently, several bioinformatics tools have been developed for comparative genome analysis. However, a fewer online tools is available for comparative genome analysis, especially for bacterial genomes. Among them, EDGAR 3.0 is one of the most popular webserver based tool for the comparative bacterial genome study (
Blom et al. 2009;
Blom et al. 2016;
Yu et al. 2017;
Dieckmann et al. 2021). This tool could be used successfully in genomic subset calculation, construction of Venn diagram or synteny plots (
Supplementary Fig. S1) in terms of robustness, user friendliness and graphical interpretation of the obtained results. The evolutionary relationships among the closely related bacterial genera
Erwinia,
Tatumella and
Pantoea as well as the species within
Pantoea have been recently done by using EDGAR 2.0 software (
Palmer et al. 2017). The genetic basis and phylogenetic distribution of the insecticidal traits of
P. fluorescens against herbivorous insects has also been reported by using the same software (
Flury et al. 2016). EDGAR has resolved the structure of the heterogenous actinobacterial taxon of
Rhodococcus to identify the boundaries for making intrageneric and supra-specific classification (
Sangal et al. 2016). The similar approach was found effective to assess the structure of the genus
Geobacillus (
Aliyu et al. 2016). This tool provides a deep understanding on the whole genome information, especially in the aspects of core genome, dispensable genome and singletons for better interpretation of phylogenetic relationships and species diversity. This approach offers opportunity to develop simple, quick, sensitive and accurate PCR based detection technique compared to other techniques. Herein, we aimed in differentiating the genome content of
Xci from a pool of bacterial population for developing
Xci specific molecular markers in PCR detection (
Fig. 2).
Several molecular methods are used in detection of closely related pathogenic bacterial strains that affecting various crops (
Berg et al. 2005).
Xci specific markers have been yet to develop for diagnosing its’ infection in field grown crucifers. In this regards, 10 singleton genes were randomly selected from 82 retrieved genes for developing
Xci specific markers including one T3E gene for designing 11 PCR based primers. Five primer pairs were produced amplicons specific to
Xci strain (
Fig. 3,
Supplementary Table S1).
T3E protein, a crucial virulence factors in
Xci bacteria, which can be disrupted host cell signaling, cytoskeletal rearrangement, vesicular traffic as well as autophagy in the process of disease development. Bacteria are penetrated into the host cell via a specialized injection apparatus called T3E secretion system and mimic host defense molecules (
Abe et al. 2005). Hoobs
et al. (2016) have been used Genome Search Effector Tool (GenSET) for mining species specific effectors to study the different translocation signals across different
T3E gene families. Currently, researches are concentrated on the prediction and analysis of
T3E gene to understand the bacterial pathogenicity (
Goldberg et al. 2016). T3E genes are believed to confer specificity in
Xci for pathogenicity in ornamental crucifers. Except one
T3E gene, the remaining 81 genes are encoded hypothetical proteins might also play role in species specificity. Thus, out of five well amplified
Xci specific primers, four are situated on genes of hypothetical proteins and could also be used effectively in discriminating
Xci from a mixture of bacterial populations.
Four primer pairs (XCI_1F/R, XCI_3F/R, XCI_5F/R and XCI_6F/R) were able to detect genomic DNA at a low limit of 0.00007 ng/µL (
Fig. 4) which an agreement with the dilution test in detecting
Xcc (
Singh et al. 2014). For understanding the effective detection limit, it is important to diagnose the presence of bacteria in the fields at asymptomic stage. We claim that our primers are very effective and sensitive to detect the
Xci strain even at very low DNA concentration. This is the first report to develop
Xci-specific markers through comparative genomics approach using EDGAR.
CONCLUSION
Bacterial blight disease is an important disease of many crops or plants which is caused by Xci pathogen but there is no marker is available for detection. Therefore, PCR based markers could be used to detect Xci in plant quarantine. However, the efficiency of Xci specific PCR based marker was tested in eighteen bacerial species, but test experiment could be broaden by including more Xanthomonas species upon availability.
COMPLIANCE WITH ETHICAL STANDARDS
The authors have all ethical responsibilities of this manuscript. This research has conducted without any commercial or financial benefits, the authors also declared no conflict of interest and none of the animal and/or human trials were included.
Supplemental Materials
ACKNOWLEDGEMENTS
We thank Joana G. Vicente, University of Warwick, UK for providing different strains with known races of Xcc, Xci and Xcr isolates; also thank to Dr. Pilar Soengas for giving Xcc race 8 strain and the Korean Agriculture Culture Collection (KACC), Korea and the International Collection of Microorganisms from Plants (ICMP), New Zealand for providing respective bacterial strains.
-
AUTHOR CONTRIBUTIONS
I-SN, J-IP and H-TK conceived and designed the study. DMIJ conducted the in silico analysis. MHR prepared and cultured the samples, isolated DNA, performed wet lab validation and wrote the manuscript. UKN extensively revised the manuscript and edited it. HJJ prepared the bacterial inocula and managed the field infected samples. All authors read and approved the final draft of the manuscript.
-
FUNDING
This work was supported by the Center for Horticultural Seed Development (Golden Seed Project No. 213007-05-5-SB510) of the Ministry of Agriculture, Food and Rural Affairs (MAFRA), Republic of Korea.
Fig. 1Circular plot representation of gene clusters present in the X. campestris and X. citri pathovars.
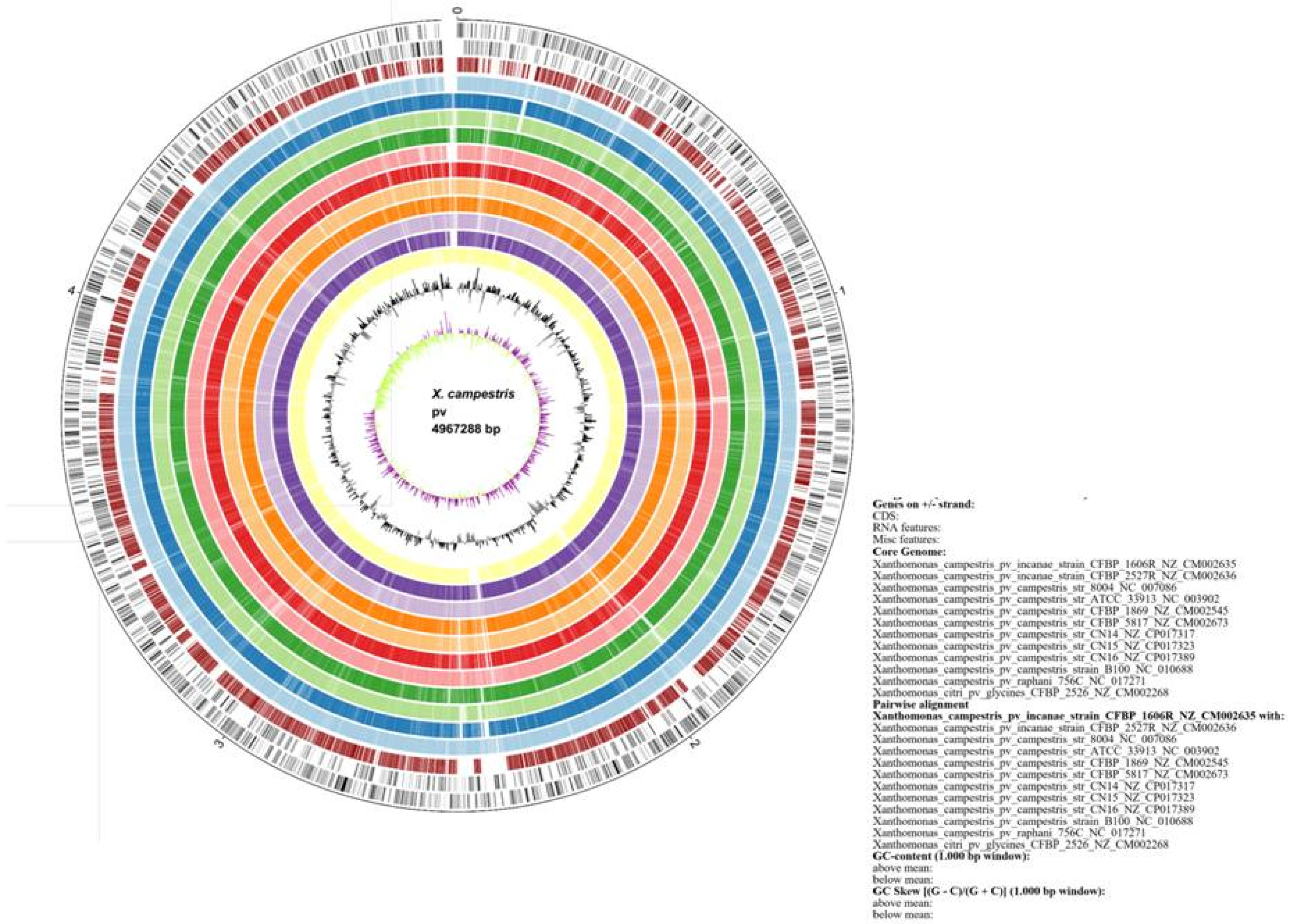
Fig. 2Classification of core (2834), dispensable (1108) and singleton (82) genes. Pie charts showing the distribution of genes in Xanthomonas campestris pv. incanae genome in three segments: core genome (70%) (blue); dispensable genome (28%) (red) and singletons (2%) genes (green).
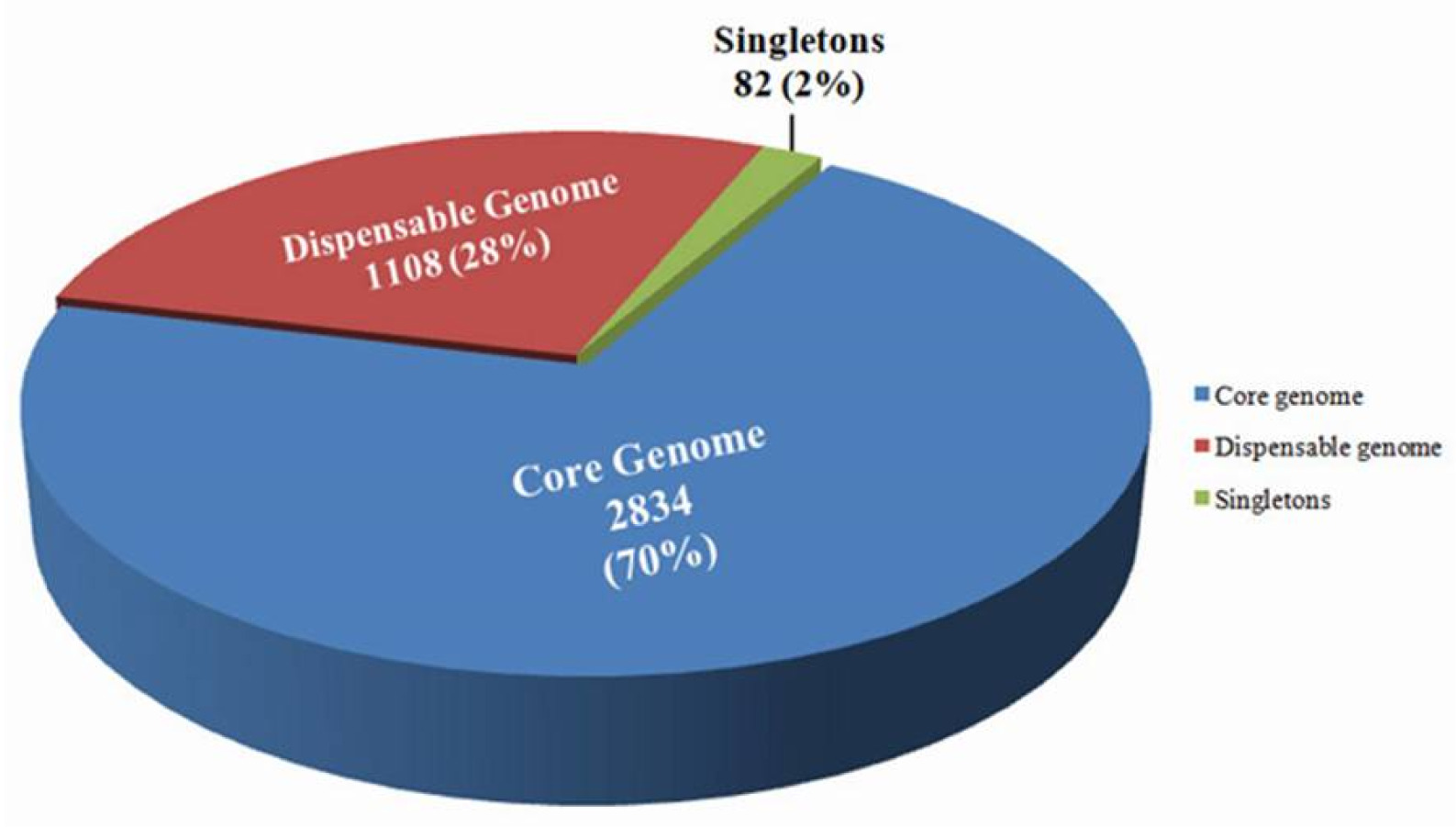
Fig. 3Specific detection of Xanthomonas campestris pv. incanae strain from other closely related Xanthomonas campestris pathovars such as pv. campestris, pv. raphani and other bacterial strains using the Xci specific developed primers (XCI_1F/R, XCI_2F/R, XCI_3F/R, XCI_5F/R and XCI_6F/R) in PCR based assay. 100 bp DNA ladder; Lane 1: Xci (HRI-W-6377); Lane 2-9: Xcc (HRI-W-3811, HRI-W-3849A, HRI-W-5212, HRI-W-1279A, HRI-W-3880, HRI-W-6181, HRI-W-8450A and MBG145.3); Lane 10: Xcr (HRI-W-8305) Lane 11: Xev (KACC11153); Lane 12: Xag (KACC10491); Lane 13: Xcz (KACC17126); Lane 14: Xc (KACC10490); Lane15: Pseudomonas syringae pv. maculicola (ICMP13051); Lane16: Erwinia carotovora subsp. carotovora (ICMP12464); Lane 17: Plasmodiophora brassicae; Lane 18: Didymella bryoniae (NIHHS1326) and Lane 19: Negative control (distilled water).
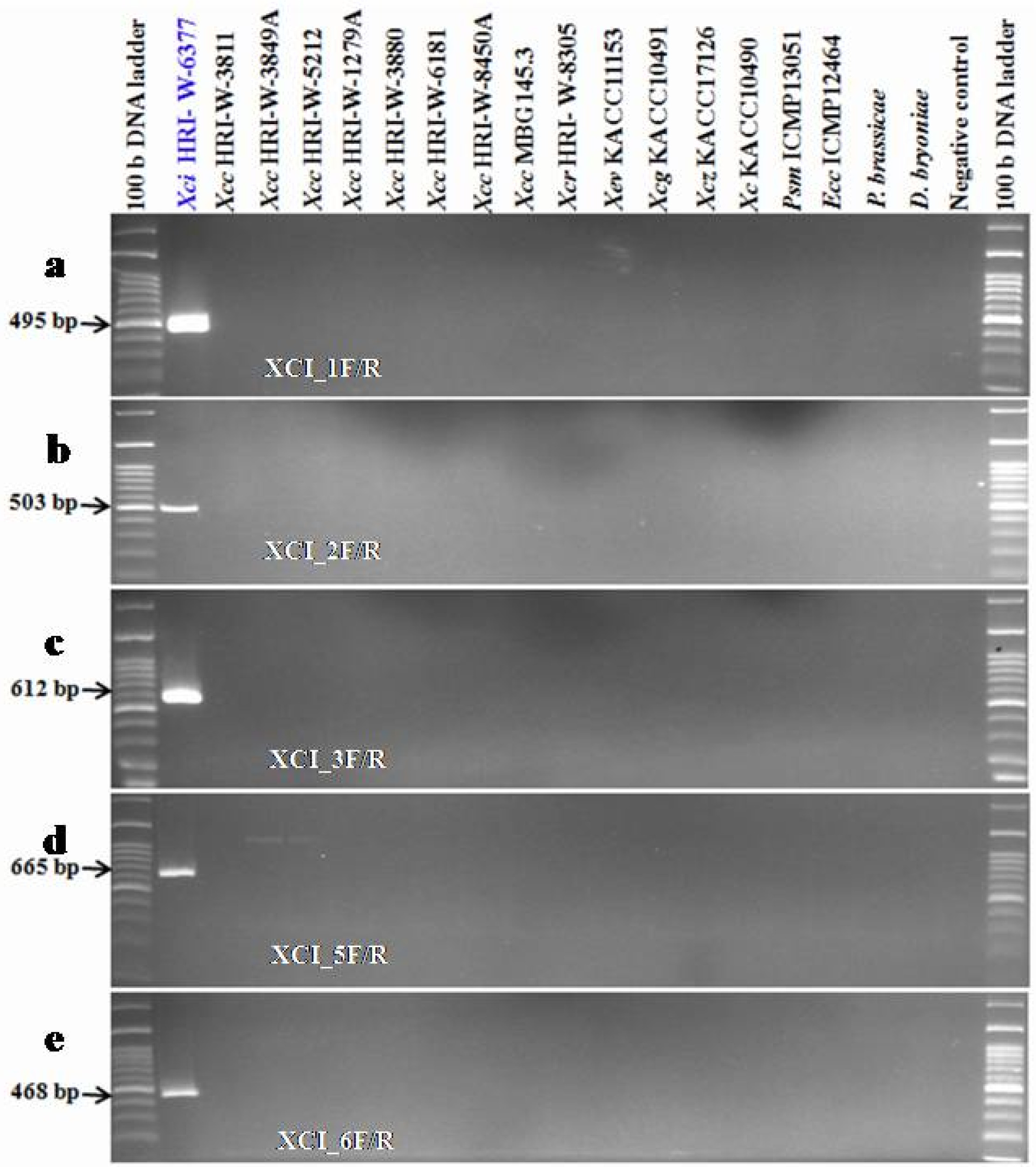
Fig. 4PCR detection limits regards to the genomic DNA concentration of Xci strain (HRI-W-6377) using developed markers (XCI_1F/R, XCI_2F/R, XCI_3F/R, XCI_5F/R and XCI_6F/R) in PCR based assay.
Table 1
Xanthomonas chromosomes description including genome size, total gene and protein of each individual strain with NCBI taxonomic ID.
Table 1
|
Strain |
Xcc B100 |
Xcc ATCC33913 |
Xcc CFBP5817 |
Xcc 8004 |
Xcc CN17 |
Xcr 756C |
Xci CFBP1606R |
Xci CFBP2527R |
Xev LMG930 |
Xcg CFBP2526 |
Xad LMG695 |
Xac Xac 29 |
|
Size (Mbp) |
5.07 |
5.07 |
4.97 |
5.14 |
4.99 |
4.94 |
4.96 |
4.92 |
5.07 |
5.363 |
4.99 |
4.60 |
|
Gene |
4,542 |
4,240 |
4,447 |
4,584 |
4,398 |
4,372 |
4,423 |
4,380 |
4,977 |
4,856 |
4,592 |
4,641 |
|
Protein |
4,236 |
4,179 |
4,122 |
4,297 |
4,086 |
4,055 |
4,071 |
4,032 |
4,505 |
4,461 |
3,919 |
4,237 |
|
NCBI ID |
NC_010688 |
NC_003902 |
NZ_CM002673 |
NC_007086 |
CP011946.1 |
NC_017271 |
NZ_CM002635 |
NZ_CM002636 |
NZ_CP018467 |
NZ_CM002268 |
NZ_CP014347 |
NC_020800 |
Table 2List of primer sequences with expected product size and annealing temperatures for specific amplification of Xanthomonas campestris pv. incanae.
Table 2
|
Primers |
Forward (5’-3’) |
Reverse (5’-3’) |
Expected size (bp) |
Annealing temperature (°C) |
|
XCI_1F/R |
ATGTCGATCCAGCTCAGGTG |
TCAGGTTGCAGGAGGGCTTGG |
495 |
64 |
|
XCI_2F/R |
CCTTCAGGTCATGCACCATTG |
ATCCGCATTCTGCCCTCCCCCCT |
503 |
65 |
|
XCI_3F/R |
GCGATGGATACAGTGACTGGAAG |
CAACTGCAAGGCCGTATCGTA |
612 |
62 |
|
XCI_4F/R |
CAGGCGGCCGATGCGAAGG |
TGATGTTGAAGAGTGTGCGGAC |
604 |
63 |
|
XCI_5F/R |
CATTCTCGCGGAGCGGCGTCTGC |
AGCTGCTGAGTTGCCGCGCTCT |
665 |
68 |
|
XCI_6F/R |
TCTGGCTCCCACTATGTGC |
CGGGATGGATGAGGCTGTCC |
468 |
60 |
|
XCI_7F/R |
GTTCGGACTGGCGCTCTGGC |
TCTTCATCATCACTAACCCA |
303 |
60 |
|
XCI_8F/R |
GATCATGGTGTCGCCCATG |
TTAAGCCACTTTGAGCTGCACC |
324 |
60 |
|
XCI_9F/R |
GTGACCCTGCGATACATCGG |
GGTGCTGCCAAGGCCGGAGC |
769 |
65 |
|
XCI_10F/R |
GGTACTACTGGATGCCAACC |
TCAGGCCTTCATAAGATTTTG |
397 |
57 |
|
XCI_11F/R |
ATGGATGCCGCTGCCGGTCG |
GCGGGTAGGTGAGGCCGTCGA |
595 |
65 |
Table 3Molecular marker information for specific detection of Xanthomonas campestris pv. incanae.
Table 3
|
Primers |
Gene name |
Start |
End |
Length (bp) |
Direction |
Protein description |
|
XCI_1F/R |
XCICFBP2527_RS16430
|
3641561 |
3641067 |
495 |
- |
Hypothetical protein |
|
XCI_2F/R |
XCICFBP2527_RS13555
|
2955263 |
2956369 |
1,107 |
+ |
Hypothetical protein |
|
XCI_3F/R |
XCICFBP2527_RS21720
|
4861522 |
4862766 |
1,245 |
+ |
Hypothetical protein |
|
XCI_5F/R |
XCICFBP2527_RS03935
|
886319 |
884361 |
1,959 |
- |
Hypothetical protein |
|
XCI_6F/R |
XCICFBP2527_RS21980
|
196812 |
196171 |
642 |
- |
Type III effector |
References
- Abby S, Daubin V. 2007. Comparative genomics and the evolution of prokaryotes. Trends in Microbiology. 15: 135-141.
- Abe I, Utsumi Y, Oguro S, Morita H, Sano Y, Noguchi H. 2005. A plant type III polyketide synthase that produces pentaketide chromone. J. Am. Chem. Soc.. 127: 1362-1363.
- Afrin KS, Rahim MA, Rubel MH, Natarajan S, Song JY, Kim HT, et al. 2018. Development of race-specific molecular marker for Xanthomonas campestris pv. campestris race 3, the causal agent of black rot of crucifers. Can. J. Plant Sci.. 98: 1119-1125.
- Aliyu H, Lebre P, Blom J, Cowan D, De Maayer P. 2016. Phylogenomic re-assessment of the thermophilic genus Geobacillus. Syst. Appl, Microbiol.. 39: 527-533.
- Berg T, Tesoriero L, Hailstones D. 2005. PCR based detection of Xanthomonas campestris pathovars in Brassica seed. Plant Pathol.. 54: 416-427.
- Blom J, Albaum SP, Doppmeier D, Pühler A, Vorhölter FJ, Zakrzewski M, et al. 2009. EDGAR: a software framework for the comparative analysis of prokaryotic genomes. BMC Bioinformatics. 10: 1-14.
- Blom J, Kreis J, Spänig S, Juhre T, Bertelli C, Ernst C, et al. 2016. EDGAR 2.0: an enhanced software platform for comparative gene content analyses. Nucleic Acids Rese.. 44: W22-W28.
- Cruz J, Tenreiro R, Cruz L. 2017. Assessment of diversity of Xanthomonas campestris pathovars affecting cruciferous plants in Portugal and disclosure of two novel X. campestris pv. campestris races. J. Plant Pathol.. 99: 403-414.
- Cruz J, Tenreiro R, Cruz L. 2018. Inference of the phylogenetic diversity and population structure of Xanthomonas campestris affecting Brassicaceae using a multilocus sequence typing based approach. Plant Pathol.. 67: 948-956.
- Dieckmann MA, Beyvers S, Nkouamedjo-Fankep RC, Hanel PHG, Jelonek L, Blom J, et al. 2021. EDGAR3. 0: comparative genomics and phylogenomics on a scalable infrastructure. Nucleic Acids Res.
- Ecker R, Zutra D, Barzilay A, Osherenko E, Rav-David D. 1995. Sources of resistance to bacterial blight of stock (Matthiola incana R. Br.). Genet. Resour. Crop Evol.. 42: 371-372.
- Fargier E, Fischer-Le Saux M, Manceau C. 2011. A multilocus sequence analysis of Xanthomonas campestris reveals a complex structure within crucifer-attacking pathovars of this species. Syst. Appl. Microbiol.. 34: 156-165.
- Fargier E, Manceau C. 2007. Pathogenicity assays restrict the species Xanthomonas campestris into three pathovars and reveal nine races within X. campestris pv. campestris. Plant Pathol.. 56: 805-818.
- Flury P, Aellen N, Ruffner B, Péchy-Tarr M, Fataar S, Metla Z, et al. 2016. Insect pathogenicity in plant-beneficial pseudomonads: phylogenetic distribution and compa-rative genomics. ISME J.. 10: 2527-2542.
- Garrigues C, Johansen E, Crittenden R. 2013. Pangenomics-an avenue to improved industrial starter cultures and probiotics. Curr. Opin. Biotechnol.. 24: 187-191.
- Goldberg T, Rost B, Bromberg Y. 2016. Computational prediction shines light on type III secretion origins. Sci. Rep.. 6: 1-10.
- Hobbs CK, Porter VL, Stow ML, Siame BA, Tsang HH, Leung KY. 2016. Computational approach to predict species-specific type III secretion system (T3SS) effectors using single and multiple genomes. BMC Genomics. 17: 1-11.
- Kałużna M, Pulawska J, Waleron M, Sobiczewski P. 2014. The genetic characterization of Xanthomonas arboricola pv. juglandis, the causal agent of walnut blight in Poland. Plant Pathol.. 63: 1404-1416.
- Kendeick J, Baker KF. 1942. Bacterial blight of garden Stocks and its control by hot-water seed treatment. Bulletin of the California Agricultural Experiment Station..
- Laala S, Cesbron S, Kerkoud M, Valentini F, Bouznad Z, Jacques MA, et al. 2021. Characterization of Xanthomonas campestris pv. campestris in Algeria. Phytopathologia Mediterr.. 60: 51-62.
- Minardi P, Mazzucchi U, Parrini C. 1988. Epidemie di avvizzimento batterico della violaciocca (Matthiola incana R Br.) da Xanthomonas campestris pv. incanae in Toscana. Inf. Fitopatologico..
- Mira A, Martín-Cuadrado AB, D'Auria G, Rodríguez-Valera F. 2010. The bacterial pan-genome: a new paradigm in microbiology. Int. Microbiol.. 13: 45-57.
- Palmer M, Steenkamp ET, Coetzee MP, Chan WY, Van Zyl E, De Maayer P, et al. 2017. Phylogenomic resolution of the bacterial genus Pantoea and its relationship with Erwinia and Tatumella. Antonie Van Leeuwenhoek. 110: 1287-1309.
- Parkinson N, Cowie C, Heeney J, Stead D. 2009. Phylogenetic structure of Xanthomonas determined by comparison of gyrB sequences. Int. Jo. Syst. Evol. Microbiol.. 59: 264-274.
- Pirone PP. 1978. 'Diseases and pests of ornamental plants' (John Wiley & Sons).
- Rahimian H, Okhovatian H. 1989. Bacterial blight of stock in Mazadaran. Iran. J. Plant Pathol. (Iran).
- Reinhardt JA, Baltrus DA, Nishimura MT, Jeck WR, Jones CD, Dangl JL. 2009. De novo assembly using low-coverage short read sequence data from the rice pathogen Pseudomonas syringae pv. oryzae. Genome Res.. 19: 294-305.
- Reno ML, Held NL, Fields CJ, Burke PV, Whitaker RJ. 2009. Biogeography of the Sulfolobus islandicus pan-genome. Proc. Natl. Acad. Sci. U.S.A.. 106: 8605-8610.
- Rey MW, Ramaiya , Nelson BA, Brody-Karpin SD, Zaretsky EJ, Tang M, et al. 2004. Complete genome sequence of the industrial bacterium Bacillus liche-niformis and comparisons with closely related Bacillus species. Genome Biol.. 5: 1-12.
- Roux B, Bolot S, Guy E, Denancé N, Lautier M, Jardinaud MF, et al. 2015. Genomics and transcriptomics of Xanthomonas campestris species challenge the concept of core type III effectome. BMC genomics. 16: 1-19.
- Rubel MH, Hossain MR, Nath UK, Natarajan S, Lee JH, Jung HJ, et al. 2019. Development of a PCR test for detection of Xanthomonas campestris pv. raphani. Australas. Plant Pathol.. 48: 179-182.
- Rubel MH, Natarajan S, Hossain MR, Nath UK, Afrin KS, Lee JH, et al. 2019. Pathovar specific molecular detection of Xanthomonas campestris pv. campestris, the causal agent of black rot disease in cabbage. Can. J. Plant Pathol.. 41: 318-328.
- Rubel MH, Robin AHK, Natarajan S, Vicente JG, Kim HT, Park JI, et al. 2017.
- Sangal V, Goodfellow M, Jones AL, Schwalbe EC, Blom J, Hoskisson PA, et al. 2016.
- Singh D, Raghavendra B, Rathaur PS, Singh H, Raghuwanshi R, Singh R. 2014. Detection of black rot disease causing pathogen Xanthomonas campestris pv. campestris by bio-PCR from seeds and plant parts of cole crops. Seed Sci. Technol.. 42: 36-46.
- Sivashankari S, Shanmughavel P. 2007. Comparative genomics-a perspective. Bioinformation. 1: 376
- Tettelin H, Masignani V, Cieslewicz MJ, Donati C, Medini D, Ward NL, et al. 2005. Genome analysis of multiple pathogenic isolates of Streptococcus agalactiae: implications for the microbial "pan-genome". Proc. Natl. Acad. Sci. U.S.A.. 102: 13950-13955.
- Vicente JG, Conway J, Roberts S, Taylor J. 2001. Identification and origin of Xanthomonas campestris pv. campestris races and related pathovars. Phytopathology. 91: 492-499.
- Vicente JG, Everett B, Roberts S. 2006. Identification of isolates that cause a leaf spot disease of brassicas as Xanthomonas campestris pv. raphani and pathogenic and genetic comparison with related pathovars. Phyto-pathology 96. 735-745.:
- Vicente JG, Holub EB. 2013. Xanthomonas campestris pv. campestris (cause of black rot of crucifers) in the genomic era is still a worldwide threat to brassica crops. Mol. Plant pathol.. 14: 2-18.
- Vicente JG, Taylor J, Sharpe A, Parkin I, Lydiate D, King G. 2002. Inheritance of race-specific resistance to Xanthomonas campestris pv. campestris in Brassica genomes. Phytopathology. 92: 1134-1141.
- Williams PH. 1980. Black rot: A continuing threat to world crucifers. Plant Dis.. 64: 736-742.
- Yu J, Blom J, Glaeser S, Jaenicke S, Juhre T, Rupp O, et al. 2017. A review of bioinformatics platforms for comparative genomics. Recent developments of the EDGAR 2.0 platform and its utility for taxonomic and phylogenetic studies. J. Biotechnol.. 261: 2-9.
- Zhang W, Jayarao BM, Knabel SJ. 2004. Multi-viru-lence-locus sequence typing of Listeria monocytogenes. Appl. Enviro. Microbiol.. 70: 913-920.