Abstract
Cotton fiber development is a complex process regulated by multiple networks that involve various transcription factors (TFs). Recent studies (2020-2025) have elucidated the roles of TFs through silencing and overexpression experiments in cotton plants. We conducted a literature search in NCBI using the keywords ‘transcription factor, fiber elongation, cotton plant’ to identify relevant studies. This review focuses on studies published between 2020 and 2025 that were selected for review. We discussed various TFs and molecular mechanisms leading to altered fiber length. The research works included various analyses of transcription factors, genes, proteins, and metabolites as well as fiber quality results. The reviewed data highlight the complexity of fiber development networks, which involve extensive crosstalk among regulatory pathways. Despite the complexity, many novel TFs, modules, and tandems that participate in fiber elongation have been found. Their positive or negative regulatory roles in fiber formation stages promote a more profound understanding of fiber initiation and elongation.
-
Key words: Transcription factors, Cotton fiber, Fiber elongation, Fiber development, Gossypium hirsutum
Introduction
Cotton fiber quality is the primary focus of global cotton research. It aligns the discipline, such as stress-resistance, leading to improved plant growth and development (
Abdurakhmonov et al. 2016). RNA interference (RNAi) gained significant attention in cotton fiber genetics a decade ago (
Abdurakhmonov et al. 2014). Advances in scientific techniques and the integration of various disciplines studying genes, proteins, transcription factors, metabolites, and lipids have accelerated research and facilitated the interpretations leading to clarity of the results.
A review paper on trichomes in plants, published in 2021, reported more than thirty genes involved in trichome development as positive or negative regulators (
Wang et al. 2021b). In this context, special attention has been paid to genes and transcription factors (TFs) involved in cotton fiber initiation, elongation, and development (
Han et al. 2022). In the last five years, more than 50 papers devoted to studying fiber elongation-related TFs have been published in the NCBI database. Recent approaches pay attention to discussing fiber elongation-related features together with other points, including plant growth and development, secondary plant metabolism, including phytohormones and lipids, and other points (
Wen et al. 2023).
Diverse mechanisms and networks are involved in fiber elongation. Recent papers are highlighting the significance of TFs together with genes. AtZAT10/STZ1 is one of the TFs belonging to C2H2-type zinc finger proteins involved in fiber development. Its expression in cotton led to enhanced agronomic traits of the plant, including drought resistance as well as enhanced fiber yield. Transcriptome analysis showed the significance of TFs aligned with kinases, aquaporins, and heat-shock proteins for the above-mentioned enhanced features (
Qin et al. 2024).
GhFL10 was found as a functional gene involved in fiber elongation. Its role was linked with NF-YA transcription factors to negatively regulate fiber elongation (
Zhang et al. 2022). These and other scientific data explain the complexity of fiber formation, which can be interpreted by the integration of various approaches.
Integrating data from multiple approaches can improve our understanding on fiber elongation and development. In this paper, we will discuss cotton fiber elongation-related features obtained in the last decade since 2020.
Day-dependent Expressions
Plant and cultivation (subtitle)
The complexity of fiber initiation and elongation has been demonstrated in recent papers. The analyses of differentially expressed levels of genes, TFs, and miRNAs and comparative studies using various techniques can facilitate the interpretation of results (
Arora et al. 2021). In this manner, day-dependent expressions of TFs and genes play pivotal roles in fiber formation and development. In one work, GhMYB3 and ARF5 were reported to regulate the expression of
GhROT3-1 and
GhDET2-1 genes, respectively; their roles in fiber initiation and elongation were highlighted. The predominant expression of GhROT3-1 was in 10 and 15 days post anthesis (DPA), and high expression of GhDET2-1 was in 5 DPA. Thus, the significance of genes related to brassinosteroid biosynthesis was established (
Cui et al. 2024). In another work, 1467 TFs belonging to MYB, bHLH, TCP, and bZIP were identified, and 148 of them were upregulated in fibers. The authors found GhMYB25 and GhMYB25-like TFs as the main factors during cell differentiation and early fiber elongation. GhMYB46 was highly expressed in 20 DPA, which was linked with the secondary cell wall deposition. GhMYB109 was found in the differentiation and elongation, and was claimed to contribute to fiber length reduction. The authors emphasized the need for further analyses to better understand of the regulatory roles of TFs in fiber elongation and development (
Yang et al. 2021). Another group of researchers identified 5131 differentially expressed genes (DEGs) in a comparative study. Among them, 290 belonged to TFs of MYB, bHLH, ERF, and C2H2. Studying the expression of seven of them showed day-dependent differences in their expression levels. Some of them showed similar levels for 20 days, whereas others reached a peak in 25 DPA and then declined in 30 DPA. These comparisons ease the understanding of the roles of various TFs in one of three stages of fiber formation: fiber initiation, fiber elongation, or fiber development (
Li et al. 2024a).
Several works showed day-dependent expressions of various TFsand genes involved in fiber development. In one work, NAC1 TF activaing GhGLU18 glucanase, known to cleave β-1,3-glucan bonds, was explored. The overexpression of GhGLU18 was found to affect secondary cell wall formation as a contributing factor to fiber elongation. Several isoforms of Glu were expressed day-dependently (
Fang et al. 2023). GhZFP8 is another TF involved in fiber elongation, which was found to be expressed in 3-30 DPA. Its overexpression led to a higher number of trichomes. Its interference caused smaller bolls and shorter fiber length. Thus, the involvement of GhZFP8 in fiber development was established (
Liu et al. 2024b). Future studies should integrate day-dependent expression levels of TFs and genes involved in fiber elongation.
Cotton fiber elongation is the second and primary stage of cotton fiber development, occurring between fiber initiation and secondary wall biosynthesis. The first 2-3 DPA are the initiation, followed by elongation for the next three-four weeks. Secondary wall biosynthesis is observed in the next 20-25 days and ends with the maturation of fiber (
Jareczek et al. 2023;
Fig. 1).
Fiber Elongation TFs
Several TFs have been identified involved in fiber elongation so far. Their expression levels were linked with the regulations of genes, proteins, and metabolites participating in this process. In this section, we discuss those TFs found in cotton plant playing roles in fiber elongation and development.
MYB TFs
MYB TFs are among the most extensively studied TFs families involved in various biological functions, including fiber development (
Zhai et al. 2023). More than 400
MYB genes are expected to be expressed during fiber development in cotton plants (
Huang et al. 2021). MYB TFs are important regulators of cell elongation.
GhMYB4, one of the members of these genes, was reported to suppress two genes and reduce fiber cell elongation. Its knockdown in cotton increased the fiber length, and the overexpression in model plants lowered the levels of root length and plant height. Thus, it was established to be a part of cell elongation (
Duan et al. 2024). The involvement of GhMYB30D04 in cotton fiber development was explained by linking it with the accumulation of phosphatidylinositol phosphates (PIPs). Its silencing lowered the expression of GhLTPG1 transcripts and the accumulation of PIP, which were accompanied by lower fiber yield and length. Its overexpression enriched PIPs, elongated the cotton fiber, and upregulated GhLTPG1. The authors concluded that the enhancement of GhLTPG1 by the interaction between PIP and GhMYB30D04 leads to cotton fiber elongation (
Song et al. 2024). These results explain the significance of GhMYB30D04 TF in cotton fiber elongation and demonstrate the involvement of GhLTPG1 transcript in this framework.
GhMYB201 is another R2R3-MYB TF, the knockout of which reduced the fiber length in cotton plants. The shorter fiber length in the GhMYB201 knockout plants was linked with the reduced levels of
GhRDLs (cell wall loosening) and
GhKCSs (β-ketoacyl-CoA synthase) genes. Thus, GhMYB201 TF was suggested to be responsible for fiber elongation and cell wall loosening by involving
GhRDLs and
GHKCSs genes (
Suo et al. 2024). GhMYB44 was reported as another factor, the overexpression of which reduced the fiber length. Correlational analysis proved the relationship between the gene expression level and N6-methyladenosine (m6 A). The methylation of m6 A altered the levels of genes related to fiber elongation. Silencing GhMYB44 resulted in elongated fiber (
Xing et al. 2023).
GhMML3_D12, belonging to MYBMIXTA-like (MML) TFs, was found as another protein that regulates fiber elongation. Its overexpression in cotton developed fuzz fiber, and its duplication caused the formation of a fuzzless phenotype. The integration of comparative transcriptome analyses and RNA sequencing determined the central role of GhMML3 in network regulating fiber early-stage elongation. GhMML3_D12 was suggested to promote GhHD-1 and GhMYB25, which are involved in fiber initiation (
Chen et al. 2025). Comparing the expression of TFs in various cotton types is another approach to identifying the role of various TFs or genes. For example, MYB5 and WER TFs were found to cause fiber elongation in
G. stocksii. WER and MYB5 were found to be upregulated in
G. stocksii, whereas downregulated in
G. arboreum. The authors suggested the involvement of these two TFs in
G. stocksii fiber elongation (
Sun et al. 2022).
TCP TFs represent another class involved in functions, including fiber development (
Zhai et al. 2023). In one work carried out on
G. raimondii, 37 non-redundant TCP proteins were found, and their involvement in fiber development was studied. Among the factors,
GrTCP11 was suggested to be important for fiber development as its function was explained to suppress the biosynthesis of JA.
GrTCP11 was also suggested to influence root hair cell elongation in the same way (
Hao et al. 2021). GhTCP4 is another TF responsible for cell wall expansion and fiber elongation. The functions of GhTCP4 and GhHOX3 were antagonistic in cell elongation, and the GhHOX3-GhTCP4 complex was concluded as a general mechanism for designing fiber traits. Its overexpression led to the acceleration of secondary cell wall pathways. The contents of crystalline cellulose in GhTCP4-overexpressed lines were greater than in wild-type plants. The authors concluded the contribution of the miR319-GhTCP4 module in the regulation of cotton fiber development (
Cao et al. 2020).
WRKY TFs also play roles in cotton fiber initiation and elongation.
GhWRKY16 was reported as one of these genes playing a role in these processes. Its silencing reduced the fiber length compared to wild-type plants. Its phosphorylation was concluded to bind to several promoters (GhHOX3, GhMYB109, GhCesA6D-D11, and GhMYB25) and activate them in early fiber development. The authors concluded the fiber elongation-promoting role of GhWRKY16 by regulating GhHOX3 and GhMYB109 (
Wang et al. 2021a). In one work, the day-dependent genome-wide mapping of DNase I hypersensitive sites involved in fiber development was carried out. The authors found the involvement of WRKY46 in fiber development. The TF regulating the lignin content significantly reduced the primary xylem area in the systemic stem of cotton plants (
Bao et al. 2023). The significant roles of WRKY TFs in cell elongation were also shown in root elongation of the model plant by studying 25
WRKY genes (
Bao et al. 2024).
GhHDZ76, a member of the HD-Zip family, is known to be expressed during the early days of fiber development. Its knockout in cotton plants resulted in shorter fibers compared to wild-type plants. The transcript levels of fiber initiation-related genes were lower in the GhHDZ76 knockout plants. GhHDZ76 has been established to regulate cotton fiber initiation and elongation (
Wu et al. 2024). Phosphatidic acid was reported as one of the molecules contributing to fiber elongation. In one work, it was found to interact with HD-ZIP TF GhHOX4. The GhHOX4 overexpression in cotton enhanced the fiber length, and the reverse effects were found with its silencing. Basic amino acids such as arginine and lysine, located in the START domain of GhHOX4, were claimed to bind phosphatidic acid, which leads to changes in their localizations, causing alterations from fiber elongation to cell wall thickening (
Wang et al. 2024b).
Basic helix-loop-helix (bHLH) TFs are involved in multiple regulatory networks, including those governing fiber development. The overexpression of GhFP1 protein, regulated by bHLH, was found to lead to fiber elongation. Brassinosteroid-related genes were up- and downregulated in GhFP1-overexpressed and -silenced plants, respectively. Thus, the authors showed the activation of brassinosteroid-related genes by GhFP1, suggesting its positive regulatory role on other genes (
Liu et al. 2020). bHLH/HLH TFs are among the significant key factors in cell elongation. In one work, the involvement of bHLH/HLH in fiber elongation was found, and the authors concluded that the regulation of fiber elongation by many TFs, including those of brassinosteroid synthesis. GhACE1 TF was found to promote fiber elongation via the activation of
GhPIP2;7 and
GhEXP8 genes (
Lu et al. 2022). Pectate lyases have also been studied for their role in fiber development. qRT-PCR analysis showed the predominant expression of the
GhPEL76 gene in cotton fiber. The involvement of the gene in fiber elongation was established in cotton fiber and long fiber cultivars. The gene promoted β-glucuronidase activity in various organs of Arabidopsis. Its silencing in cotton resulted in reduced fiber length. Seemingly, GhbHLH13 TF promoted
GhPEL76 expression, which can be one of the mechanisms of bHLH TFs leading to longer cotton fibers (
Sun et al. 2020).
A recent review by
Jan et al. (2022) describes the involvement of various phytohormones and TFs in cotton fiber development. Various pathways involving brassinosteroids, gibberellic acid, ethylene, fatty acids, indole acetic acid, and others are included in the fiber development (
Jan et al. 2022;
Fig. 2).
A brassinosteroid transcriptional regulatory network is involved in fiber elongation. In another work, GhBES1 enhanced fiber elongation as an activator of brassinosteroid signaling. In GhBES1-overexpressed plants, hemicellulose accumulation was increased (
Zhu et al. 2023). GhBES1.4 is another TF of brassinosteroid signaling involved in fiber elongation. It was found to modulate long chain fatty acids biosynthesis. Silencing GhBES1.4 caused significantly shorter fiber length, and its overexpression resulted in longer fiber length. These results explain the significance of GhBES TF and long chain fatty acids in the fiber elongation (
Yang et al. 2023). The overexpression of GhBES1.4 in cotton led to fiber elongation. Seven genes regulated by GhBES1.4 were found to participate in fiber elongation. The integration of various approaches data enabled the authors to conclude the involvement of the gene in cell development and cell processes. The authors concluded that the cross-talk of brassinosteroids and other hormones in the regulation of fiber development (
Liu et al. 2023). Gh_DNF_YB19, a nuclear TF, was reported to participate in the regulation of fiber development by interacting with GhSK13, a component of the brassinosteroid signaling pathway. The results highlighted the significance of phosphorylation/dephosphorylation in the brassinosteroid signaling transduction pathway (
Wen et al. 2023).
The crosstalk between Gibberellin (GA) and strigolactones (SLs) is suggested to be involved in fiber development. Growth-regulating factor 4 (GhGRF4) is one of the key molecules activating the biosynthesis of SL. Knocking out by GA-induced GhGRF4 altered SL-mediated fiber cell elongation. Thus, it was established to affect fiber cell wall length and thickness (
Tian et al. 2022). Synergistic effects were implied between TFs and phytohormones that contribute to the regulation of fiber elongation (
Zhai et al. 2023).
Ethylene response TFs are known to participate in plant growth and development. The overexpression of GhERF109, a member of these factors, led to reduced cotton plant height. Its silencing altered the levels of 4123 genes, among which various genes of the networks, including auxin response, cell expansion, cell cycle regulation, and others, were increased. Thus, GhERF109 was found to be one of the key molecules that can significantly improve plant growth (
Shi et al. 2025).
Auxin response factor (ARF) genes are another class of molecules playing significant roles in fiber development.
GhARF2b was found specifically higher in developing fibers of
G. hirsutum among 9
GhARF2 genes. Its overexpression reduced fiber elongation, and silencing promoted longer fibers. The alterations in the level of
GhHOX3 were concluded as an intermediate step (
Zhang et al. 2021). In another work, the significance of MYB TF was linked with ribosome biosynthesis and phytohormone regulation. The expression levels of four isoforms - MYB4, MYB66, MYB96, and MYB106 were upregulated in 2DPA and were consistent with
ARF19a,
ARF12b, and
BSL1 participating in auxin and brassinosteroid signaling. Other isoforms, including MYB6, MYB16, MYB17, MYB73 were downregulated in 2DPA. Approximately 50 genes related to phytohormones were downregulated. These results explain the complexity of understanding the regulation of fiber initiation and elongation (
Ando et al. 2021).
ARFs have also been determined to be involved in fiber development. Two ARFs, such as GhARF7-1 and GhARF19-1, were found to show a positive regulatory effect on fiber cell initiation and elongation. These two TFs were determined to interact with each other and form heterodimers. Both of them revealed binding ability to GhXTH9, which was selected as their downstream targets. These results explain the significance of binding properties in fiber elongation and development (
Jin et al. 2024). TF-binding motifs play crucial roles in fiber development, and their analyses can be a significant point at early stages of the work (
Chen et al. 2023).
Involvement of Lipids in Fiber Elongation
Lipids are among the compounds playing roles in fiber cell elongation. Various lipids have been identified in this process, with mechanisms including TFs. GhZFP8 and GhBLH1 are two TFs that affect fiber elongation by promoting biosynthesis of very long-chain fatty acids (VLCFAs). These two TFs and GhSLR1 which interfere with them were also identified in the regulation of trichome development. These findings suggest that elucidating the mechanisms and outcomes of various crosstalks will clarify the overall network of fiber elongation and development (
He et al. 2024). Brassinazole-resistant (BZR) TF, playing a role in brassinosteroid signaling, has also been found to be an essential element in fiber elongation. Silencing GhBZR3, a member of BZR TF, in cotton resulted in longer plant height and fiber length. VLCFA quantity was significantly higher in GhBZR3-silenced plants, which was explained by GhBZR3 binding sites of the GhKCS13 promoter gene involved in VLCFA biosynthesis (
Shi et al. 2022). In another work, lipid transfer protein4 (GhLTP4) was shown to promote fiber elongation. Its overexpression enhanced the fiber length by increasing ceramides quantity, and conversely, downregulation reduced that parameter. Significantly higher quantities of transcripts of IAA-responsive genes were determined in the GhLTP4-overexpressing plants. The treatment with exogenous ceramides, composed of sphingosine and fatty acids, led to longer fiber length. These results explain the essential roles of fatty acids and the TFs involved in their metabolism for cotton fiber elongation (
Duan et al. 2023).
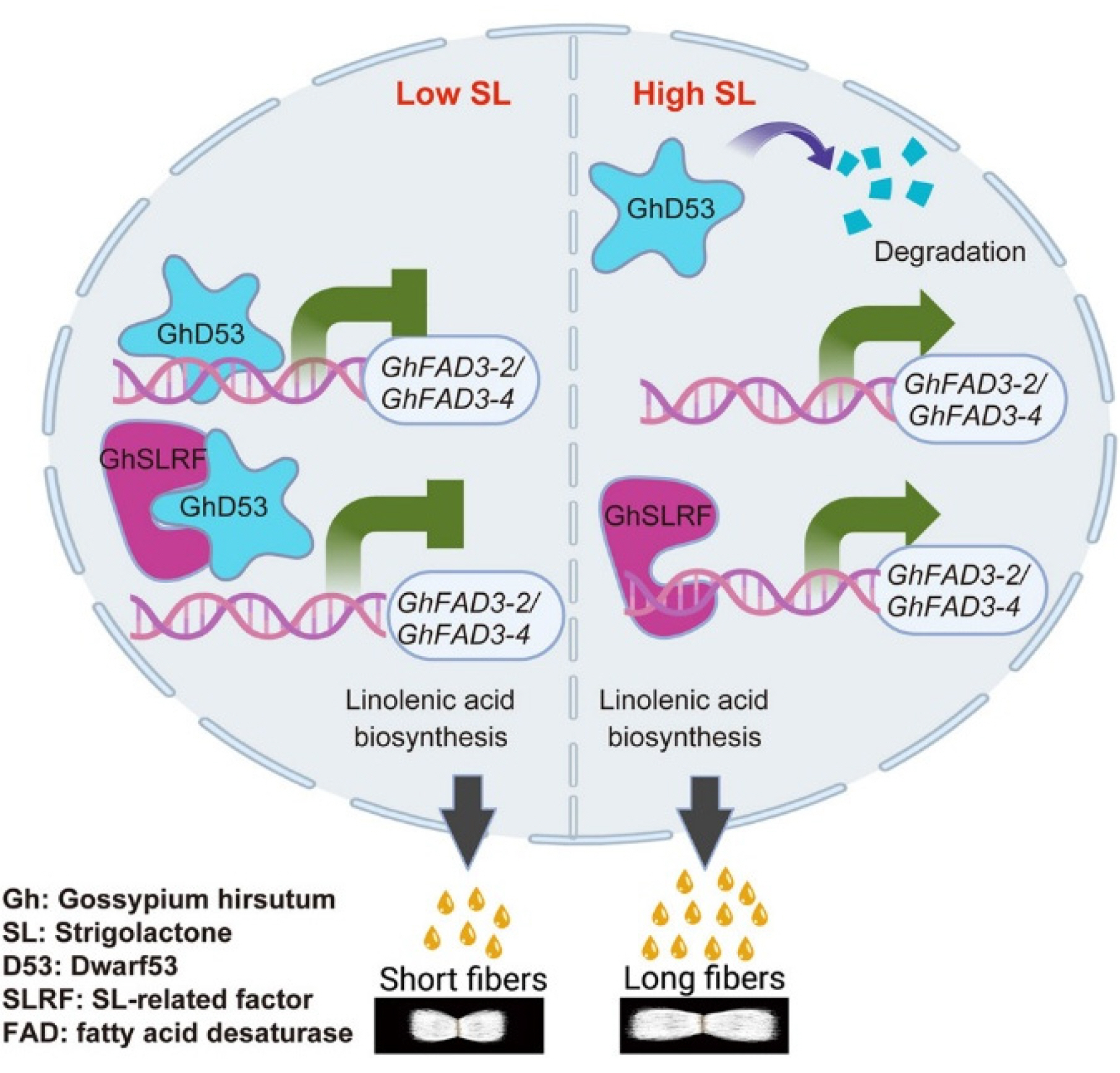
In one unique work, the effects of sphingolipids on fiber development were studied. The inhibitors of serine palmitoyltransferase, including myriocin, dihydrosphingosine, and phytosphingosine, were found to alter cotton fiber elongation. Myriocin suppressed the process, and sphingosine analogues enhanced the elongation. The fiber treatment with phytosphingosine resulted in the enhanced quantity of four lipids and the reduction of 18 of them, which aligned with up- and downregulations of 238 and 194 genes. The important point is that auxin-related genes increased, and their metabolites reduced (
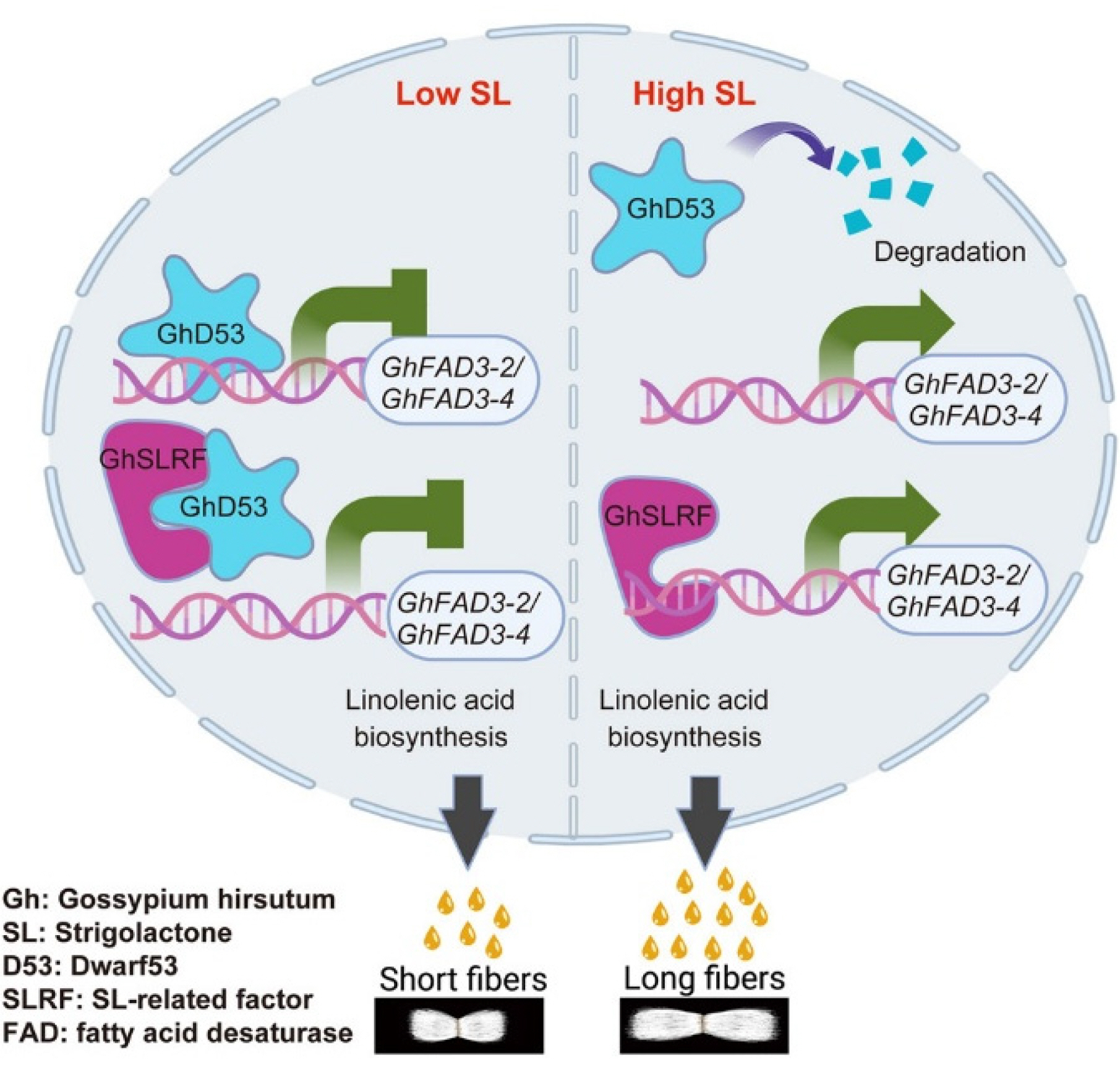
Wang et al. 2024a). GhD53 is another TF involved in fiber elongation, which is known to repress GhFAD3 genes and control the biosynthesis of linolenic acid. GhD53 activation leads to the inhibition of GhFAD3 transcription. Overall results demonstrate the role of strigolactones (SLs) in fiber cell elongation by altering the level of linolenic acid biosynthesis (
Wang et al. 2025a;
Fig. 3).
GhMYB86 is another myeloblastosis TF involved in fiber elongation. It was found to participate in the negative control of fiber elongation. Its interaction with MBE2, linked with sudden ROS increases in a cell, forms the GhMYB86-MBE2 module, which participates in the membrane sphingolipid regulation during fiber elongation. Thus, the involvement of GhMYB86 in fiber elongation was linked with its role in lipid regulation (
Xu et al. 2024).
Sugar Accumulation
Glucose, the monomer of cellulose and hemicellulose, constitutes the primary mass of the fiber. Therefore, the accumulation of glucose is pivotal for fiber elongation and development. Here, we discuss aspects of glucose accumulation relevant to fiber formation. In one unique work, three cotton cultivars varying in their fiber qualities were compared. Correlations were shown among fiber length, fiber strength, and oil content. Differentially expressed levels of genes belonged to various proteins, including enzymes, binding proteins, transporting proteins, kinases, and TFs. MYB and WRKY TFs were found among varying levels. NAC52 was identified as a TF involved in secondary cell wall development. These results explain the involvement of various networks in fiber formation (
Liu et al. 2024a). Gram-domain proteins have been reported to be involved in cotton fiber development. Interactions between TF GhTTG1 and GhGRAM5 glycosyl transferase, as well as between GhHOX1 or GhHD1 and GhGRAM35, were found responsible for fiber elongation among many GhGRAM isoforms. The authors suggested the probability of modulation of histone epigenetic modification of GhTTG1, GhHOX1, or GhHD1 TFs. The genome-wide analysis proved the need of GRAM-domain proteins for fiber development (
Ye et al. 2021). GhDOFD45 is another TF playing a role in fiber elongation. GhSWEET10 genes, encoding the protein that transports sucrose, are controlled by GhDOFD45. Silencing GhSWEET10 or GhDOFD45 resulted in shorter fiber length and smaller seeds, respectively, which was attributed to lower accumulation of sucrose and glucose. The GhSWEET10 overexpression caused fiber elongation due to higher sugar amount transported/accumulated. Thus, the positive controlling role of GhDOFD45 in fiber elongation was established (
Du et al. 2024).
The TFs, including MYB, ERF, and bHLH, have been reported to be included in fiber initiation. The comparison of various cotton lines by using high-throughput sequencing enabled the identification of key molecules in fiber development. The essential roles of the fatty acid pathway and pectin metabolism in fiber development were shown as well (Zou et al. 2022). NAC1 is an NAC TF that activates GhGLU18 glucanase, known to cleave β-1,3-glucan bonds. Its overexpression enhanced cellulose synthesis by promoting the accumulation of polysaccharides. The overexpression of GhGLU18 reconstructed secondary cell wall formation and was concluded to contribute to fiber elongation. Day-dependent expressions of various GLU isoforms were observed between 0 DPA and 25 DPA, with the highest relative expressions in 18-25 DPA. The presence of NAC-binding elements in the secondary cell wall shows the crucial role of NAC TFs (
Fang et al. 2023). In another work, the significance of the modulation of cellulose biosynthesis was shown in an example that was regulated by GhGRF4 TF in the interaction with GhARF2. Thus, GhGRF4/GhARF2 was suggested to enhance the cell wall thickness (
Tian et al. 2024).
Class II KNOX proteins are another class of TFs involved in fiber development. GhKNL1, one of the proteins of this class, was reported to be involved in fiber formation. The GhKNL1-silenced plants developed longer fibers. Reverse results were observed in GhKNL1 dominant lines. It was concluded as one of the potential targets to improve fiber quality. The mechanism of action of GhKNL1 was linked with its function to repress the expression of target genes in cellulose synthesis (
Wang et al. 2022b).
Transcription Factor Interaction Networks Promote Cotton Development and Stress Adaptation
Extensive research has established the crucial roles of transcription factors (TFs) in regulating cotton fiber development and stress responses. These factors operate within complex networks, controlling processes through hierarchical cascades, cooperative and antagonistic protein-protein interactions, and integration with phytohormone, metabolic, and stress signaling pathways (
Liu et al. 2022;
Liu et al. 2024a;
Lu et al. 2022;
Wang et al. 2024b). This section synthesizes key findings on TF-mediated regulation of agronomic traits.
TF interactions are pivotal for fiber development. During elongation, an antagonistic pair embedded in brassinosteroid signaling regulates growth: the bHLH factor GhACE1 directly activates cell wall loosening genes (e.g.,
GhPIP2;7, GhEXP8), while the atypical HLH protein GhFP2 inhibits GhACE1-mediated transcription. GhBZR1 regulates this module by controlling GhFP2 expression (
Lu et al. 2022). The transition to secondary cell wall (SCW) biosynthesis involves cooperative TF complexes. For instance, HD-ZIP III TFs GhHB14_D10 and GhREV_D5 physically interact to co-activate cellulose synthase genes (*
GhCesA4-4*, *
GhCesA7-2*), a process essential for SCW formation (
Liu et al. 2024a). TF activity is also modulated by lipid signaling. The HD-ZIP IV factor GhHOX4 promotes elongation by activating GhEXLB1D and GhXTH2D genes, but phosphatidic acid binds GhHOX4 during the elongation-to-SCW transition, altering its nuclear localization and reducing its transcriptional activity (
Wang et al. 2024b). Fiber SCW development is regulated by the convergence of phytohormone signaling pathways via interacting TF cascades. A transcriptional complex that combines ethylene and auxin signaling is formed when GhERF108 physically interacts with the auxin response proteins GhARF7-1 and GhARF7-2. This combination boosts cellulose deposition in fiber SCWs by activating the MYB transcription factor GhMYBL1, resulting in activation of cellulose synthase genes (
Wang et al. 2023). Hormone signaling and structural cell wall production are clearly linked mechanistically by this hierarchical TF cascade.
The initiation of trichomes and fibers is controlled by multi-component TF networks featuring balanced positive and negative regulation. HD-ZIP IV TFs GhHD1 and GhHOX3 form positive modules that promote epidermal hair fate. Their activity is antagonized by repressors like GhGIR1 and GoSTR, which physically interact with GhHD1/GhHOX3 to inhibit downstream transcription (
Xu et al. 2023;
Yu et al. 2025). Intriguingly, GhHD1 directly activates *GhGIR1/2* transcription, while GhGIR proteins subsequently inhibit GhHD1 transactivation capacity, establishing a precise negative feedback loop that modulates epidermal hair density (
Wang et al. 2024c).
Stress-responsive TF networks integrate transcriptional control with defense signaling. The R2R3-MYB factor GhMYB36 confers drought and Verticillium wilt resistance by directly activating the pathogenesis-related gene PR1, increasing resistance to drought and Verticillium wilt (
Liu et al. 2022). Broader defense networks involve cascading interactions; for instance, group IIc WRKY TFs activate a MAPK cascade (GhMKK2-GhNTF6) that subsequently activates GhMYC2, enhancing flavonoid biosynthesis and disease resistance. Despite functioning as a direct transcriptional activator, transcriptome studies show that GhMYB36 is involved in broader defence-related TF networks. In response to pathogen infection, group IIc WRKY transcription factors activate a MAPK signaling cascade, including GhMKK2, GhNTF6, and GhMYC2, that promotes flavonoid production and disease resistance (
Wang et al. 2022a). Genome-wide studies of the MYB family further indicate extensive indirect connectivity among TFs, with many MYBs potentially linking to central regulators like GhMYC2. The idea of densely interwoven transcriptional regulatory landscapes is further supported by genome-wide investigations of the MYB gene family, which point to indirect connections between several MYB TFs and central regulators like GhMYC2 (
Dai et al. 2023).
TF interaction networks also influence metabolic regulation. GhWRI1, an AP2/EREBP transcription factor, directly bindsAW-box elements in the GhKASI_A05 promoter to regulate cottonseed oil biosynthesis, displaying classic TF-cis-element TF-cis-element interactions associated with natural variation. TF networks also regulate metabolic adaptation. GhWRI1 (AP2/EREBP) directly binds the GhKASI_A05 promoter to regulate oil biosynthesis, an interaction linked to natural variation (
Le et al. 2024). In silico analyses show that Cotton leaf curl Multan virus proteins interact extensively with host transcriptional gene silencing factors such as AGO4, RDRs, and chromatin modifiers, implying that disrupting TF-associated silencing machinery is a key viral pathogenic strategy (
Jain et al. 2024).
Systems biology techniques emphasize higher-order TF interaction networks that regulate complex features beyond specific pathways. Interconnected miRNA-TF-gene regulation modules, including miR164-NAC and miR166-HD-ZIP networks, were found by meta-transcriptomic investigations of heterosis. These modules collectively control cotton growth, metabolism, and stress adaptability (
Hamid et al. 2025).
HD-ZIP III transcription factors create cooperative complexes during the formation of (SCW). Together, GhHB14_D10 and GhREV_D5 physically interact to activate cellulose biosynthesis genes, including GhCesA4-4 and GhCesA7-2, which promote the creation of SCW in cotton fibers (
Li et al. 2024b). This regulatory module is disrupted when either factor is silenced, highlighting the significance of TF-TF collaboration in cell wall formation. Lipid signaling regulates TF activity during fiber formation in addition to TF complexes. GhEXLB1D and GhXTH2D are activated by the HD-ZIP IV transcription factor GhHOX4, which encourages fiber elongation. However, during the transition from elongation to SCW thickening, phosphatidic acid directly binds GhHOX4 and changes its nuclear position, lowering its transcriptional activity (
Wang et al. 2024b).
Multi-component TF interaction networks control the initiation and differentiation of epidermal hair in cotton. GhHD1 and GhHOX3, two HD-ZIP IV transcription factors, create positive regulatory modules that encourage the beginning of trichomes and fibers. Repressors like GhGIR1 and GoSTR, that physically bind to GhHD1/GhHOX3 and inhibit downstream transcription, oppose these complexes (
Xu et al. 2023;
Yu et al. 2025). Interestingly, GhHD1 directly stimulates the transcription of GhGIR1 and GhGIR2, while GhGIR proteins limit GhHD1's capacity to induce transcription, generating a negative feedback loop that regulates epidermal hair density (
Wang et al. 2024c). These reciprocal TF interactions serve as an example of the dynamic regulatory mechanisms that determine the fate of epidermal cells.
In summary, cotton development and stress adaptation are governed by sophisticated TF interaction networks. These networks feature feedback loops, hormone-responsive hierarchies, cooperative/antagonistic complexes, and integration with metabolic and epigenetic systems. Deciphering these interaction frameworks provides a systems-level understanding of complex traits and identifies precise targets for genetic improvement and precision breeding.
Molecular and Biotechnological Advancements in Cotton Fiber Quality
Fiber quality parameters like micronaire, length, strength, and uniformity are conventional quantitative variables governed by numerous loci with minor to moderate impacts. In one study, the authors successfully fine-mapped the fiber micronaire QTL qMIC-A11 to a 407 kb interval and identified the causative gene as
GbGELP, which encodes a GDSL esterase/lipase.
GbGELP expression varied during fiber elongation due to promoter variations influencing an HD-ZIP IV transcription factor binding location. The regulatory divergence between
GbGELP and its
G. hirsutum homolog reveals how modest cis-regulatory polymorphisms can have a major impact on fiber growth, making them useful targets for marker-assisted selection (
Wang et al. 2025b).
Beyond single-locus dissection, large-scale allele and transcriptome mining techniques have increased the number of potential genes involved in fiber quality. Whole-genome variant analysis and transcriptome profiling on 132 historic cotton cultivars to discover unique allelic variation impacting fiber strength, elongation, maturity, and even textile flammability were used. Importantly, this work identified both known QTL-linked genes (e.g., qA13-MIC, qA07-STR) and newly discovered transcription factors, confirming the polygenic nature of fiber properties. The publicly available variation datasets are an invaluable genetic resource for breeders looking to discover beneficial alleles (
Thyssen et al. 2025).
Spatial transcriptomic analyses were used to create a 1-day post-anthesis cotton ovule atlas, which allowed them to analyze tissue-specific gene regulation networks. This method revealed GhLPF1, an R2R3-MYB transcription factor found in the QTL-LP-ChrA06, as a significant regulator of lint percentage. GhLPF1 acts predominantly as a transcriptional repressor in the outer integument epidermis and is controlled by miR828. Its downstream target, GhHB6, was validated as a direct regulatory node with a negative correlation with lint percentage, indicating that GhLPF1 is a promising functional marker for yield improvement (
Wu et al. 2025b).
Abiotic stressors, such as salinity, significantly limit cotton establishment and output, especially on marginal fields. Extracellular ATP (eATP) serves as a signaling molecule, improving salt tolerance while preserving cellular energy homeostasis was found. The transcriptome study indicated substantial reprogramming of genes involved in photosynthesis, hormone signaling, and transcriptional control. Notably, GhG6PD, which encodes glucose-6-phosphate dehydrogenase, was discovered as an important ATP-responsive gene involved in NADPH generation and redox equilibrium. Gene silencing tests demonstrated that disrupting GhG6PD exacerbates oxidative damage under salt stress, emphasizing the importance of energy metabolism in stress tolerance (
Wu et al. 2025a).
While fiber enhancement is still the primary breeding goal, increasing cottonseed value is becoming more important for long-term cotton production. CRISPR/Cas9 genome editing was utilized to disrupt the (+)-δ-cadinene synthase gene (GhCAD), which is essential for gossypol synthesis. Targeted alterations lowered seed gossypol content by up to 64% while maintaining fatty acid composition and protein content. Transcriptome research also demonstrated that the transcription factor GhMYC2-D09 positively regulates gossypol accumulation by indirectly activating GhCAD genes, revealing a jasmonate-associated regulatory layer. This study shows how genome editing can produce exceptional cotton germplasm with higher nutritional value while maintaining agronomic efficiency (
Zhou et al. 2025).
These efforts demonstrate the value of combining QTL mapping, spatial transcriptomics, and genome editing to examine complex characteristics in cotton. The discovery of causative genes such as GbGELP, GbKAP, GhLPF1, and GhG6PD creates actionable targets for marker-assisted selection and precision breeding. Meanwhile, CRISPR/Cas9-based techniques allow for rapid trait change beyond the bounds of natural variation. Future cotton improvement efforts will benefit from merging molecular tools with systems-level regulatory network modeling, which will speed up the production of high-yielding, high-quality, and stress-resistant cotton cultivars.
Conclusion
Cotton fiber elongation is regulated by complex networks utilizing various genes, TFs, phytohormones, lipids, sugars, and other factors. Here, we highlighted the roles of TFs, including MYB, TCP, WRKY, HD-ZIP, bHLH, and others, in the regulation of fiber development. The key points we tried to attract attention to are day-dependent expressions of TFs during fiber elongation and the roles of phytohormones and lipids. Studying these factors in shorter intervals of time can help to specify pathways to understand the complex process more specifically. Despite advances, we lackcomplete understanding of the integrated network that determines fiber quality. The integrations of novel experimental designs, including analyses of TFs, genes, proteins, and metabolites, can improve our understanding of the complex processes underlying fiber development.
Acknowledgments
This work was supported by the Uzbekistan Academy of Sciences.
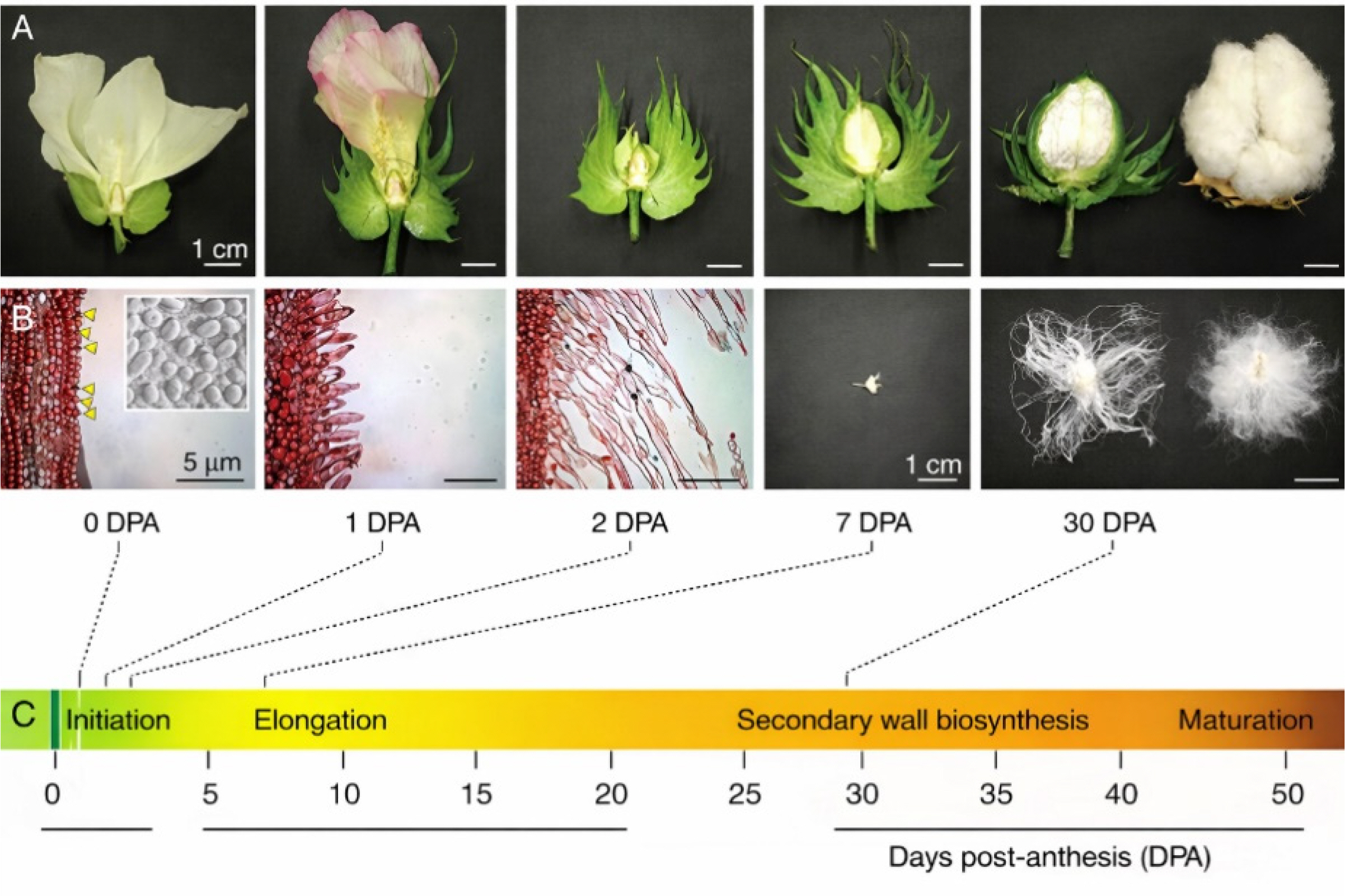
Fig. 1Macro (A) and microscopic (B) depiction of cotton fiber development in various day intervals (C). The figure has been reused from (
Jareczek et al. 2023) with the permission of Frontiers.* For 2-3 days (0-2 DPA), fibers initiate in ovules and turn to elongation, which lasts till 30 DPA. Until 50 DPA, secondary wall is synthesized and strenghtened. Thus, mature cotton fiber is formed. TFs, discussed below, are involved in the fiber development.
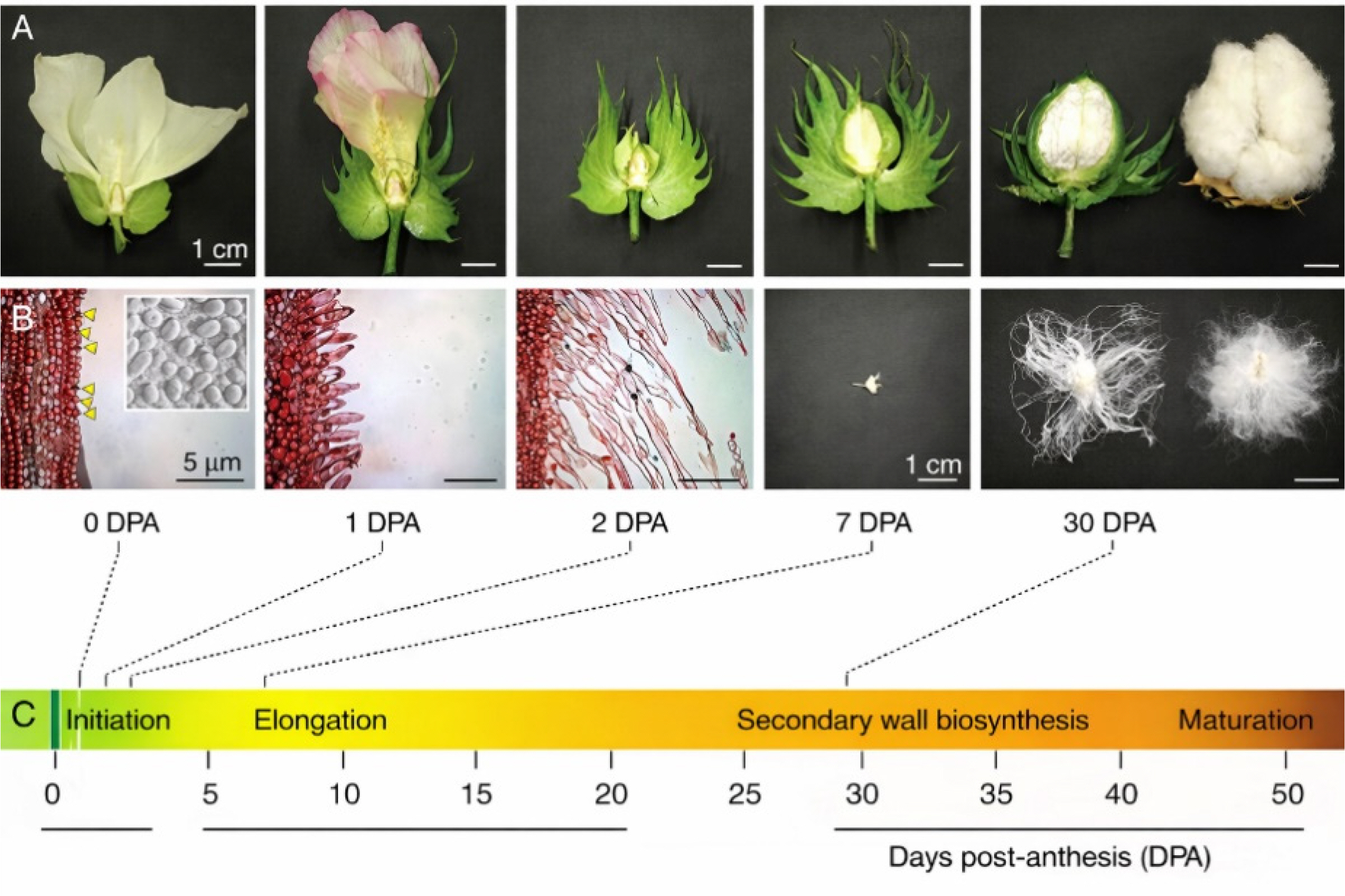
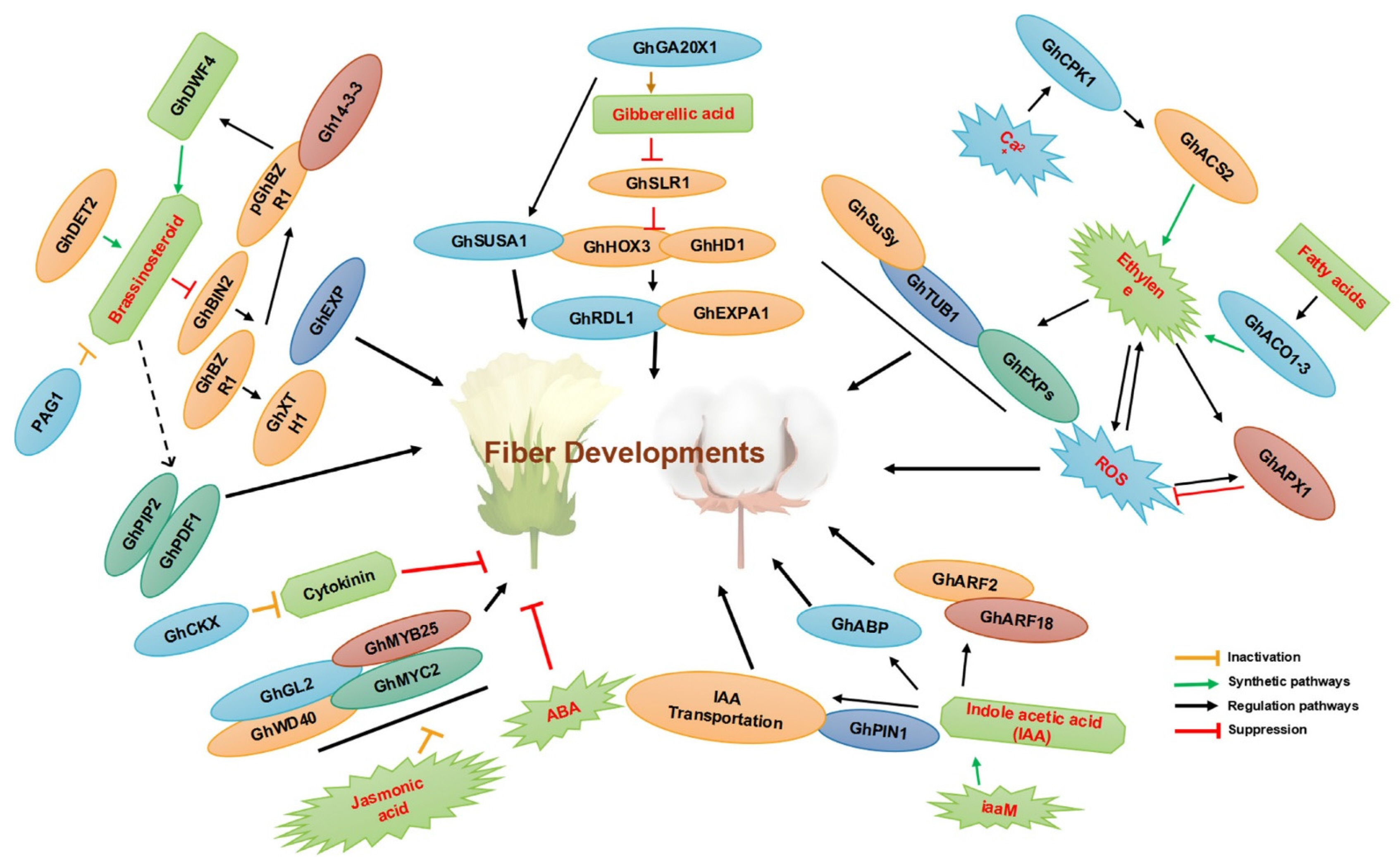
Fig. 2A summary of involvement of TFs and phytohormones in cotton fiber development. The figure has been reused from (
Jan et al. 2022) with the permission of MDPI.
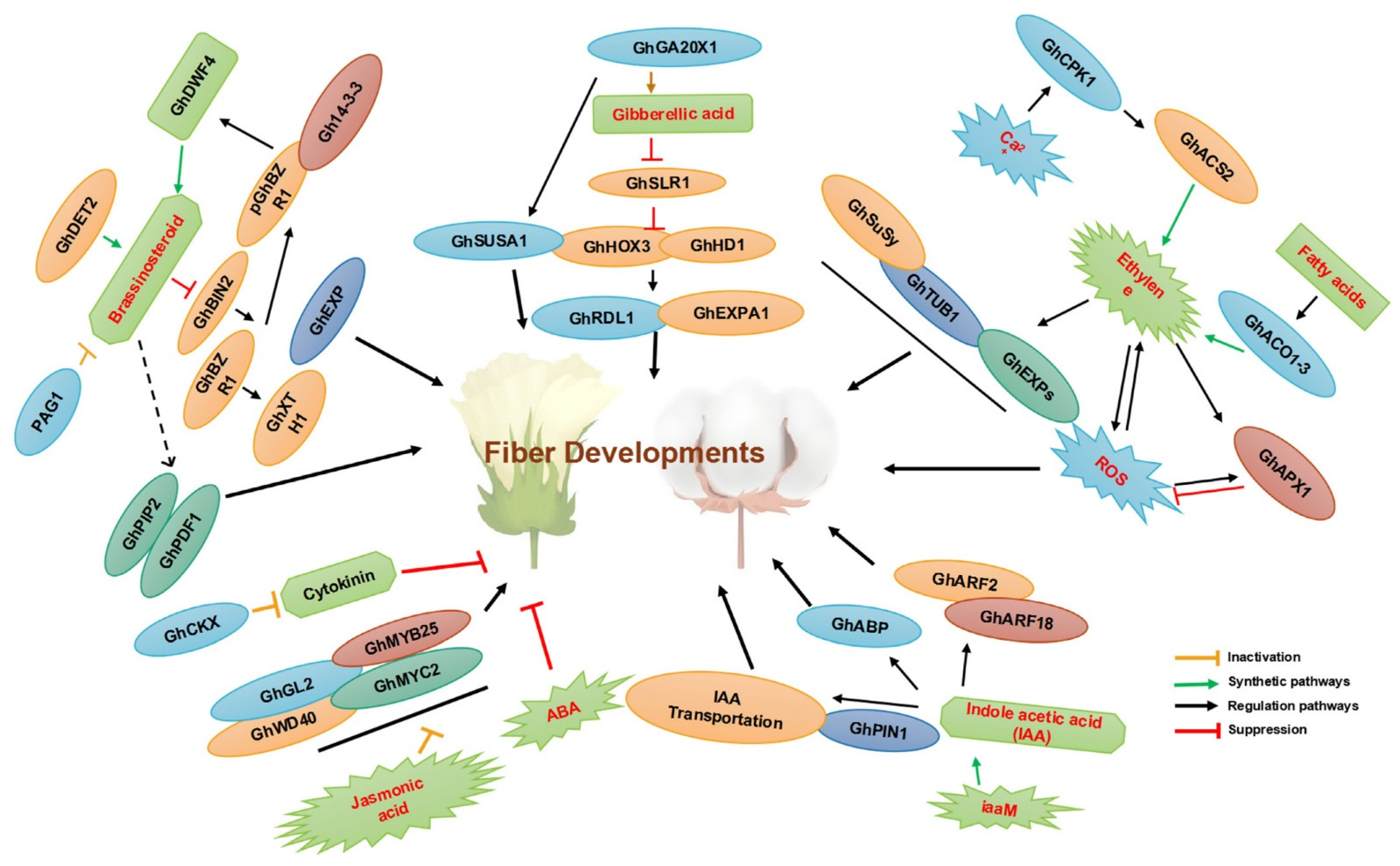
Fig. 3The interaction between GhD53 TF and GhSLRF that contributes to fiber length. The figure has been reused from (
Wang et al. 2025a) with the permission of Elsevier.
References
- Abdurakhmonov IY, Ayubov MS, Ubaydullaeva KA, Buriev ZT, Shermatov SE, Ruziboev HS, et al. 2016. RNA interference for functional genomics and improvement of cotton (Gossypium sp.). Front. Plant Sci. 7: 202
- Abdurakhmonov IY, Buriev ZT, Saha S, Jenkins JN, Abdukarimov A, Pepper AE. 2014. Phytochrome RNAi enhances major fibre quality and agronomic traits of the cotton Gossypium hirsutum L. Nat. Commun. 5: 3062
- Ando A, Kirkbride RC, Jones DC, Grimwood J, Chen ZJ. 2021. LCM and RNA-seq analyses revealed roles of cell cycle and translational regulation and homoeolog expression bias in cotton fiber cell initiation. BMC Genomics. 22(1): 309
- Arora S, Chaudhary B. 2021. Global expression dynamics and miRNA evolution profile govern floral/fiber architecture in the modern cotton (Gossypium). Planta. 254(3): 62
- Bao Y, Wei Y, Liu Y, Gao J, Cheng S, Liu G, et al. 2023. Genome-wide chromatin accessibility landscape and dynamics of transcription factor networks during ovule and fiber development in cotton. BMC Biol. 21(1): 165
- Bao Y, Zou Y, An X, Liao Y, Dai L, Liu L, et al. 2024. Overexpression of a ramie (Boehmaeria nivea L. gaud) group I WRKY gene, BnWRKY49, increases drought resistance in Arabidopsis thaliana. Plants (Basel). 13(3): 379
- Cao JF, Zhao B, Huang CC, Chen ZW, Zhao T, Liu HR, et al. 2020. The miR319-Targeted GhTCP4 promotes the transition from cell elongation to wall thickening in cotton fiber. Mol. Plant. 13(7): 1063-1077.
- Chen G, Liu Z, Li S, Liu L, Lu L, Wang Z, et al. 2023. Characterization of chromatin accessibility and gene expression reveal the key genes involved in cotton fiber elongation. Physiol. Plant. 175(4): e13972
- Chen R, Zhang J, Li J, Chen J, Dai F, Tian Y, et al. 2025. Two duplicated GhMML3 genes coordinately control development of lint and fuzz fibers in cotton. Plant Commun. 6(4): 101281
- Cui S, Zhou X, Xiao G, Feng H. 2024. Genomic analysis of brassinosteroid biosynthesis gene family reveals its roles in cotton development across gossypium species. Biology. 13(6): 380
- Dai Y, Liu S, Zuo D, Wang Q, Lv L, Zhang Y, et al. 2023. Identification of MYB gene family and functional analysis of GhMYB4 in cotton (Gossypium spp.). Mol. Genet. Genomic Med. 298(3): 755-766.
- Du C, Sun W, Song Q, Zuo K. 2024. GhDOFD45 promotes sucrose accumulation in cotton seeds by transcriptionally activating GhSWEET10 expression. Plant J. 120(6): 2468-2484.
- Duan Y, Shang X, He Q, Zhu L, Li W, Song X, et al. 2023. LIPID TRANSFER PROTEIN4 regulates cotton ceramide content and activates fiber cell elongation. Plant Physiol. 193(3): 1816-1833.
- Duan Y, Shang X, Wu R, Yu Y, He Q, Tian R, et al. 2024. The transcription factor GhMYB4 represses lipid transfer and sucrose transporter genes and inhibits fiber cell elongation in cotton. Plant Physiol. 197(1): kiae637
- Fang S, Shang X, He Q, Li W, Song X, Zhang B, et al. 2023. A cell wall-localized β-1,3-glucanase promotes fiber cell elongation and secondary cell wall deposition. Plant Physiol. 194(1): 106-123.
- Hamid R, Panahi B, Jacob F. 2025. Identification of key pathways and associated transcription factor-miRNA-gene regulatory networks driving heterosis in cotton (Gossypium spp.). Funct. Plant Biol. 52(6): FP25041
- Han G, Li Y, Yang Z, Wang C, Zhang Y, Wang B. 2022. Molecular mechanisms of plant trichome development. Front. Plant Sci. 13: 910228
- Hao J, Lou P, Han Y, Chen Z, Chen J, Ni J, et al. 2021. GrTCP11, a Cotton TCP transcription factor, inhibits root hair elongation by down-regulating jasmonic acid pathway in Arabidopsis thaliana. Front. Plant Sci. 12: 769675
- He P, Zhu L, Zhou X, Fu X, Zhang Y, Zhao P, et al. 2024. Gibberellic acid promotes single-celled fiber elongation through the activation of two signaling cascades in cotton. Dev. Cell. 59(6): 723-739.e4.
- Huang G, Huang JQ, Chen XY, Zhu YX. 2021. Recent advances and future perspectives in cotton research. Annu. Rev. Plant Biol. 72: 437-462.
- Jain H, Rawal E, Kumar P, Sain SK, Siwach P. 2024. In silico investigation of the interactions between cotton leaf curl multan virus proteins and the transcriptional gene silencing factors of Gossypium hirsutum L. J. Mol. Evol. 92(6): 891-911.
- Jan M, Liu Z, Guo C, Sun X. 2022. Molecular regulation of cotton fiber development: A review. Int. J. Mol. Sci. 23(9): 5004
- Jareczek JJ, Grover CE, Wendel JF. 2023. Cotton fiber as a model for understanding shifts in cell development under domestication. Front. Plant Sci. 14: 1146802
- Jin F, Zhu L, Hou L, Li H, Li L, Xiao G. 2024. Auxin resistant 2 and short hypocotyl 2 regulate cotton fiber initiation and elongation. Plant Physiol. 195(3): 2032-2052.
- Le Y, Zhao W, Liu X, Chen M, Xiong X, Zhang X, et al. 2024. Natural variation in GhKASI_A05 modulates cottonseed oil content in Gossypium hirsutum L. Plant Physiol. Biochem. 216: 109159
- Li C, Zhao J, Liu Z, Yang Y, Lai C, Ma J, et al. 2024a. Comparative transcriptomic analysis of Gossypium hirsutum fiber development in mutant materials (xin w 139) provides new insights into cotton fiber development. Plants (Basel). 13(8): 1127
- Li S, Yu M, Qanmber G, Feng M, Hussain G, Wang Y, et al. 2024b. GhHB14_D10 and GhREV_D5, two HD-ZIP III transcription factors, play a regulatory role in cotton fiber secondary cell wall biosynthesis. Plant Cell Rep. 43(3): 76
- Liu L, Chen G, Li S, Gu Y, Lu L, Qanmber G, et al. 2023. A brassinosteroid transcriptional regulatory network participates in regulating fiber elongation in cotton. Plant Physiol. 191(3): 1985-2000.
- Liu L, Grover CE, Kong X, Jareczek J, Wang X, Si A, et al. 2024a. Expression profile analysis of cotton fiber secondary cell wall thickening stage. PeerJ. 12: e17682
- Liu T, Chen T, Kan J, Yao Y, Guo D, Yang Y, et al. 2022. The GhMYB36 transcription factor confers resistance to biotic and abiotic stress by enhancing PR1 gene expression in plants. Plant Biotechnol. J. 20(4): 722-735.
- Liu Y, Ma X, Li Y, Yang X, Cheng W. 2024b. Zinc finger protein8 (GhZFP8) regulates the initiation of trichomes in Arabidopsis and the development of fiber in cotton. Plants (Basel). 13(4): 492
- Liu ZH, Chen Y, Wang NN, Chen YH, Wei N, Lu R, et al. 2020. A basic helix-loop-helix protein (GhFP1) promotes fibre elongation of cotton (Gossypium hirsutum) by modulating brassinosteroid biosynthesis and signalling. New Phytol. 225(6): 2439-2452.
- Lu R, Li Y, Zhang J, Wang Y, Zhang J, Li Y, et al. 2022. The bHLH/HLH transcription factors GhFP2 and GhACE1 antagonistically regulate fiber elongation in cotton. Plant Physiol. 189(2): 628-643.
- Qin L, He H, Yang L, Zhang H, Li J, Zhu Y, et al. 2024. AtZAT10/STZ1 improves drought tolerance and increases fiber yield in cotton. Front. Plant Sci. 15: 1464828
- Shi H, Qanmber G, Yang Z, Guo Y, Ma S, Shu S, et al. 2025. An AP2/ERF transcription factor GhERF109 negatively regulates plant growth and development in cotton. Plant Sci. 352: 112365
- Shi Z, Chen X, Xue H, Jia T, Meng F, Liu Y, et al. 2022. GhBZR3 suppresses cotton fiber elongation by inhibiting very-long-chain fatty acid biosynthesis. Plant J. 111(3): 785-799.
- Song Q, Du C, Xu Y, Wang J, Lin M, Zuo K. 2024. Transcriptional regulation of phospholipid transport in cotton fiber elongation by GhMYB30D04-GhHD1 interaction complex. J. Integr. Plant Biol. 66(11): 2431-2449.
- Sun H, Hao P, Gu L, Cheng S, Wang H, Wu A, et al. 2020. Pectate lyase-like Gene GhPEL76 regulates organ elongation in Arabidopsis and fiber elongation in cotton. Plant Sci. 293: 110395
- Sun Y, Zhang D, Zheng H, Wu Y, Mei J, Ke L, et al. 2022. Biochemical and expression analyses revealed the involvement of proanthocyanidins and/or their derivatives in fiber pigmentation of gossypium stocksii. Int. J. Mol. Sci. 23(2): 1008
- Suo Q, Fang N, Zeng J, Yan F, Zhu X, Wang Y, et al. 2024. R2R3 MYB transcription factor GhMYB201 promotes cotton fiber elongation via cell wall loosening and very-long-chain fatty acid synthesis. Int. J. Mol. Sci. 25(17): 9559
- Thyssen GN, Smith W, Naoumkina M, Pinnika G, Jenkins JN, McCarty JC, et al. 2025. Allele and transcriptome mining in Gossypium hirsutum reveals variation in candidate genes at genetic loci affecting cotton fiber quality and textile flammability. BMC Plant Biol. 25(1): 305
- Tian Z, Chen B, Sun Y, Sun G, Gao X, Pan Z, et al. 2024. GhGRF4/GhARF2-GhGASA24 module regulates fiber cell wall thickness by modulating cellulose biosynthesis in upland cotton (Gossypium hirsutum). Plant J. 120(5): 1842-1856.
- Tian Z, Zhang Y, Zhu L, Jiang B, Wang H, Gao R, et al. 2022. Strigolactones act downstream of gibberellins to regulate fiber cell elongation and cell wall thickness in cotton (Gossypium hirsutum). Plant Cell. 34(12): 4816-4839.
- Wang H, Zhu L, Fan M, Weng S, Zhou X, Zhao H, et al. 2025a. Strigolactone promotes cotton fiber cell elongation by de-repressing DWARF53 on linolenic acid biosynthesis. Dev. Cell. 60(7): 1101-1117.e7.
- Wang L, Guo D, Zhao G, Wang J, Zhang S, Wang C, et al. 2022a. Group IIc WRKY transcription factors regulate cotton resistance to Fusarium oxysporum by promoting GhMKK2-mediated flavonoid biosynthesis. New Phytol. 236(1): 249-265.
- Wang L, Jin C, Zhang W, Mei X, Yu H, Wu M, et al. 2024a. Sphingosine promotes fiber early elongation in upland cotton. Plants (Basel). 13(14): 1993
- Wang N-N, Li Y, Chen Y-H, Lu R, Zhou L, Wang Y, et al. 2021a. Phosphorylation of WRKY16 by MPK3-1 is essential for its transcriptional activity during fiber initiation and elongation in cotton (Gossypium hirsutum). Plant Cell. 33(8): 2736-2752.
- Wang NN, Ni P, Wei YL, Hu R, Li Y, Li XB, et al. 2024b. Phosphatidic acid interacts with an HD-ZIP transcription factor GhHOX4 to influence its function in fiber elongation of cotton (Gossypium hirsutum). Plant J. 118(2): 423-436.
- Wang X, Dai P, Li H, Wang J, Gao X, Wang Z, et al. 2024c. The genetic basis of leaf hair development in upland cotton (Gossypium hirsutum). Plant J. 120(2): 729-747.
- Wang X, Shen C, Meng P, Tan G, Lv L. 2021b. Analysis and review of trichomes in plants. BMC Plant Biol. 21(1): 70
- Wang Y, Li Y, Gong SY, Qin LX, Nie XY, Liu D, et al. 2022b. GhKNL1 controls fiber elongation and secondary cell wall synthesis by repressing its downstream genes in cotton (Gossypium hirsutum). J. Integr. Plant Biol. 64(1): 39-55.
- Wang Y, Li Y, He SP, Xu SW, Li L, Zheng Y, et al. 2023. The transcription factor ERF108 interacts with AUXIN RESPONSE FACTORs to mediate cotton fiber secondary cell wall biosynthesis. Plant Cell. 35(11): 4133-4154.
- Wang Z, Cai C, Yu Z, Reyimu Y, Han X, Lv M, et al. 2025b. A GDSL esterase/lipase gene GbGELP identified from a fiber micronaire QTL qMIC-A11 modulates cell elongation and fiber development. Theor. Appl. Genet. 138(6): 126
- Wen X, Chen Z, Yang Z, Wang M, Jin S, Wang G, et al. 2023. A comprehensive overview of cotton genomics, biotechnology and molecular biological studies. Science China. Life Sci. 66(10): 2214-2256.
- Wu C, Xiao S, Zhang X, Ren W, Shangguan X, Li S, et al. 2024. GhHDZ76, a cotton HD-Zip transcription factor, involved in regulating the initiation and early elongation of cotton fiber development in G. hirsutum. Plant Sci. 345: 112132
- Wu F, Chen X, Wang J, Yang J, Wang L, Lan H, et al. 2025a. eATP activates energy dynamic gene GhG6PD to improve salt tolerance in cotton. Plant Physiol. Biochem. 229(Pt C): 110553
- Wu H, Wang L, Zhao S, Gao M, Cao J, Hao Y, et al. 2025b. GhLPF1 associated network is involved with cotton lint percentage regulation revealed by the integrative analysis of spatial transcriptome. Adv. Sci. (Weinh). 12(13): e2414175
- Xing K, Liu Z, Liu L, Zhang J, Qanmber G, Wang Y, et al. 2023. N(6) -Methyladenosine mRNA modification regulates transcripts stability associated with cotton fiber elongation. Plant J. 115(4): 967-985.
- Xu B, Zhang J, Shi Y, Dai F, Jiang T, Xuan L, et al. 2023. GoSTR, a negative modulator of stem trichome formation in cotton. Plant J. 116(2): 389-403.
- Xu F, Li G, He S, Zeng Z, Wang Q, Zhang H, et al. 2024. Sphingolipid inhibitor response gene GhMYB86 controls fiber elongation by regulating microtubule arrangement. J. Integr. Plant Biol. 66(9): 1898-1914.
- Yang J, Gao L, Liu X, Zhang X, Wang X, Wang Z. 2021. Comparative transcriptome analysis of fiber and nonfiber tissues to identify the genes preferentially expressed in fiber development in Gossypium hirsutum. Sci. Rep. 11(1): 22833
- Yang Z, Liu Z, Ge X, Lu L, Qin W, Qanmber G, et al. 2023. Brassinosteroids regulate cotton fiber elongation by modulating very-long-chain fatty acid biosynthesis. Plant Cell. 35(6): 2114-2131.
- Ye Z, Qiao L, Luo X, Chen X, Zhang X, Tu L. 2021. Genome-wide identification of cotton GRAM family proteins reveals that GRAM31 regulates fiber length. J. Exp. Bot. 72(7): 2477-2490.
- Yu L, Zhao X, Hua L, Yuan R, Wei Y, Luo K, et al. 2025. Functionally differentiated GL2-interacting-repressor 1 homoeologs regulate epidermal hair development of Gossypium hirsutum. Plant Physiol. 198(2): kiaf184
- Zhai Z, Zhang K, Fang Y, Yang Y, Cao X, Liu L, et al. 2023. Systematically and comprehensively understanding the regulation of cotton fiber initiation: A review. Plants (Basel). 12(21): 3771
- Zhang R, Shen C, Zhu D, Le Y, Wang N, Li Y, et al. 2022. Fine-mapping and candidate gene analysis of qFL-c10-1 controlling fiber length in upland cotton (Gossypium hirsutum L.). Theor. Appl. Genet. 135(12): 4483-4494.
- Zhang X, Cao J, Huang C, Zheng Z, Liu X, Shangguan X, et al. 2021. Characterization of cotton ARF factors and the role of GhARF2b in fiber development. BMC Genomics. 22(1): 202
- Zhou L, Wang Y, Wang J, Wang P, Lu G, Su X, et al. 2025. CRISPR/Cas9-mediated mutation of GhCAD decreases the gossypol content of cottonseed. J Biol Eng. 19(1): 83
- Zhu L, Wang H, Zhu J, Wang X, Jiang B, Hou L, et al. 2023. A conserved brassinosteroid-mediated BES1-CERP-EXPA3 signaling cascade controls plant cell elongation. Cell Rep. 42(4): 112301