Abstract
Grape (Vitis spp.), which is an important economic fruit crop worldwide, is the fourth most common fruit produced in Korea. Low temperature is one of the most important environmental factors that affect the successful cultivation of grapes. To screen for specific expression of abiotic stress-related genes against cold treatment in ‘Campbell Early’ and ‘Muscat Bailey A’ grapevines, the expression of various defense-related genes was investigated by real-time PCR and semi-quantitative RT-PCR. Among 67 genes analyzed by real-time PCR and RT-PCR, 43 and 10 were found to be up-regulated, while 4 and 37 were down-regulated in low temperature treated ‘Campbell Early’ and ‘Muscat Baily A’ grapevines, respectively. Genes encoding cold induced protein (CIP), lipoxygenase (LOX), glutathione S-transferase (GST), polygalacturonase-inhibiting protein (PGIP), and cold acclimation specific protein (CAS15A) were up-regulated, while those encoding cinnamyl alcohol dehydrogenase 2 (CAD2) and cold acclimation protein (WCS120) were down-regulated by low temperature treatment in both ‘Campbell Early’ and ‘Muscat Bailey A’. These changes in gene expression showed that defense response-related genes for cold acclimation were induced and wheat cold-specific gene decreased in grapevines. Among tested genes, the transcript levels of CLP, cell wall protein (CW), TIP, TLP, and WRKY transcription factor 10 (WRKY10) increased considerably in response to low temperature in ‘Campbell Early’ grapevine. These findings suggest that differentially expressed genes could be utilized as candidates for useful genetic resources and molecular markers after additional confirmation in various grapevine cultivars treated with low temperature in breeding programs for grapes with cold-tolerance.
-
Key words: Cold stress, Freezing tolerance, Grape breeding, Vitis
INTRODUCTION
Grapes (
Vitis spp.) are an important economic fruit crop cultivated globally; however, they often encounter cold stresses due to changes in environmental conditions. Accordingly, cold damage caused by extremely low temperature is a serious problem associated with grape production. As grapevines are deciduous perennial plants, they acquire tolerance to freezing in advance of annual freezes to enable to overwinter in canes and buds (
Sreekantan et al. 2010).
Crops respond and adapt to cold stress by changing their patterns of gene expression (Yang et al. 2011). Many cold responsive genes and their gene products are considered to function in cold tolerance at transcriptional and biochemical levels (
Sanghera et al. 2011).
Consistent with the expected effects of a calcium signal, cold-acclimation causes phosphorylation of a number of proteins (
Momroy and Dhindsa 1995;
Monroy et al. 1993). When plants are exposed to cold or chilling stress at temperatures of 0 to −15°C, they attempt to maintain homeostasis to acquire freezing tolerance, which involves extensive reprogramming of gene expression and metabolism and changes in structural components (
Hughes and Dunn 1990). Both cold acclimation and chilling tolerance are involved in an array of biochemical, molecular and metabolic processes (
Cook et al. 2004;
Kotak et al. 2007;
Larkindale et al. 2005;
Thomashow 1999;
Zhu et al. 2007). Therefore, understanding the molecular basis of cold stress responses in plants will help us develop cold-tolerant grape cultivars. Recently, damage by extremely low temperatures during winter has become a serious problem in viticulture in Korea (
Ahn et al. 2012); therefore, programs for selection of new grape cultivars tolerant to coldness are required. This study was conducted to obtain a better understanding of the general differential expression and provide a basis for further analysis. The specific goals of this study were to detect differentially expressed genes in grapevines following low temperature treatment and provide useful genetic resources for breeding programs of cold-tolerant grapes.
MATERIALS AND METHODS
Plant materials and cold treatment
One-year old vine cane with latent buds of ‘Campbell Early’(moderately tolerant to cold stress) and ‘Muscat Bailey A’ (MBA, sensitive to cold stress) grapevines were harvested in February 2011, cut to small cuttings with 1 bud, and used for cold treatment. Cold treatment of buds was carried out at −20°C in the dark for 0, 6, 12, and 24 h, respectively. The buds treated with coldness were collected at the indicated times, frozen in liquid nitrogen and stored at −80°C until further use. Untreated buds were collected for use as a control (0 h).
RNA isolation and cDNA synthesis
Total RNA was extracted from grapevine buds using a modified version of the method described by
Yang et al. (2008). Fifty buds from each treatment were used for RNA extraction. The yield and quality of total RNA were measured based on the absorbance at 230, 260, and 280 nm using a NanoDrop spectrophotometer (ND-1000, Technologies Inc., USA). First-strand cDNA was synthesized from the total RNA (500 ng) using a PrimeScript
TM 1
st strand cDNA synthesis kit (Takara Bio Inc., Japan) and subsequently used as a template for PCR.
The various defense-related genes showing changes of their expression against foreign stresses were selected to analyze their expression patterns to cold treatment in grapevines. The expression of selected genes was tested by real-time PCR and semi-quantitative RT-PCR using gene specific primer pairs (
Table 1). Real-time PCR was performed on a C1000
TM Thermal Cycler (BioRad, USA) using SYBR Premix Ex (TaKaRa Bio Inc., Japan) as the fluorescent dye. Amplification was conducted by subjecting the samples to one cycle at 95°C for 5 min, 40 cycles of 95°C for 20 s, 60°C for 30 s and 72°C for 30 s. All reactions were performed in triplicate to ensure consistency of the results. Each real-time assay was tested to ensure that there was onlya single PCR product using a high resolution melting profiling technique. Real-time PCR results were analyzed as previously described (
Wu et al. 2010). Semi-quantitative RT-PCR was performed by subjecting the samples to the following conditions: initial denaturation at 94°C for 5 min, followed by 30 cycles at 94°C for 45 sec, 55°C for 45 sec, and 72°C for 1 min and then final extension for 7 min at 72°C. The actin gene primers were used as an internal control. The PCR products were identified by 1.5% (W/V) agarose gel electrophoresis with 0.5X TBE running buffer. After taking the gel picture using a transilluminator (UVP, CA, USA), the expression levels were measured by analysis of the images using a public domain image analysis system (NIH ImageJ, NIH Image, Bethesda, USA).
REULTS AND DISCUSSION
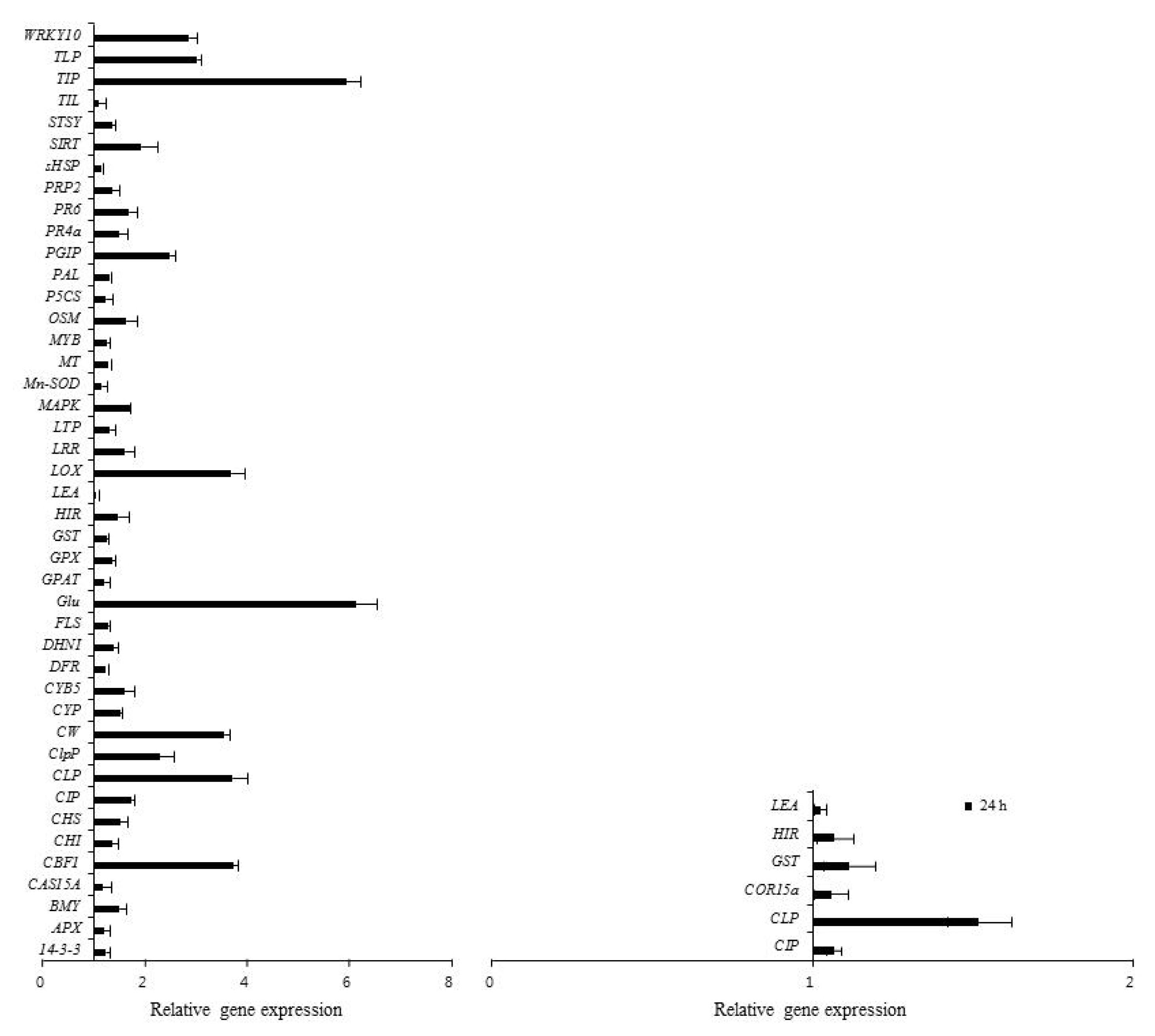
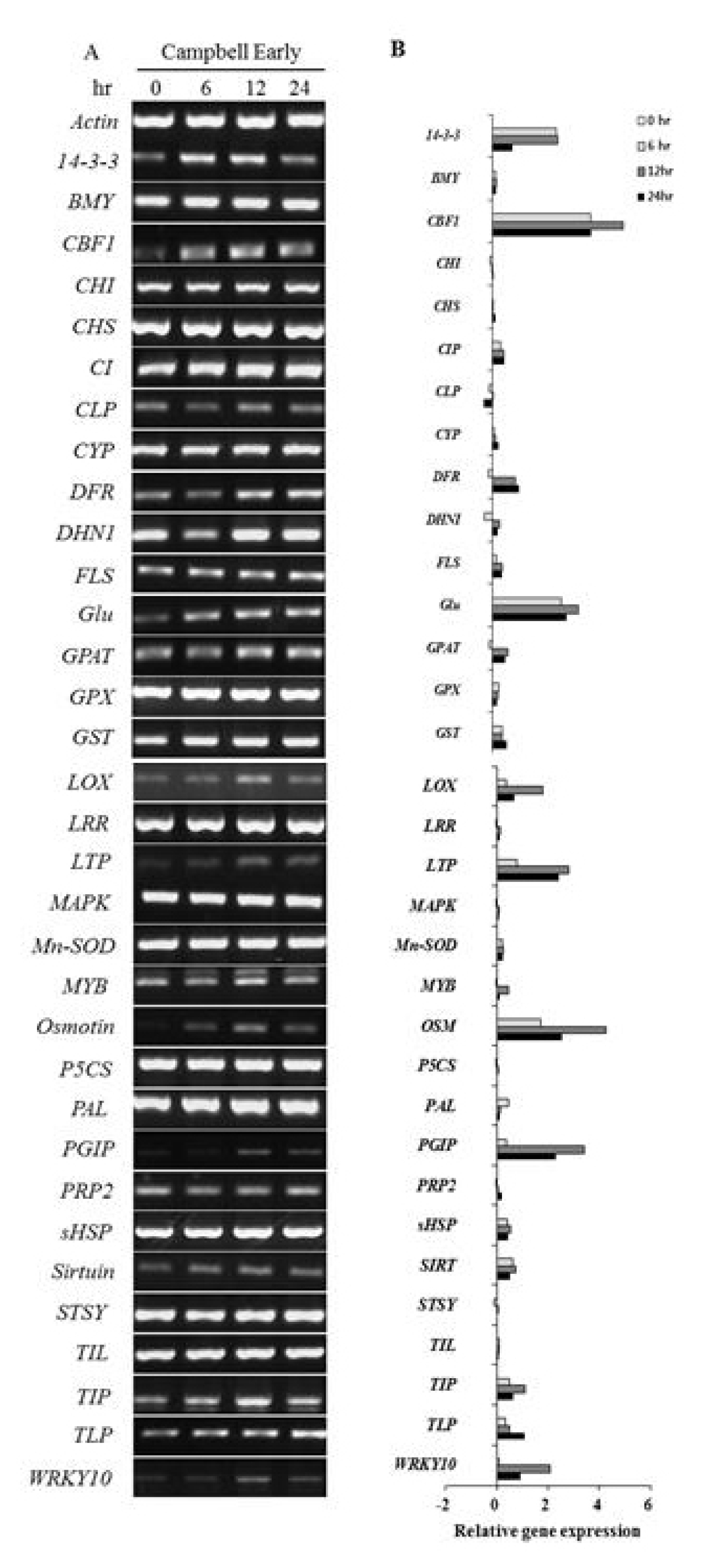
The differential expression of genes was evaluated by real-time PCR and semi-quantitative RT-PCR with primer pairs (
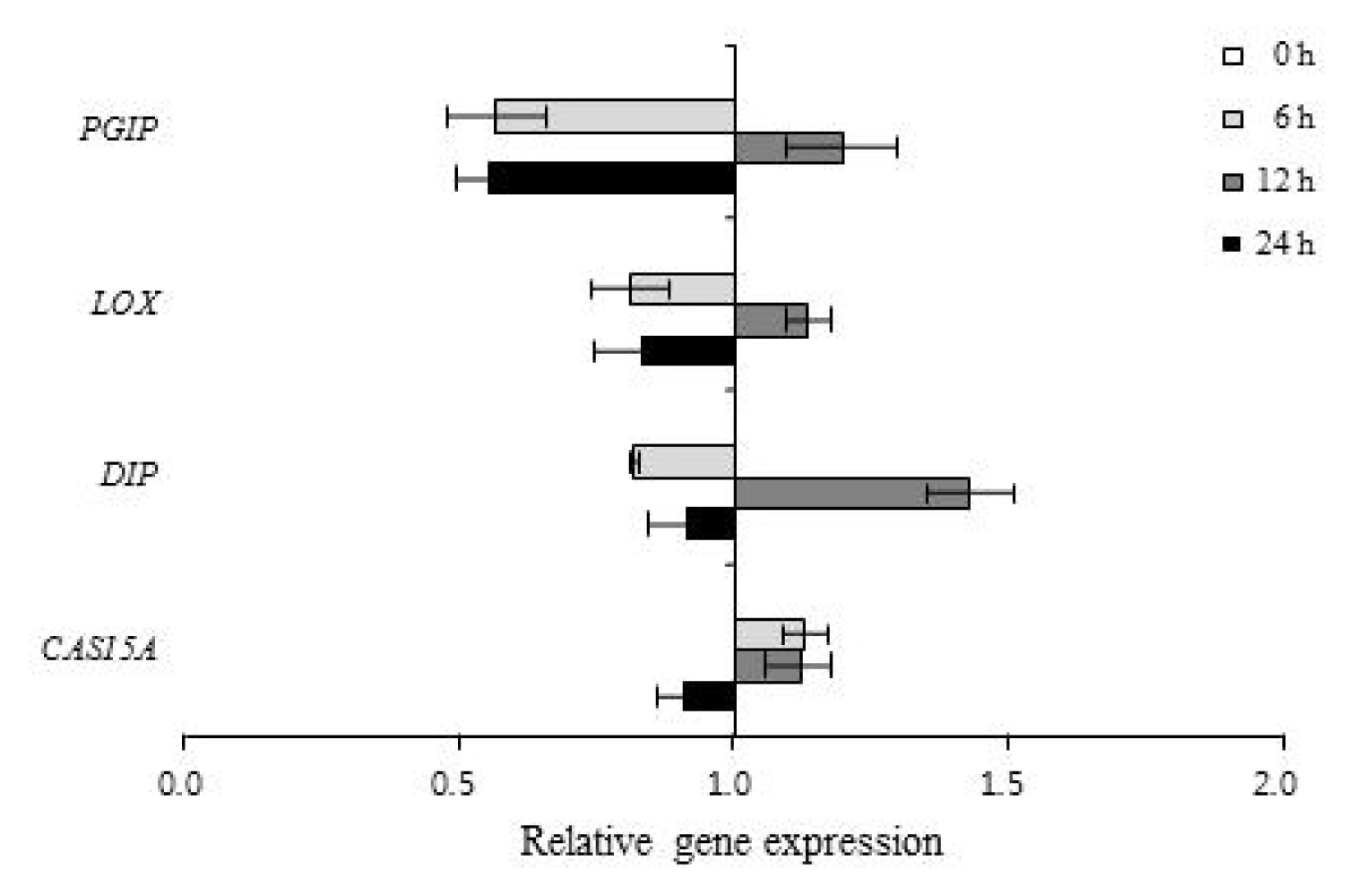
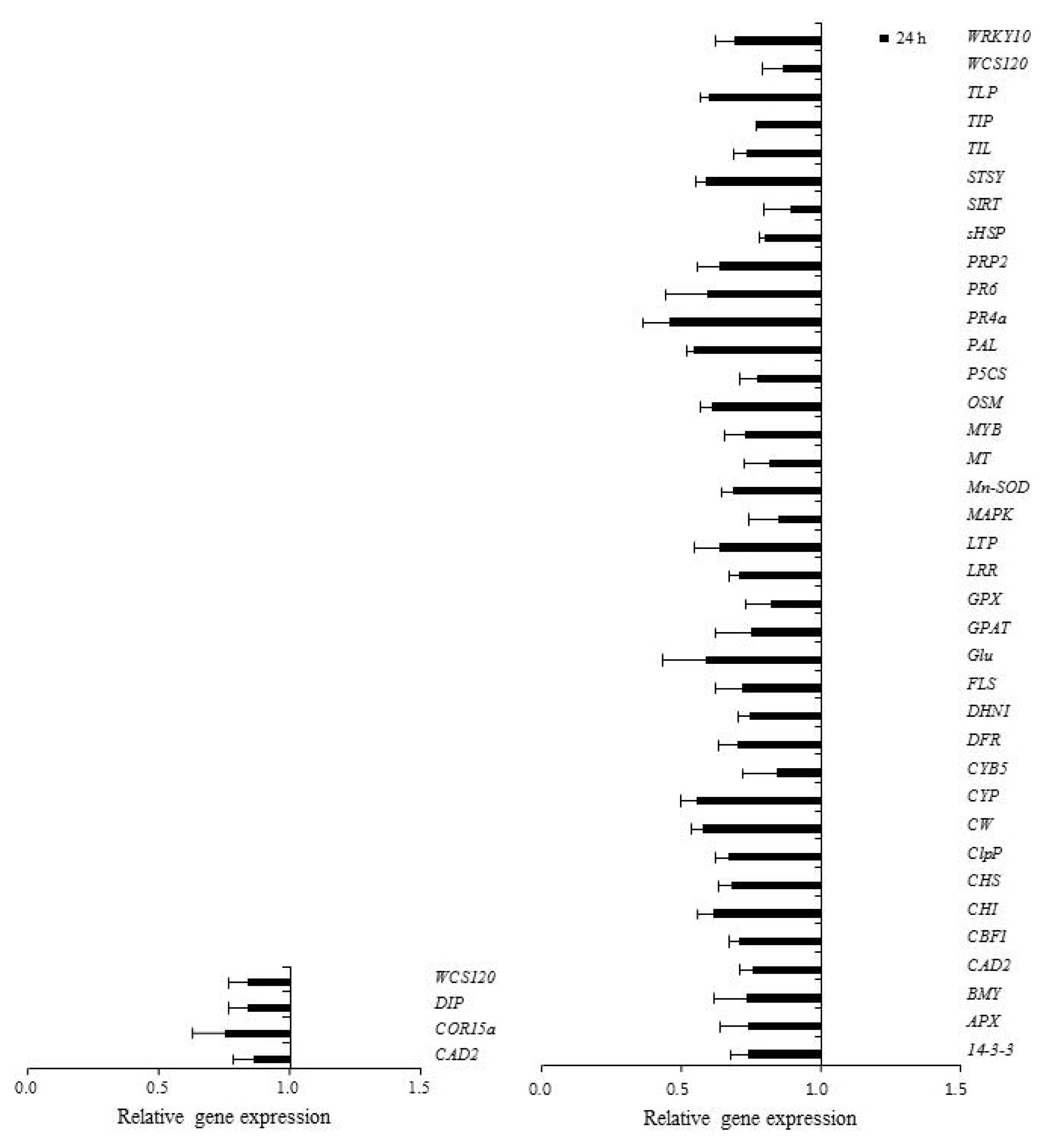
Table 1). The expression patterns of 67 genes involved in defense responses to abiotic stress were evaluated following low temperature treatment of ‘Campbell Early’ and ‘MBA’ grapevines using quantitative real-time PCR (
Fig. 1,
2, and
3). Among the tested genes, 43 and 10 were up-regulated, while 4 and 37 genes were down-regulated by low temperature treatment in ‘Campbell Early’ and ‘MBA’, respectively.
Among the tested genes, the transcript levels of cold-tolerance such as CBF like transcription factor (
CBF1), ATP dependent Clp protease (
ClpP), cell wall protein (
CW), sirtuin (
SIRT), tonoplast intrinsic protein (
TIP), pathogenesis-related (PR) genes such as chitinase (
CLP), β-1,3-glucanase (
Glu), and thaumatin (
TLP), lipoxygenase (
LOX), polygalacturonase-inhibiting protein (
PGIP), and WRKY transcription factor 10 (
WRKY10) increased considerably in response to low temperature at 24 h after treatment. Various genes related with defense-response and cold acclimation were up-regulated in cold-treated grapevines. In ‘MBA’,
CLP and glutathione
S-transferase (
GST) were highly up-regulated by low temperature 24 h after treatment (
Fig. 1). The expression of cold acclimation-specific protein (
CAS15A),
DIP,
LOX and
PGIP increased in response to low temperature treatment for 12 h when compared with untreated controls in ‘MBA’ (
Fig. 2).
However, all genes which were up-regulated by low temperature in ‘Campbell Early’ showed decreases in gene expression values in ‘MBA’. The greatest decreases were observed in
PR4a and stilbene synthase (
SYST) gene expression in ‘MBA’ grapevines treated with low temperature (
Fig. 3). The expression of genes related with defense responses such as PR-proteins decreased in grapevines treated with low-temperature.
Among the screened genes, those encoding CAS15A, cold induced protein (CIP), glutathione S-transferase (GST), hypersensitive induced response protein (HIR), and late embryogenesis abundant protein (LEA), were up-regulated, while genes encoding cinnamyl alcohol dehydrogenase2 (CAD2) and cold acclimation protein (WCS120) were down-regulated by low temperature treatment in both ‘Campbell Early’ and ‘MBA’.
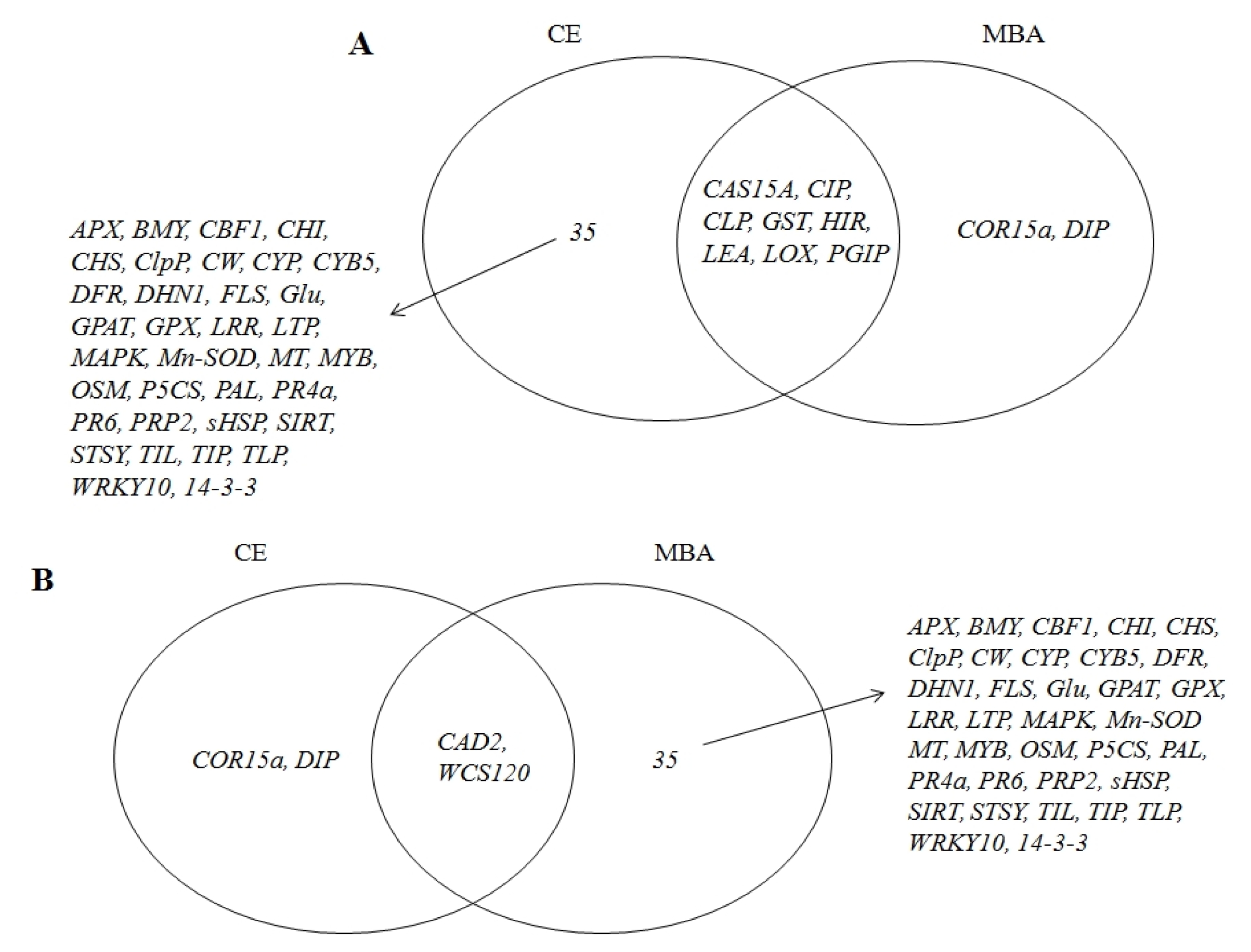
The Venn diagram (
Fig. 4) presents the distribution of significantly induced gene expression patterns in response to the low-temperature treatment in both ‘Campbell Early’ and ‘MBA’. Overall, 35 genes encoding ascorbate peroxidase (
APX),
CBF1, chalcone synthesis related (
CHS and
CHI),
ClpP,
CW, cytochrome B5 (
CYB5),
Glu, glycerol-3-phosphate acyltransferase (
GPAT), leucine rich repeat (
LRR), lipid transfer protein (
LTP), O-methyltransferase (
MT), MYB transcription factor (
MYB), phenylalanine ammonia lyase (
PAL), pathogen related proteins (
PR4a and
PR6), small heat shock protein (
sHSP),
SIRT, STSY, TIP, TLP, WRKY10, and regulator of cellular molecules (
14-3-3) were up-regulated in ‘Campbell Early’ and down-regulated in ‘MBA’ in response to low-temperature treatment. These changes in gene expression showed that defense response-related genes for cold acclimation were induced and wheat cold-specific gene decreased in grapevines. However,
DIP and cold regulated protein 15a (
COR15a), which were up-regulated in ‘MBA’, were down-regulated in ‘Campbell Early’ in response to low-temperature treatment. It is worth noting that the expression of the 35 genes following cold treatment differed between ‘MBA’ (sensitive) and ‘Campbell Early’ (moderately tolerant) grapevines. The differential expression of the described genes likely resulted from the difference in responses to cold and cold-acclimation capabilities. Therefore, these candidate sequences with different expression patterns in response to cold could be useful as functional molecular markers to assist in selecting cold-tolerant vines for grape breeding programs. The significant differences in the expression of the aforementioned genes were confirmed by RT-PCR. Specifically, 33 of the aforementioned genes were analyzed by RT-PCR and their expression patterns were found to be consistent with the results of real-time PCR (
Fig. 5 and
6).
Low temperature affects virtually all aspects of cellular function in plants. Lipid molecules are also very important in signal transduction during cold stress (
Yadav 2010).
COR15a expression resulted in a decrease in the propensity of the membranes to form hexagonal II phase lipids in response to freezing (
Uemura and Steponkus 1997). WCS120 protein acts as an important protective agent of many vital cellular proteins in cold-acclimated plant tissue (
Houde et al. 1995;
Nikolai et al. 2008;
Sarhan et al. 1997). Although genes encoding the WCS120 protein were reported to be induced in wheat and barley exposed to low temperature, they showed decrease of expression in 2 grapevine cultivars treated with freezing temperature in this study. Their expression and roles in defense responses to low temperature in plants including grapevines should be studied in the near future. The relationship between
WRKY and abiotic stress responses has been reported in some plant species. For example, at least 4
WRKY genes were shown to be involved in the drought, cold, or salinity responses in Arabidopsis (
Seki et al. 2002).
Generally, expression of late embryogenesis abundant proteins (
LEA) responsible for the stabilization of membranes and proteins through detergent-like or chaperone activities (
Close 1997) and heat shock proteins (
HSPs) responsible for the functional conformations of the protein (
Wang et al. 2004) are induced in response to different abiotic stresses in plants. Various stress-responsive genes were induced by drought, high-salinity, cold and ABA in the abiotic-stress signal transduction pathway systems (
Ishitani et al. 1997; Xiong and Zhu 2002).
TIL gene expression was up-regulated during cold acclimation and heat-shock treatment from wheat and Arabidopsis (
Charron et al. 2002). In addition, the accumulation of compatible osmolytes and antioxidants was stimulated by many factors including cold, drought, and salt stress in plants, and such changes caused by cold stress adversely affected the growth and development of plants (
Hasegawa et al. 2000).
Zhang et al. (2005) reported that transcript levels of
CHI, which is related to anthocyanin synthesis, differed among interspecies, but were elevated to much higher levels in
Poncirus rootstock during cold acclimation.
PAL and
CHS mRNAs were accumulated at similar levels in response to low temperatures (
Leyva et al. 1995).
In plants, cold, drought, and salt stresses stimulated the accumulation of compatible osmolytes and antioxidants (
Hasegawa et al. 2000).
Xiong and Zhu (2001) reported that the signal transduction networks for cold, drought, and salt stress could be divided into three major signaling types, and that among them, Type I signaling may contribute to the production of compatible osmolytes and antioxidants, and may also be related to cell cycle regulation under osmotic stress.
Seki et al. (2001) reported that, in Arabidopsis, induction of
β-amylase occurred as early as 2 h after exposure to cold stress.
Kaplan and Guy (2004) reported that
BMY8 expression increased dramatically as early as 6 h and peaked at 24 h, then decreased gradually, but remained higher under cold shock at 5°C than control levels at 192 h.
The accumulation of
PGIP transcripts after storage in the cold has been reported in apple (
Yao et al. 1999), and cold induced the expression of
AtPGIP1, but not
AtPGIP2, has been observed in Arabidopsis (
Ferrari et al. 2003). These findings suggested that low temperatures can increase susceptibility to diseases, and that cold induction of defensive proteins might provide protection from pathogen infections. Finally, antimicrobial proteins such as
β-glucanase and chitinase, which are highly homologous with PR proteins, were accumulated in the leaf apoplast of winter rye after cold exposure (Hon et al. 1995).
Plant temperature-induced lipocalin and HSP are thought to participate in plant responses to heat and cold stress (
Chi et al. 2009), but it has been suggested that they function in different protection systems. Expression of
TIL was induced during treatment of heat shock and cold acclimation in wheat and Arabidopsis (
Charron et al. 2002). Additionally, the quantity of Arabidopsis
TIL1 was significantly increased upon cold acclimation in Arabidopsis leaf (
Kawamura and Uemura 2003).
In this study, 67 genes responding to biotic or abiotic stress were selected for screening for differential expression in ‘Campbell Early’ and ‘Muscat Bailey A’ by low temperature treatment. Most of genes tested in this study showed differential expression patterns in transcription level in response to cold stress in grapevines.
Analysis of the differential expression of defense related genes in two grapevines could provide useful resources to breeding programs for development of low temperature tolerant grapevines. A number of valuable genes specifically expressed in cold tolerant grapevines should be selected through further studies such as transcriptome analysis. Sequences of genes with specific expression to low temperature could be useful in development of molecular markers based on SNP/Indels in tolerant grapevine breeding programs.
ACKNOWLEDGEMENTS
This work was supported by a grant (PJ008224) from the Agricultural R&D Project, Rural Development Administration, Republic of Korea.
Fig. 1Quantitative real-time PCR analysis of up-regulated gene expression in ‘Campbell Early’ (left) and ‘MBA’ (right) in response to low temperature. Each bar indicates the SEs (n=3).
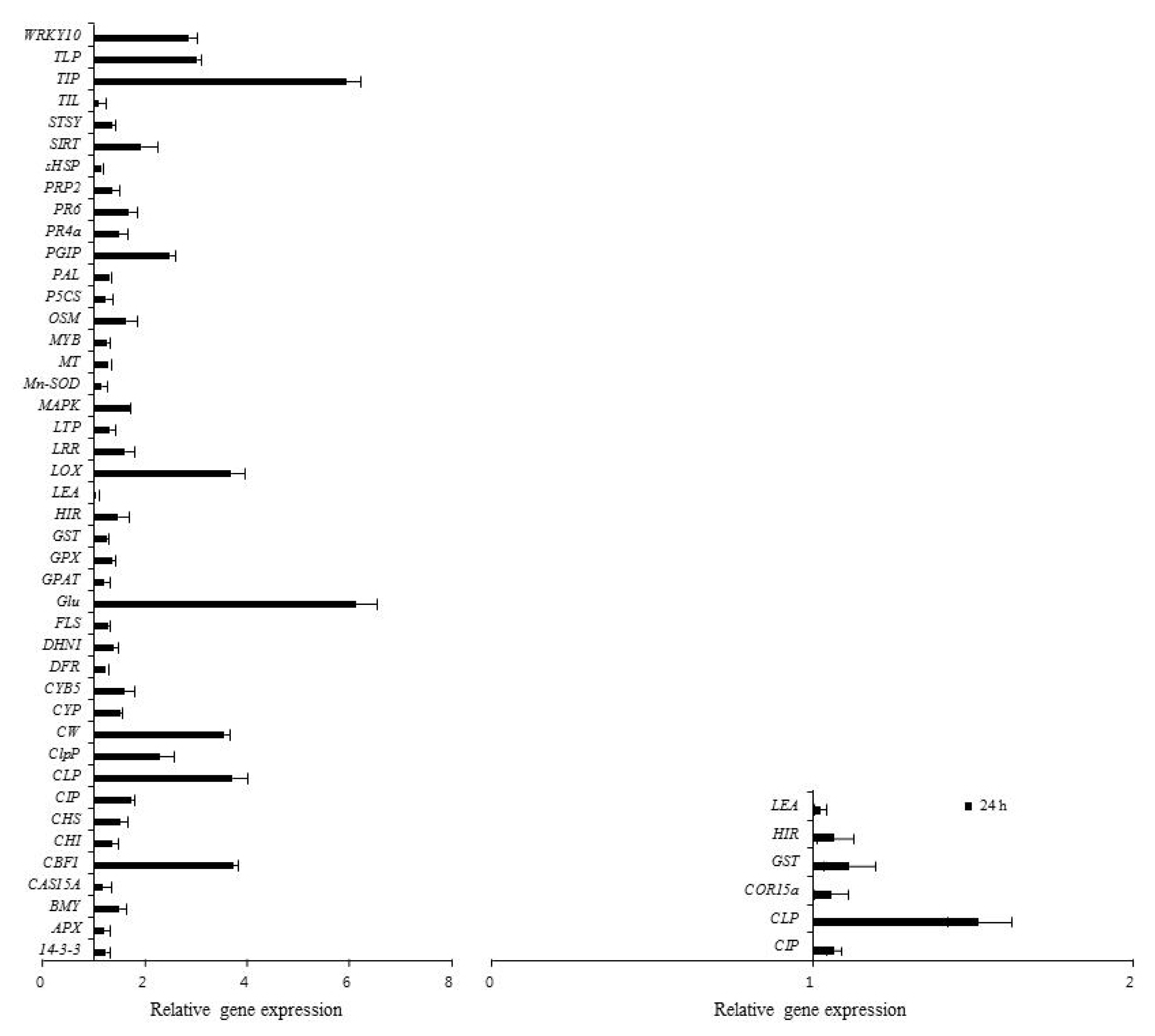
Fig. 2Quantitative real-time PCR analysis of differential gene expression in ‘Muscat Bailey A’ in response to low temperature. Each bar indicates the SEs (n=3).
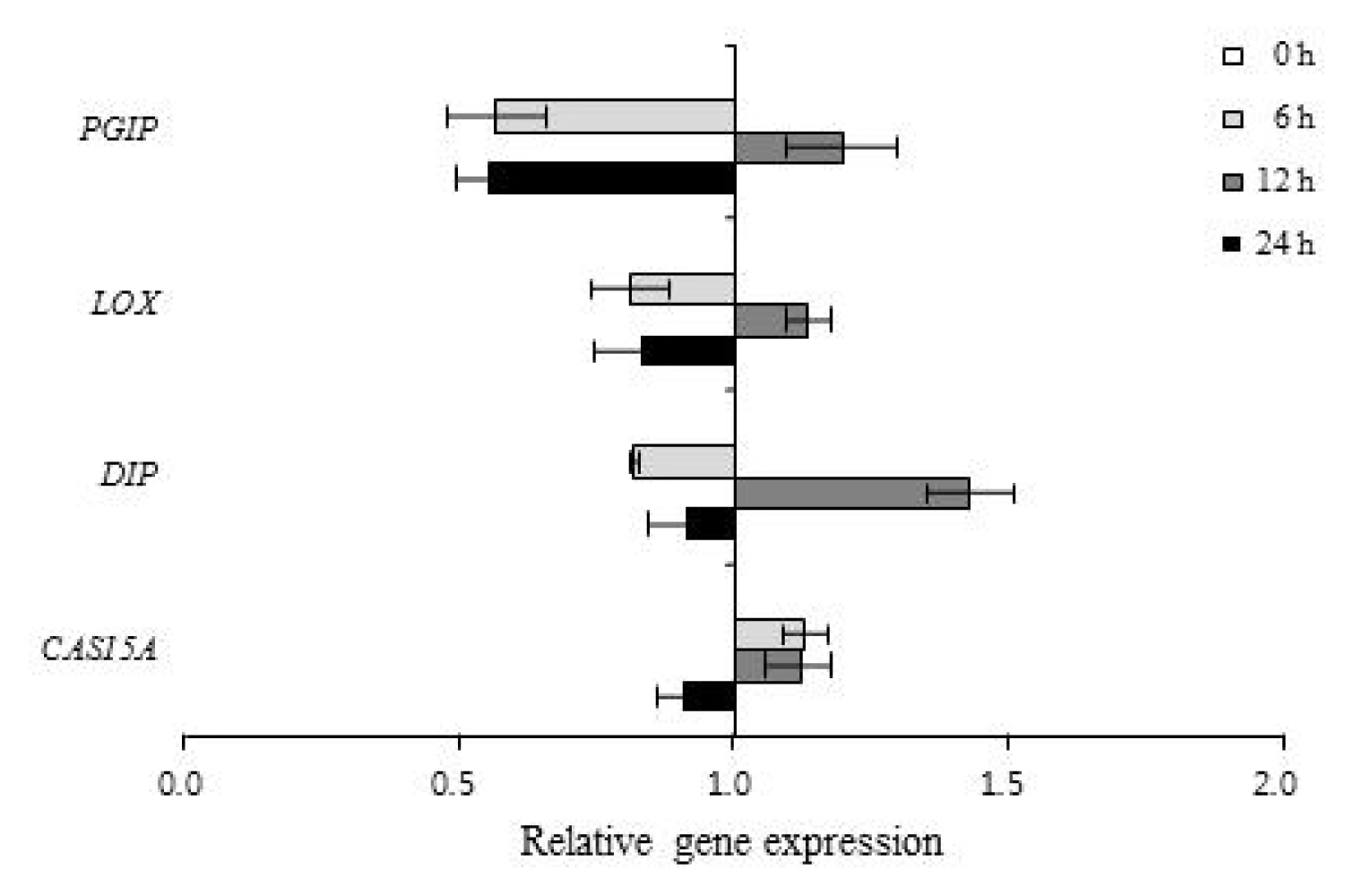
Fig. 3Quantitative real-time PCR analysis of down-regulated gene expression in ‘Campbell Early’ (left) and ‘Muscat Bailey A’ (right) in response to low temperature. Each bar indicates the SEs (n=3).
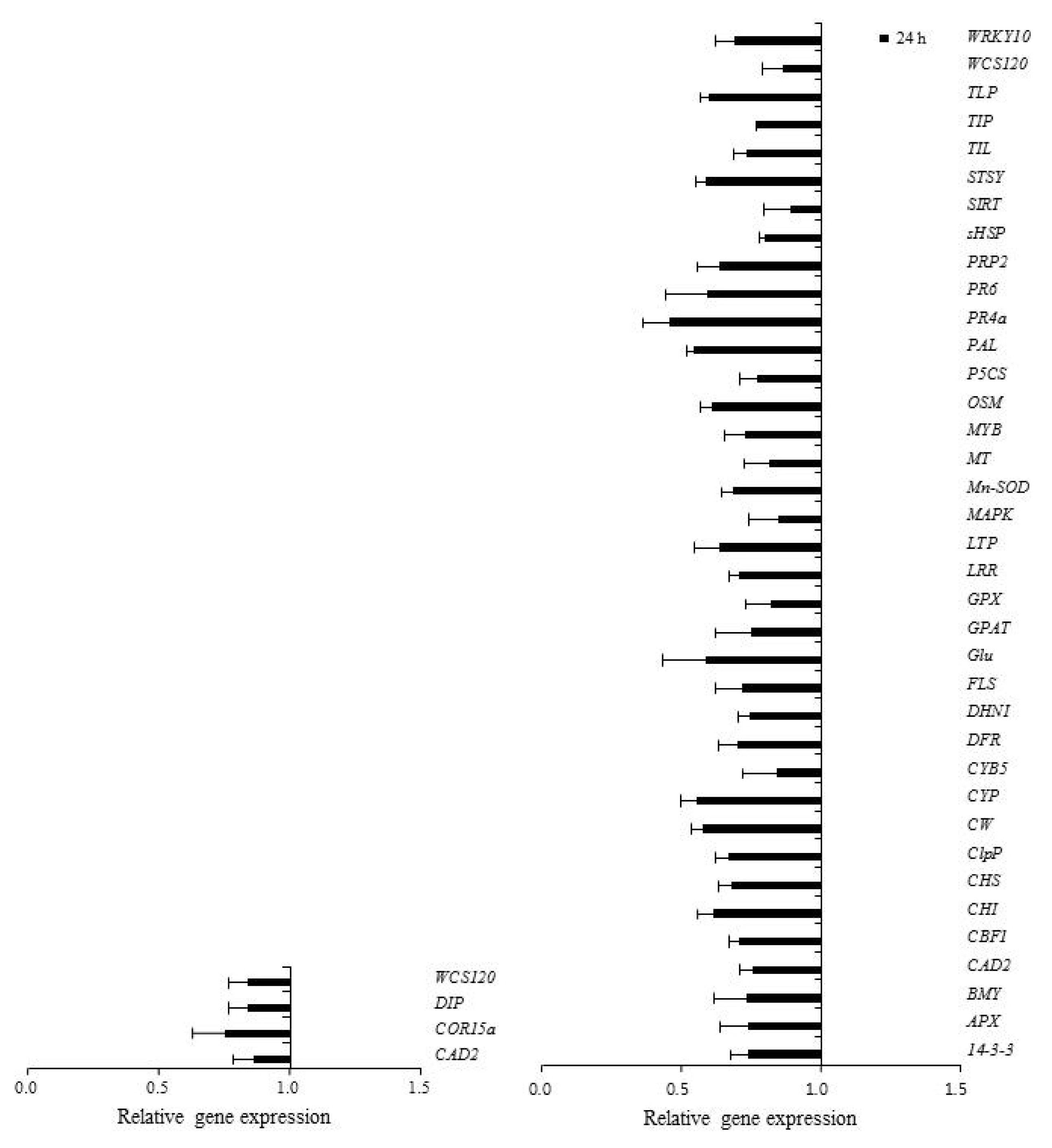
Fig. 4Venn diagrams of differentially expressed genes in dormant buds of ‘Campbell Early’ and ‘Muscat Bailey A’ grapevines in response to low temperature. A, up-regulated genes; B, down-regulated genes.
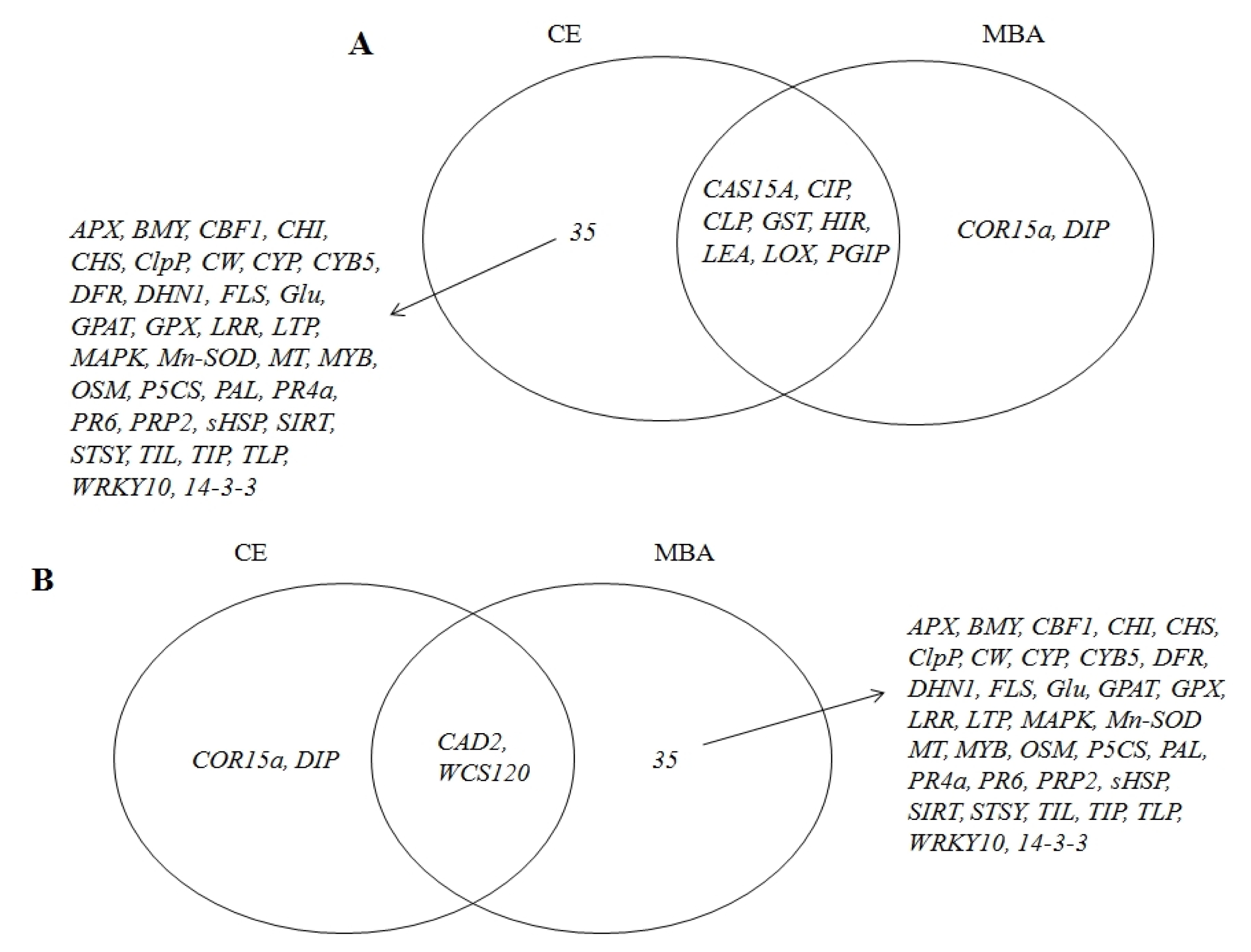
Fig. 5Semi-quantitative RT-PCR analysis to screen genes induced in dormant buds of ‘Campbell Early’ grapevines in response to low temperature. A, RT-PCR gel image; B, quantitative data for gel images from ImageJ analysis.
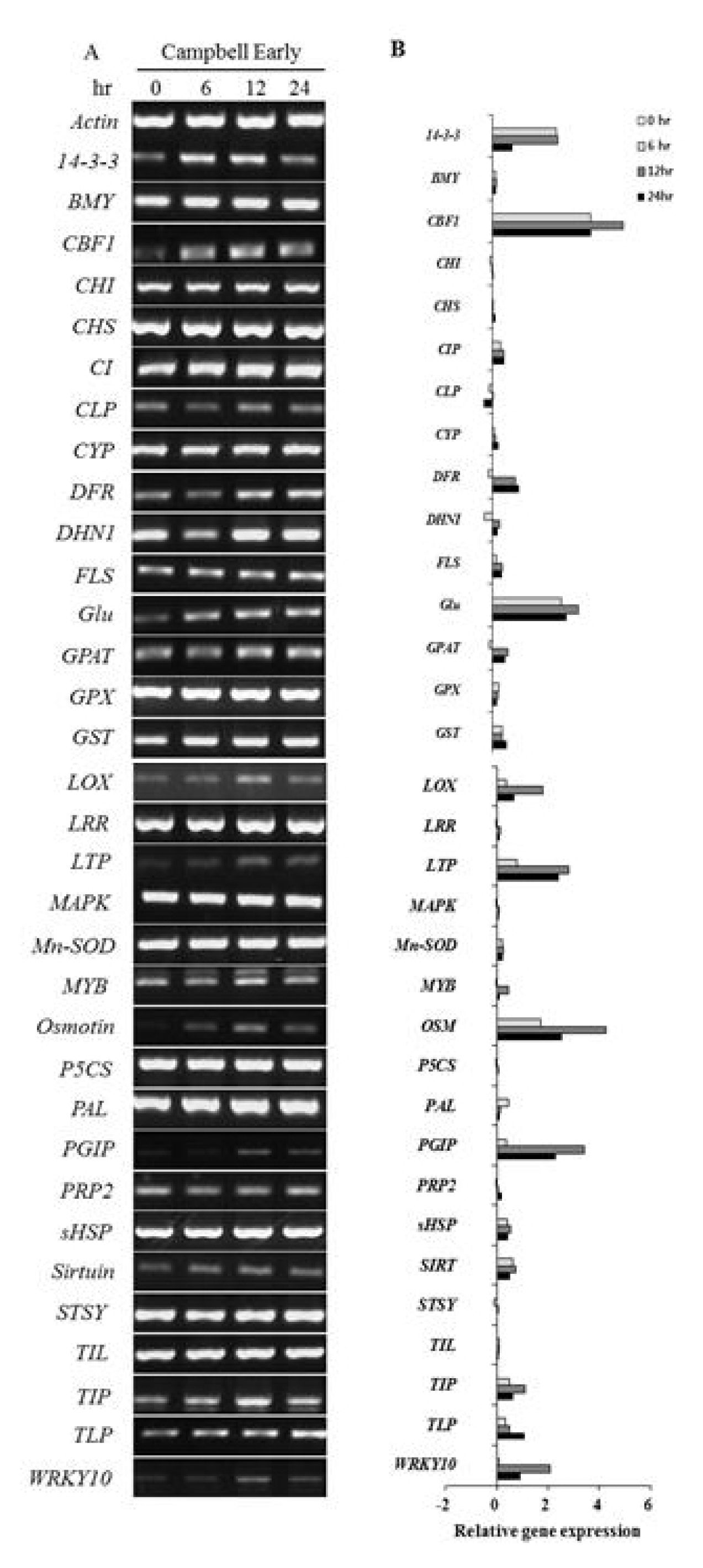
Fig. 6Semi-quantitative RT-PCR analysis to screen genes induced in dormant buds of ‘Muscat Bailey A’ grapevines in response to low temperature. A, RT-PCR gel image; B, quantitative data for gel images from ImageJ analysis.

Table 1Sequences of gene specific primers used for real-time PCR analysis.
Table 1
|
Name |
Primer sequences |
|
β-amylase (BMYl) |
5′-TCCACTCCCAGAATGGGTTAGA-3′ |
|
(XM0022745762) |
5′-TTTTAAAACCCGCTCCTTGTCA-3′ |
|
Ascorbate peroxidase (APX) |
5′-GGTCCGTTTGGGACAATGAA-3′ |
|
(EU280159.1) |
5′-CGGAAATTGCTCCTTGATCG-3′ |
|
ATP dependent Clp protease (ClpP) |
5′-TCACCCGACCAAATTGAAGC-3′ |
|
(XM00227683.1) |
5′-AACTCCCCGATCCTCAGAGC-3′ |
|
CBF like transcription factor (CBF1) |
5′-GGGTTTGCTCCTTACTCCACCT-3′ |
|
(AY390372.1) |
5′-CATCATCCCAGCTGAATCCTTT-3′ |
|
Cell wall protein (CW) |
5′-TTGACAAGAAGCAGCCTGAGTG-3′ |
|
(XM_002277798.1) |
5′-TGCTGCCATCATAGGGAGTGTA-3′ |
|
Chalcone isomerase (CHI) |
5′-TACACTGACGCAGAAGCCAAAG-3′ |
|
(XM002282072.2) |
5′-GACCCATCTTTGGAGAAGCTCA-3′ |
|
Chalcone synthesis (CHS) |
5′-AGTTCAAGCGCATGTGTGAAAA-3′ |
|
(EF192464.1) |
5′-CTTCAACCACCACCATGTCTTG-3′ |
|
Chitinase-like protein (CLP) |
5′-CCGATTTCTTCCAGACCTACCA-3′ |
|
(XM002269123.1) |
5′-CAAATCCGTGAGGCTGGTAAAC-3′ |
|
Cinnarmyl alcohol dehydrogenase2 (CAD2) |
5′-GGGCGTCCTCTCTCCATTTC-3′ |
|
(NM127743.3) |
5′-TCCCCAATGGTTCTTGATGG-3′ |
|
Cold acclimation protein WCS120 (WCS120) |
5′-CATGGACACGCTGGAGTGAT-3′ |
|
(M93342.2) |
5′-TGTCCCAGTGCCAGTCGTT-3′ |
|
Cold acclimation-specific (CAS15A) |
5′-CATGCTGGTGACCACAAAGG-3′ |
|
(L12462.1) |
5′-TTCCCCATGGTATCCCTCCT-3′ |
|
Cold induced protein (CIP) |
5′-AAGGGTGCAAAGCAAAGAGATG-3′ |
|
(XM002283501.1) |
5′-ATGCTGATGATCCATGGGAAGT-3′ |
|
Cold regulated protein 15a (COR15a) |
5′-ACCTCAACGAGGCCACAAAG-3′ |
|
(NM_129815.4) |
5′-CCGCAGCTTTCTCAGCTTCT-3′ |
|
Cytochrome B5 (CYB5) |
5′-AGAGGAGCATCCTGGTGGTG-3′ |
|
(XM002283578.1) |
5′-GCATGCTGTGGCCCAAAA-3′ |
|
Cytochrome p450 (CYP) |
5′-TGCAAGCAGTGGTGAAAGAGAC-3′ |
|
(CAB85635.1) |
5′-GACCTATAGCCCATGCGTTCAC-3′ |
|
Dehydrin1 (DHN1) |
5′-GGGAGAAGGAAGAAAGGGATGA-3′ |
|
(JF896552.1) |
5′-GTACTTGTGGCGCTGGTATGC-3′ |
|
Dihydroflavonol-4-reductase (DFR) |
5′-TGTCTCCAAGACACTGGCTGAG-3′ |
|
(JQ308621.1) |
5′-TCGGGGAAAGAGCAGTTATGAG-3′ |
|
Draught induced protein (R1G1A) (DIP) |
5′-CGTCGGTGACATGGATGAAA-3′ |
|
(AF503585.1) |
5′-TCCTGTTCACGGAAGCCAAT-3′ |
|
Endo-β-glucanase precursor (Glu) |
5′-GGGGTTATTTGGATCCCATCAT-3′ |
|
(AB601116.1) |
5′-CAGAAGCGGCGACTTATTGTCT-3′ |
|
Flavonol synthase (FLS) |
5′-TGCTGAGCAATGGGAAGTACAA-3′ |
|
(XM002285805.1) |
5′-GAGGTCCGATCATTGCCTTATG-3′ |
|
Glutathione peroxidase (GPX) |
5′-GAGCACAGGAACCTGGGAGTAA-3′ |
|
(XM003631370.1) |
5′-AGCACTATCGCCATTCACATCA-3′ |
|
Glutathion-S-transferase (GST) |
5′-TATAATGTGTGGGCAGCAAACG-3′ |
|
(AY156048) |
5′-CCAATGTCCAGAAAACCCAAAG-3′ |
|
Glycerol-3-phosphate acyltransferase (GPAT) |
5′-GAAGCTTCAGCAGGGTCACAAT-3′ |
|
(XM002276065.1) |
5′-GCTGGGTCTGCTTCTGTTTGAT-3′ |
|
Hypersensitive induced response protein (HIR) |
5′-TGCATCCTCAAAGGCTTCGT-3′ |
|
(AY159555.1) |
5′-TGCTGTCTCAGCCTGAAGGA-3′ |
|
Late embryogenesis abundant protein (LEA) |
5′-CTGACGTGGCAGAAGCAATG-3′ |
|
(NM129540.4) |
5′-TCGGACGCCATTTTCTTAGC-3′ |
|
Lipid transfer protein (LTP) |
5′-TGGAAGCAACCATAACATGTGG-3′ |
|
(LOC100256718) |
5′-TTGAGGCTCTTAATCCCACTGC-3′ |
|
Lipoxygenase (LOX) |
5′-AACCTTGCGAGGTAATGGTCAA-3′ |
|
(XM_002285538.2) |
5′-TACCACCAAGTACCGGTCGAGT-3′ |
|
Leucine-rich repeats (LRR) |
5′-GCCGATTTGGATCTCTCTCTGA-3′ |
|
(XM002285517.2) |
5′-GTATGCTCACCGCCGAGTTAAT-3′ |
|
Manganese superoxide dismutase (Mn-SOD) |
5′-GTTCATGAGGGAGGTGGTGAAC-3′ |
|
(EU280161.1) |
5′-TCTATAGCCCAACCCAGTGAGC-3′ |
|
Mitogen-activated protein kinase (MAPK) |
5′-CCCTCCTGATGAGGATTCATTG-3′ |
|
(AY395740.1) |
5′-ATTCGAACTCCATCCCTGTTGA-3′ |
|
MYB Transcripton factor (MYB) |
5′-CACAAAGTCCATCTCCCCAAAC-3′ |
|
(XM_002265978.1) |
5′-TCTCCATTTCCTCCTGAACTCG-3′ |
|
O-methyltransferase (MT) |
5′-CCAGACACCATCCACAACCA-3′ |
|
(JF808016.1) |
5′-GCAGCAAGGAAGCCAGAATG-3′ |
|
Osmotin like protein (OSM) |
5′-ACTGCAACTTCGATGCGTCA-3′ |
|
(Y10992.1) |
5′-TGCGAATTCGGCTAAGGTGT-3′ |
|
Pathogen related protein 4a (PR4a) |
5′-GCTGCCCAGAGCGCTAGTAA-3′ |
|
(AF061329.1) |
5′-TCCCAAGTGGAGCAGTAGGC-3′ |
|
Pathogen related protein 6 (PR6) |
5′-CTGGTGGGAGTTCAGGGAGA-3′ |
|
(AY156047.1) |
5′-CCAAACACGGACCCTAGTGC-3′ |
|
Phenylalanine ammonia lyase (PAL) |
5′-TGAACAATGGCGAAAGTGAGAA-3′ |
|
(X75967.1) |
5′-TCTCTTGCGCTCTCAACCTCTT-3′ |
|
Polygalacturonase-inhibiting protein (PGIP) |
5′-GTCTTACTCGCCACTCGTCCAT-3′ |
|
(AF305093.1) |
5′-GTTGGGATTCCACGAAGCTAGA-3′ |
|
Proline rich protein2 (PRP2) |
5′-CCACCTAAGATTAAACCGCGACT-3′ |
|
(XR078193.2) |
5′-CGAGGAATCGGAGGAAGCTTAT-3′ |
|
Pyrroline-5-carboxylate synthase (P5CS) |
5′-AGCCAATGCACTGGAAACAAAT-3′ |
|
(AJ005686.1) |
5′-CAGGCTTTAGAACCAGCCTTGA-3′ |
|
Regulator of cellular molecules (14-3-3) |
5′-TCAGGATATTGCAAACGCAGAA-3′ |
|
(FJ824803.1) |
5′-AAATGCCTGTTTCGCAAGGTTA-3′ |
|
Sirtuin (SIRT) |
5′-TGATGGCCTCCATCTTCGTT-3′ |
|
(JN252254.1) |
5′-CACCTCAAAATCCCGCAAGT-3′ |
|
Small heat shock protein (sHSP) |
5′-CATCTCGAGGTATGGGAACTGG-3′ |
|
(XM003634002.1) |
5′-GTGTTCTGCTCCACGCTTACCT-3′ |
|
Stilbene (STSY) |
5′-GGTGCCATTGCAGGAAACTTAC-3′ |
|
(X76892.1) |
5′-CAAGTGGGTCAAAAGCCTGAGT-3′ |
|
Temperature induced lipocalin (TIL) |
5′-TCAACGAGACTTGGAGTGATGG-3′ |
|
(DQ222993.1) |
5′-AACAGGGATAATGGGGAGGAAA-3′ |
|
Thaumatin-like protein (TLP) |
5′-TTCGCACTTAACCAATTCAGCA-3′ |
|
(XM002282928.2) |
5′-TGCACCCATTGGAAGTAGGATT-3′ |
|
Tonoplast intrinsic protein (TIP) |
5′-GGCACCCTAATTTTTGTGTTCG-3′ |
|
(XM003632211.1) |
5′-ACACTGCCACAAACAGTCCAAG-3′ |
|
WRKY transcription facter 10 (WRKY10) |
5′-CAGGTGTTCAGTTGATGGATGC-3′ |
|
(XM002279371.2) |
5′-AAGGGCTTTCGTGGTTATGGAT-3′ |
|
β-actin |
5′-ACGAGAAATCGTGAGGGATG-3′ |
|
(AB372563.1) |
5′-ATTCTGCCTTTGCAATCCAC-3′ |
References
- Ahn SY, Kim SH, Choi SJ, Yun HK. 2012. Characteristics of cold hardiness and growth in grapevines grown in rain shelter type cultivation system in the vineyard. Kor J Hort Sci Technol. 30: 626-634.
- Caruso A, Morabito D, Delmotte F, Kahlem G, Carpin S. 2004. Dehydrin induction during drought and osmotic stress in Populus. Plant Physiol Biochem. 40: 1033-1042.
- Charron JBF, Breton G, Badawi M, Sarhan F. 2002. Molecular and structural analyses of a novel temperature stress-induced lipocalin from wheat and Arabidopsis. FEBS Lett. 517: 129-132.
- Chi WT, Fung RW, Liu HC, Hsu CC, Charng YY. 2009. Temperature induced lipocalin is required for basal and acquired thermotolerance in Arabidopsis. Plant Cell Environ. 32: 917-927.
- Close TJ. 1997. Dehydrins: A commonality in the response of plants to dehydration and low temperature. Physiol Plant. 100: 291-296.
- Cook D, Fowler S, Fiehn O, Thomashow MF. 2004. A prominent role for the CBF cold response pathway in configuring the low temperature metabolome of Arabidopsis. Proc Natl Acad Sci (USA). 101: 15243-15248.
- Ferrari S, Vairo D, Ausubel FM, Cervone F, De Lorenzo G. 2003. Tandemly duplicated arabidopsis genes that encode polygalacturonase-inhibiting proteins are regulated coordinately by different signal transduction pathways in response to fungal infection. Plant Cell. 15: 93-106.
- Gilmour SJ, Artus NN, Thomashow MF. 1992. cDNA sequence analysis and expression of two cold-regulated genes of Arabidopsis thaliana. Plant Mol Biol. 18: 13-21.
- Guo W, Ward RW, Thomashow MF. 1992. Characterization of a cold-regulated wheat gene related to Arabidopsis Cor47. Plant Physiol. 100: 915-922.
- Hara M, Fujinaga M, Kuboi T. 2004. Radical scavenging activity and oxidative modification of citrus dehydrin. Plant Physiol Biochem. 42: 657-662.
- Hasegawa PM, Bressan RA, Zhu JK, Bohnert HJ. 2000. Plant cellular and molecular responses to high salinity. Annu Rev Plant Mol Plant Physiol. 51: 463-499.
- Hon WC, Griffith M, Chong P, Yang DCS. 1994. Extraction and isolation of antifreeze proteins from winter rye (Secale cereale L.) leaves. Plant Physiol. 104: 971-980.
- Houde M, Daniel C, Lachapelle M, Allard F, Laliberte S, Sarhan F. 1995. Immunolocalization of freezing-tolerance associated proteins in the cytoplasm and nucleoplasm of wheat crown tissues. Plant J. 8: 583-593.
- Hughes MA, Dunn MA. 1990. The effect of temperature on plant growth and development. Biotechnol Genet Eng Rev. 8: 161-187.
- Ishitani M, Xiong L, Stevenson B, Zhu JK. 1997. Genetic analysis of osmotic and cold stress signal transduction in Arabidopsis: interactions and convergence of abscisic acid-dependent and abscisic acid-independent pathways. Plant Cell. 9: 1935-1949.
- Kaplan F, Guy CL. 2004. β-Amylase induction and the protective role of maltose during temperature shock. Plant Physiol. 135: 1674-1684.
- Kawamura Y, Uemura M. 2003. Mass spectrometric approach for identifying putative plasma membrane proteins of Arabidopsis leaves associated with cold acclimation. Plant J. 36: 141-154.
- Kotak S, Larkindale J, Lee U, von Koskull-Doring P, Vierling E, Scharf KD. 2007. Complexity of the heat stress response in plants. Curr Opin Plant Biol. 10: 310-316.
- Lang V, Palva ET. 1992. The expression of a rab-related gene, rab18, is induced by abscisic acid during the cold acclimation process of Arabidopsis thaliana (L.) Heynh. Plant Mol Biol. 20: 951-962.
- Larkindale J, Hall JD, Knight MR, Vierling E. 2005. Heat stress phenotypes of Arabidopsis mutants implicate multiple signaling pathways in the acquisition of thermotolerance. Plant Physiol. 138: 882-888.
- Leyva A, Jarillo JA, Salinas J, Martinez-Zapater JM. 1995. Low temperature induces the accumulation of phenylalanine ammonialyase and chalcone synthase mRNAs of Arabidopsis thaliana in a light-dependent manner. Plant Physiol. 108: 39-46.
- Monroy AF, Dhindsa RS. 1995. Low-temperature signal transduction – Induction of cold acclimation-specific genes of alfalfa by calcium at 25C. Plant Cell. 7: 321-331.
- Monroy AF, Sarhan F, Dhindsa RS. 1993. Cold-induced changes in freezing tolerance, protein-phosphorylation, and gene-expression – Evidence for a role of calcium. Plant Physiol. 102: 1227-1235.
- Nikolai KC, Ryozo I, Yaroslav B. 2008. Differential expression of two winter wheat alpha-tubulin genes during cold acclimation. Cell Biol Int. 32: 574-578.
- Sanghera GS, Wani SH, Hussain W, Singh NB. 2011. Engineering cold stress tolerance in crop plants. Curr Genomics. 12: 30-43.
- Sarhan F, Ouellet F, Vazquez-Tello A. 1997. The wheat wcs120 gene family. A useful model to understand the molecular genetics of freezing tolerance in cereals. Physiol Plant. 101: 439-445.
- Seki M, Narusaka M, Abe H, Kasuga M, Yamaguchi-Shinozaki K, Carninci P, Hayashizaki Y, Shinozaki K. 2001. Monitoring the expression pattern of 1300 Arabidopsis genes under drought and cold stresses by using a full-length cDNA microarray. Plant Cell. 13: 61-72.
- Seki M, Narusaka M, Ishida J, Nanjo T, Fujita M, Oono Y, Kamiya A, Nakajima M, Enju A, Sakurai T, Satou M, Akiyama K, Taji T, Yamaguchi-Shinozaki K, Carninci P, Kawai J, Hayashizaki Y, Shinozaki K. 2002. Monitoring the expression profiles of 7000 Arabidopsis genes under drought, cold and high salinity stresses using a full-length cDNA microarray. Plant J. 31: 279-292.
- Sreekantan L, Mathiason K, Grimplet J, Schlauch K, Dickerson JA, Fennell AY. 2010. Differential floral development and gene expression in grapevines during long and short photoperiods suggests a role for floral genes in dormancy transitioning. Plant Mol Biol. 73: 191-205.
- Thomashow MF. 1999. Plant cold acclimation: freezing tolerance genes and regulatory mechanisms. Annu Rev Plant Physiol. 50: 571-599.
- Uemura M, Steponkus PL. 1997. Effect of cold acclimation on membrane lipid composition and freeze induced membrane destabilization. pp. 171-179. Li P, editor. Plant Cold Hardiness, Molecular Biology, Biochemistry and Physiology. Plenum Publishing Corp. New York:
- Wang W, Vinocur B, Shoseyov BO, Altman A. 2004. Role of plant heat-shock proteins and molecular chaperones in the abiotic stress response. Trends Plant Sci. 9: 244-252.
- Wu J, Zhang Y, Zhang H, Huang H, Folta KM, Lu J. 2010. Whole genome wide expression profiles of Vitis amurensis grape responding to downy mildew by using Solexa sequencing technology. BMC Plant Biol. 10: 234
- Xiong L, Zhu J. 2001. Abiotic stress signal transduction in plants: Molecular and genetic perspectives. Physiol Plant. 112: 152-166.
- Yadav SK. 2010. Cold stress tolerance mechanisms in plants. Agron Sustain Dev. 30: 515-527.
- Yang G, Zhou R, Tang T, Shi S. 2008. Simple and efficient isolation of high-quality total RNA from Hibiscus tiliaceus, a mangrove associate and its relatives. Prep Biochem Biotechnol. 38: 257-264.
- Yao CL, Conway WS, Ren RH, Smith D, Ross GS, Sams CE. 1999. Gene encoding polygalacturonase inhibitor in apple fruit is developmentally regulated and activated by wounding and fungal infection. Plant Mol Biol. 39: 1231-1241.
- Zhang CK, Lang P, Dane F, Ebel RC, Singh NK, Locy RD, Dozier WA. 2005. Cold acclimation induced genes of trifoliate orange (Poncirus trifoliata). Plant Cell Rep. 23: 764-749.
- Zhu J, Dong CH, Zhu JK. 2007. Interplay between cold-responsive gene regulation, metabolism and RNA processing during plant cold acclimation. Curr Opin Plant Biol. 10: 290-295.