Abstract
The latest report on the draft genome of Brassica rapa sequence has been published. To elucidate the functions of these genes and to efficiently search for agriculturally useful genes, a Full-length cDNA Over-eXpressor (FOX) gene hunting system was used. The FOX library from Chinese cabbage was introduced into rice via Agrobacterium-mediated transformation. Approximately 1,150 FOX-rice lines were generated. Genomic PCR analysis indicated that the average length of FL-cDNAs introduced into individual lines was 900~1,200 bp. Basic Local Alignment System Tool (BLAST) analysis of the FL-cDNA genes revealed that 35.5% have unknown function. Most of the randomly selected transgenic rice lines showed overexpression (92%) of these genes relative to the wild-type Gopum. Moreover, 94% of the 850 transgenic rice lines were moderately tolerant (slightly yellow) to cold and 9 lines were tolerant (seedlings were light green). Morphological evaluation of the transgenic rice lines showed minimal phenotypic alteration (12%). Approximately 25.1% and 22% of the plants were significantly ahead in the days to heading and had elevated chlorophyll content, respectively. Other agronomic traits such as filled grains, number of tiller, panicle length, and culm and plant height were relatively less variable among the transgenic lines. These results provide a resource for defining genes that are associated with tolerance in transgenic rice lines.
-
Key words: FOX-hunting system, Full-length cDNA, Abiotic stress, Transgenic plant, Rice
INTRODUCTION
Chinese cabbage (
Brassica rapa subsp. pekinensis) is one of the most important
Brassica species vegetables found in Asian countries. The draft genome sequence of
B. rapa has been recently published (The
Brassica rapa Genome Sequencing Project Consortium 2011). The Chinese cabbage genome contains 41,174 protein-coding regions, whereas 27,379 genes are currently believed to exist in the
Arabidopsis genome (TAIR9 information:
http://www.arabidopsis.org/). Appropriate biological experiments are needed to uncover the functions of these genes.
To accelerate the identification and characterization of gene function, various resources, including
Arabidopsis and rice information, are publicly available (
Nakamura et al. 2007). The
Arabidopsis Information Resource (TAIR) maintains a database of genetic and molecular biology data for the model higher plant
Arabidopsis thaliana (
www.arabidopsis.org). Data available from TAIR includes the complete genome sequence along with gene structure, gene product information, metabolism, gene expression, DNA and seed stocks, genome maps, genetic and physical markers, publications, and information about the
Arabidopsis research community. Along with this, infrastructure for rice bioin-formatics (reviewed in
Sasaki et al. 2005), including integrated databases such as RAP-DB (
Ohyanagi et al. 2006;
Rice Annotation Project 2007) and the TIGR Rice Genome Annotation Database (
Ouyang et al. 2007), BGI- RIS (
Zhao et al. 2004), and MOsDB (
Karlowski et al. 2003), is also publicly available. The available information provides an important resource for the analysis of
Brassica rapa genes.
For large-scale identification of gene function, several systems and tools have been developed to generate loss- of-function mutants (
Hirochika et al. 2004;
An et al. 2005). These loss-of-function resources and databases are very important for tagging and hunting non-redundant genes in the rice genome. However, 29% of rice genes are estimated to exist within clustered and redundant gene families (
IRGSP 2005). Therefore, it is difficult to find loss-of- function phenotypes for these duplicated genes. Moreover, when knockouts of genes confer embryonic lethality or severe developmental defects upon plants, it is impossible to the phenotypes associated with mutation or deletion of these genes (
Nakamura et al. 2007). To solve this problem, FL-cDNA clones (
Seki et al. 2002;
Ichikawa et al. 2006) were recently developed as an alternative gain-of-function approach for the systematic elucidation of gene functions in
Arabidopsis; the system is the so-called Full-length cDNA Over-eXpressor gene hunting system (FOX hunting system). FOX uses ectopic overexpression of a single or limited numbers of FL-cDNA(s) in individual transgenic plants. It generates large numbers of dominant mutations, enabling comprehensive characterization of novel and important traits and identification of the causal genes.
Studies on genetic transformation of plants have revealed an unexpected finding; the loss of mRNA encoded by either the introduced transgene or the transgene and endogenous homologous gene(s) (
Vaucheret and Fagard 2001). These gene-silencing phenomena are believed to be due to methylation induced premeiotically (MIP), repeat- induced point mutation (RIP), RNA interference (RNAi), and repeat-induced gene silencing (RIGS) (
Fagard and Vaucheret 2000;
Marathe et al. 2000). In order to avoid this problem during our study of gene function, FL-cDNAs genes from Chinese cabbage were transformed into the rice cultivar Gopum.
In this study, clones with full-length cDNA from Chinese cabbage were sequenced, and transgenic rice plants were subsequently generated via
Agrobacterium- mediated transformation. The function of each clone was classified according to the annotation function based on BLAST analysis (National Center for Biotechnology Information;
www.ncbi.nlm.gov). Moreover, the FOX rice lines generated were evaluated in the fields to determine their morpho-agronomic characteristics and phenotypic traits, as well as their responses to cold and salinity.
MATERIALS AND METHODS
Plant materials and field trials
The rice variety Gopum was used as the wild-type strain in the generation of transgenic rice. Regenerated plants were transplanted in soil consisting of 50% compost and 50% earth soil and acclimatized for 2 weeks in a greenhouse. Young leaves were collected for genomic DNA analysis using
pBigs_sfl1 and
hpt primers (GenBank Accession No. AF234296). Subsequently, plants that were confirmed to carry the gene insert were harvested, and the T
1 seeds were used in the succeeding generation in the field following the protocol of
Sun et al. (2011). The varieties were sown in a seedling-growing tray and placed in the greenhouse for 3 weeks in May, 2011. Grown seedlings were transplanted in the experimental farm spaced at 30×15 cm with one seedling per hill and arranged in randomized complete block design replicated three times. The cultivation and management were performed according to the standards of the experimental farm in Chungbuk National University.
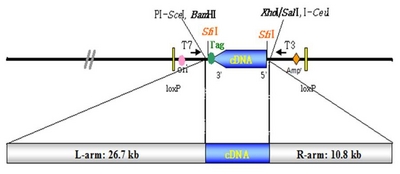
Brassica rapa clones with full-length cDNA were ligated into the
pBigs_sfiI vector. The recombinant vector carrying these clones was constructed under the control of the
pUBI-1 promoter and NOS terminator, as shown in
Fig. 1. Each clone was grown in Luria-Bertani medium containing 50 mg/L kanamycin sulfate and incubated at 28°C on a shaker for 24~36 h. Plasmid DNA, extracted according to the Qiagen plasmid kit (Cat. No. 12125) protocol, was used in the sequence analysis.
Generation of transgenic rice
Transgenic rice was generated following the protocol of
Toki et al. (2006), with some modifications (
Lee et al. 2011). Briefly, dehulled mature seeds of the
japonica rice cultivar Gopum were sterilized with 70% ethanol for 5 min. These seeds were further sterilized by strong shaking in 30–35 mL of 2.5% sodium hypochlorite containing one drop of Tween 20 for 15 min. The sterilized seeds were washed 20 times in distilled water, dried in the clean bench for 30 min, and then inoculated on embryogenic callus induction (2N6) medium (
Table 1). This set up was incubated in the dark at 32°C for 7–10 days.
Agrobacterium strain EHA105 harboring binary Ti plasmid
pFLCIII (
Fig. 1) containing the full- length cDNA gene was cultured on AB medium containing 50 mg/L kanamycin sulfate solidified with 1.5% agar incubated for 3 days at 28°C in the dark.
Agrobacterium culture was scraped from the plates and suspended in AAM medium to yield an OD
600 of approximately 0.3. Pre-cultured seeds were immersed in the
Agrobacterium suspension by gently inverting the tube for 15 min, blotted dry with a sterilized filter paper to remove excess bacteria. The seeds were transferred onto a sterilized filter paper that had been moistened with 0.5 mL of AAM medium placed on 2N6-AS medium solidified with 0.4 % Gelrite. After 3 days of co-cultivation at 25°C in the dark, seeds were washed 5 times in sterilized water, and then washed once in sterilized water containing 500 mg/L carbenicillin to remove
Agrobacterium cells. The seeds were rapidly blotted dry on a sterilized filter paper and cultured on N6D (
Table 2) medium containing 50 mg/L hygromycin and 400 mg/L carbenicillin under continuous light at 32°C for 2 weeks. Proliferating calli arising from the scutellum were transferred to REIII (
Table 2) medium. Plantlets arising from the calli were transferred to HF medium to induce root formation.
DNA extraction and PCR analysis
Genomic DNA was extracted as described by
Cho et al. (2007) with some modifications. The relative purity and concentration of extracted DNA was estimated using NanoDrop-1000 (NanoDrop Technologies, Inc. USA). PCR analyses were performed using HPT-Fw (5′-GGA-TTTCGGCTCCAATGTCCTGACGGA-3′) and HPT-Rv (5′-CTTCTACACAGCCATCGGTCCAGA-3′) primers to check the introgression of hygromycin phosphotransferase gene (
hpt) and
pBigs_SfiI-Fw (5′-TATTCGGAGAGGG-TACGTATTTTTAC-3′) and
pBigs_sfiI-Rv (5′-GCA-ACAGGATTCAATCTTAAGAAACT-3′) to check the introduction of the full-length cDNA gene.
For DNA blot analysis, 10 μg of genomic DNA was extracted as mentioned above. A digoxigenin (DIG) probe was synthesized according to the manufacturer’s instructions (PCR DIG Probe Synthesis Kit; Roche Molecular Biochemicals, USA). The template for the probe synthesis was from the vector containing the full-length cDNA clone. The forward/ reverse primers used for the synthesis of the probe were 5′-GATTAAGCATCCAAATGTTGTTCA-3′ and 5′-CCTACGAGCCTCATCTTCT-3′. The PCR amplification was initiated with a denaturation at 95°C for 2 min, followed by 30 cycles of 95°C for 30 s, 56°C for 30 s, and 72°C for 60 s, and the final elongation step at 72°C for 7 min. Ten micrograms of genomic DNA was digested overnight at 37°C with the restriction enzymes BamHI and EcoRI. The digested DNA was subjected to electrophoresis in a 0.8% agarose gel and transferred to a Hybond-N+ membrane (Amersham Biosciences, USA), according to the manufacturer’s instructions. The membrane was pre- hybridized at 42°C in a DIG East Hyb buffer for 1 h. A DIG-labeled probe was then added (2 μL of probe/mL of buffer) and hybridization was carried out overnight at 42°C. The membrane was washed twice with a low- stringency washing solution (2× SSC, 0.1% SDS) at 25°C for 5 min, and then washed twice with a high-stringency solution (0.1× SSC, 0.5% SDS) at 68°C for 15 min. The DIG-labeled DNA was detected according to the manufacturer’s instructions (DIG Nucleic Acid Detection Kit; Roche Molecular Biochemicals, USA).
Cold, salinity, and drought tolerance screen
Cold evaluation was conducted at the Chuncheon Experimental Station of the Rural Development Administration, Korea. About 50 seeds were directly seeded into germination trays (60 × 30 × 3.5 cm) with a soil-free medium of steam-sterilized sand with peat compost and vermiculite and allowed to grow for 7 weeks before cold treatment at a water temperature of approximately 13°C, and an air temperature of 30–35°C.
For the salinity and drought tolerance test, styrofoam seedling floats were fabricated as described by
Manigbas et al. (2011). Holes were drilled at the surface of the styrofoam with a nylon net supporting the bottom. Two pre-germinated seeds were placed on each hole. Seedling floats were placed on each tray comprising a replication of two, with 20 seedlings per entry. The growing plants were supplied with Murashige and Skoog (MS) solution at 4.4 gram per liter to maintain good growth and healthy seedlings throughout the experiment. After 2 weeks, solutions of 130 mM NaCl and 20% PEG were used to induce salinity and drought, respectively on rice seedlings; this solution was changed after 8 days. The lines were scored after 2 weeks, or when the susceptible check showed high susceptibility by using the standard evaluation system for rice (
IRRI-SES 2007).
Total RNA was extracted from the transgenic lines and wild-type Gopum plant by using the RNeasy Plant Mini Kit (QIAGEN, Maryland, USA), according to the manufacturer’s instructions. This process was repeated in all experiments requiring RNA extraction. The relative purity and concentration of extracted RNA was estimated using NanoDrop-1000 spectrophotometer (NanoDrop Technologies, Inc. USA), and stored in a −80°C freezer. Total RNAs were cleaned using DNaseI, and first-strand cDNA synthesis was performed by reverse transcription of mRNA using Oligo (dT)20 primer and SuperScript™ III Reverse Transcriptase.
The nucleotide sequence of each of the randomly selected clones was used to design primer pairs (
Table 3). The specific sequences of the primer pairs were used in a semi-quantitative reverse transcription PCR (RT-PCR). Actin primers were used as loading controls and used as internal controls for normalization of the quantitative RT-PCR reaction.
Data requiring statistical analysis were computed using the Statistix version 8 (
www.statistix.com). Significant P values were analyzed by using the two-sided Dunnett’s multiple comparison, with the wild-type Gopum as control by using the same software. The frequency distribution of various agronomic traits and sequences was determined using Microsoft Excel 2007.
All the inserted sequences were checked using the BLAST program in NCBI sequence database and another program in the GenBank/EMBL/DDBJ sequence database. The open reading frame (ORF) and conserved domain were generated using the BLASTN program (
http://www.ncbi.nlm.nih.gov/). Sequence alignment, ORF translation, molecular weight calculation of the predicted proteins, and structural analysis of the deduced proteins were carried out by DNAStar’s Lasergene sequence analysis software (
www.dnastar.com).
RESULTS
Characterization of selected clones from FOX hunting system
Agrobacterium clones containing FL-cDNAs genes from
Brassica rapa were subjected to genomic PCR using forward and reverse primers (
pBigs_SfiI-Fw and
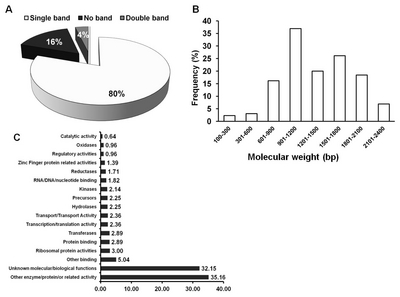
pBigs_ sfiI-Rv) derived from the flanking sequences. Of the 933 clones analyzed, 80% showed single bands, while 4% showed multiple fragments (
Fig. 2A). The remaining clones (16%) did not undergo fragment amplification. All the amplified single PCR fragments bearing the integrated FL-cDNA(s) from
B. rapa clones were used to generate their respective sequences. The sizes of the introduced FL-cDNAs were variable, ranging from 100 to 2,400 bp (
Fig. 2B). About 5% of the clones were smaller than 500 bp, whereas 95% ranged from 600 to 2,400 bp.
The Basic Alignment System Tool (BLAST) provided by the National Center for Biotechnology Information (
www.ncbi.nlm.nih.gov) was used to determine the functional annotation of each clone. The frequency distribution based on function is presented in
Fig. 2C. Variable gene function was observed from the viewpoint of molecular as well as biological importance. Most clones (32.15%) were characterized as having an unknown biological/molecular gene function. About 0.64% of clones appeared to possess functions related to catalytic activity, and 5.04% appeared to exhibit some binding activity. Transcription- and translation-related functions accounted for 2.36% of the clones. Moreover, 1.39% of the clones had homology with zinc finger-related proteins. A large number of other functions, each with a lower frequency than those reported above, collectively accounted for another 35.16% of the clones (
Fig. 2C). These genes provide important resources for studying their function in plants.
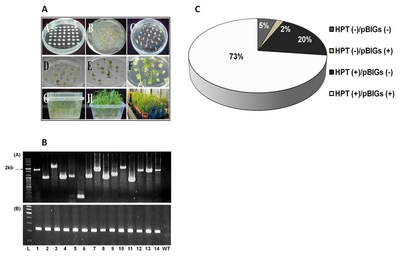
Using the protocol described by
Lee et al. (2011) and modified on the basis of that described by
Toki et al. (2006), transgenic rice plantlets were regenerated within a month starting from the aseptic culture of the mature seeds (
Fig. 3A). The plantlets were subjected to PCR analysis to verify the integration of FL-cDNAs. The genomic DNA PCR analysis is represented in
Fig. 3B. Most of the regenerated plants (T
0) (73%) contained the expected band sizes (
Fig. 3C), indicating the high efficiency of the transformation method. About 20% of the transgenic plants had
hpt gene, whereas 2% had only FL-cDNAs gene.
The confirmed FOX rice lines were advanced in the succeeding planting season from 2009 to 2011. So far, we have propagated transgenic rice up to the fourth generation (T4) (
Table 4). Most of the transgenic rice lines were in the T
1 generation that constituted 470 independent rice lines in 137 independent clones. In the T
2 generation, 408 lines with 88 gene inserts were produced. In the T
3 generation, 46 lines were generated. In summary, a total of 250 independent clones were inserted into the rice genome, leading to the generation of 1,150 independent transgenic lines (
Table 4). These transgenic rice lines are a valuable resource for future analyses designed to determine the functions of each gene.
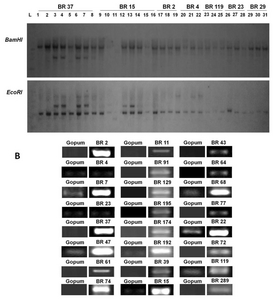
To examine the gene copy number of 31 randomly selected lines, we used the digoxigenin (DIG) system method (Roche Molecular Biochemicals, USA). Because of variability in the gene sequences, we used
hpt gene as probe. Our results showed that most of the transgenic lines have a single copy number (
Fig. 4A). However, in BR 37, which encodes CBL-interacting protein kinase 1, two gene copies were present in each of 4 lines (numbers 3, 4, 6, 7). Similarly, in BR 15, which encodes UDP-glucose-4- epimerase 1, two lines had two copies of the gene (numbers 12 and 13). Furthermore, in clone BR 4, two copies of the introduced gene (actin-depolymerizing factor 2) were observed. The rest of the lines harbored transgenes at single copy. The variability in copy number has been reported in previous studies (
Schrider and Hahn 2010). This may be due to the positional effect of the gene during transformation (
Gelvin 2003).
To examine whether the expression of transgenes was elevated at the molecular level, 24 transgenic lines encoding different gene inserts were randomly selected. Expression of these 24 genes was examined by semi- quantitative RT-PCR analysis with individual gene-specific primers (as shown in
Table 3) by using the young shoots of each rice seedling. mRNA expression was observed in all 24 genes with varying band intensities (
Fig. 4B). Strong mRNA expression was observed in BR 2 (cysteine protease 1), BR 7 (DEAD box RNA helicase RH15), BR 37 (CBL-interacting protein kinase 1), BR 47 (unknown gene function), BR 74 (haloacid dehalogenase- like hydrolase), BR 192 (glycosyltransferase), BR 15 (UDP-glucose-4- epimerase 1), BR 68 (unknown function), and BR 22 (hydrogen-transporting ATPase). Moreover, weak mRNA expression was observed in BR 4 and BR 23 lines encoding an actin-binding/depolymerizing factor 2 and endoplasmic reticulum auxin-binding protein 2 mRNA, respectively.
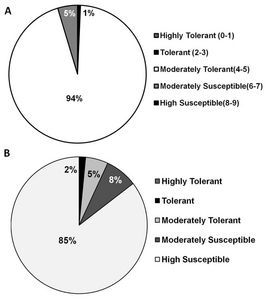
To determine the response of transgenic rice lines to cold, 840 independent lines comprising 250 genes were evaluated at the Chuncheon Experimental Station of the Rural Development Administration, Korea. Most of the transgenic rice lines (94%) were moderately tolerant (seedlings were light yellow) to cold and about 5% were moderately susceptible (seedlings were turning brown) (
Fig. 5A). Interestingly, 9 (1%) of the transgenic lines were tolerant (seedlings were light green) (
Table 1). Of the 9 tolerant lines, 4 encode genes with an unknown function (BR 5, BR 119, BR 522, BR 503). Other tolerant lines encoded UDP-galactose transporter 3 (BR 457), DEAD box RNA helicase RH 15 (BR 7), Arv1-like protein (BR 115), carbon-monooxide oxygenase (ACYB-1) (BR 39) and light harvesting complex PSII (BR 541). Conversely, we found 6 transgenic lines that were susceptible to cold stress, 4 of which encode an unknown gene function. The rest of the transgenic lines responded to the cold in a manner indistinguishable from wild-type Gopum.
In the salinity test, 85% of the transgenic rice lines were found to be highly susceptible (almost all plants were dead or dying) (
Fig. 5B). However, we found 7 tolerant transgenic lines (nearly normal growth and tillering) (
Table 1). These lines were BR 7 (unknown gene function), BR 37 (CBL-interacting protein kinase 1), BR 40 (Phosphatase 2C), BR 74 (Haloacid dehalogenase-like hydrolase), BR 91 (unknown gene function), BR 115 (BREB subfamily A-4 ERF/AP2 transcription factor), and BR 119 (unknown gene function). Moreover, 15 lines were shown to be moderately tolerant to salinity stress (their growth was nearly normal, but plants had some rolled leaves). These tolerant lines are an important resource for studying the mechanisms of resistance, and may eventually facilitate crop improvement in the future.
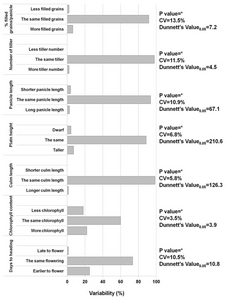
To determine the phenotype of the FOX rice lines, all developed lines were evaluated in the field arranged in an incomplete block design. As shown in
Fig. 6, the variability in the number of transgenic lines was significant in each of the agronomic traits measured. Indeed, 25.1% matured significantly earlier and 22% contained significantly higher chlorophyll content. Furthermore, 18% of the transgenic rice lines had lower chlorophyll contents. Other agronomic traits such as filled grains, number of tiller, panicle length, and culm length and plant height showed relatively low variability among lines. All morpho- agronomic traits are recorded. For example, the BR 4 line, which encodes actin-depolymerizing factor 2, exhibited a higher number of tillers. Similarly, BR 119, which encodes a gene with unknown function, gave rise to shorter plants.
The altered agronomic traits observed in the FOX lines may be caused by each gene. Further functional analysis is necessary to determine the function of each gene. Regardless of this, these results provide the basic information required to accelerate future functional studies.
DISCUSSION
A total of 1,150 transgenic rice lines with FL-length cDNA genes from Chinese cabbage were generated; these lines contained 250
Brassica rapa FL-cDNAs (
Table 4). The size distribution pattern and the average size of FL-cDNAs inserted into the genomes of 250 FOX-rice lines was 900 to 2,100 bp (
Fig. 2B). Analyses of individual gene inserts revealed that genes associated with tolerance to cold and salinity were functionally diverse, both at the molecular and biological levels (
Fig. 2C). Approximately 1/3 of the gene inserts have no known ascribed function. Furthermore, most of the randomly selected transgenic lines were present as single gene copies (
Fig. 3). We overexpressed a variety of transgenes in transgenic rice (
Fig. 4). We suggest that our approach will be useful for systematic genome-wide gain-of-function analyses of the
Brassica rapa genes that have been inserted into rice.
Phenotypic databases of rice properties have been developed by several groups. These include the International Rice Information System (IRIS) (
Wu et al. 2005), Oryzabase (
Kurata and Yamazaki 2006), Rice Mutant Database (RMD;
Zhang et al. 2006), Rice Tos17 Insertion Mutant Database (
Miyao et al. 2007), and Gramene (
http://www.gramene.org/). In addition to these useful databases, the transgenic rice lines harboring FL-cDNAs gene from
Brassica spp. are expected to provide valuable information for researchers to uncover functions of
Brassica genes. Appropriate biological or molecular methodologies are needed to reveal the function of each gene.
We found that 7.9% of the observed transgenic rice plants showed morpho-agronomically altered phenotypes (
Fig. 6). Among the 1,150 transgenic lines, 224 lines (26.4%) had a significantly altered number of days to heading. Similarly, 40% had a significant change in the chlorophyll content. Examples of these altered phenotypes are presented in
Fig. 7. This rate was much higher than that determined by using activation tagging of rice populations (
Jeong et al. 2002). However, for culm length, plant height, panicle length, number of tillers/hill, and percent filled spikelet/panicle, only 3.7% of transgenic lines were different compared to the wild type. During
Agrobacterium transformation, minimal weak growth of the regenerants was observed. This is consistent with a previous report (
Nakamura et al. 2007) and underscores the notion that transgenic regenerants often suffer from the effects of environmental factors such as humidity, temperature, macro- and micro-nutrient deficiencies or excesses, and microbes (pathogens), especially in the initial growth stage after transfer to soil. Alternatively, during tissue culture for selection and regeneration of transgenic plants, a variety of both epigenetic (e.g., DNA methylation, gene silencing, activation of retrotransposons and transposable elements) and genetic (e.g., nucleotide substitutions, deletions, insertions, rearrangements) alterations may occur in the plant genome (
Kaeppler et al. 2000;
Cheng et al. 2006;
Noro et al. 2007). These factors, i.e., abiotic and biotic factors and tissue culture-induced variations, may influence the growth conditions to some degree, and consequently, alter the phenotypes of transgenic plants. This may explain the increased rate of phenotype alteration in our transgenic lines (
Nakamura et al. 2007).
The response of transgenic rice lines to salinity and cold was tested on 900 independent lines. Eight hundred forty six (94%) of the lines were moderately tolerant to the cold stress (
Fig. 5A), and 9 lines (1%) were tolerant. This is not surprising since wild-type Gopum is moderately tolerant to cold stress (
Choi et al. 2007). However, most of the transgenic lines (85%) were highly susceptible to salinity stress (
Fig. 5B). Only 5% were moderately tolerant, and 2% were tolerant in the transgenic rice lines. In any case, our results have identified possibly agronomically important genes that should be characterized in future studies. Furthermore, manipulation of these genes may lead to improvements in the tolerance parameters of this economically important crop.
The development of full-length cDNA collections is an effective strategy for increasing the catalog of gene transcripts. These data serve as a valuable resource for describing gene expression profiles, and ultimately classify genes into families based on their functions (
Abe et al. 2011). Full-length cDNA can also serve as a powerful tool to facilitate genomic or other research (
Seki and Shinozaki 2009). Several techniques have been established to prepare enriched full-length cDNA libraries (
Seki et al. 1998). The utility of full-length cDNAs has been confirmed in various plants, including
Arabidopsis (
Yang et al. 2006;
Alexandrov et al. 2006), rice (
Ralph et al. 2008), poplar (
Jia et al. 2006), maize (
Alexandrov et al. 2009), tomato (
Town et al. 2006), and soybean (
Umezawa et al. 2008). A major advantage of this approach is that most of the clones contain the complete coding sequence, in addition to the 5′- and 3′-untranslated regions.
In this study, we have described the phenotypes and abiotic reactions of individual transgenic rice lines that carry gene inserts isolated from Chinese cabbage. Our results have thus provided a list of potentially economically important genes that can be further studied during future abiotic studies and functional analyses.
Supplementary Materials
ACKNOWLEDGEMENTS
This work was carried out with the support of Cooperative Research Program for Agriculture Science & Technology Development (Project No. PJ008529), Rural Development Administration and the research grant of the iPET, Ministry for Food, Agriculture, Forestry and Fisheries, Republic of Korea.
Fig. 1Schematic diagram of the binary Ti plasmid pFLCIII containing the full length cDNA from Chinese cabbage. pBigs vector was consisted of two different SfiI sites [SfiI(A) and SfiI(B)].
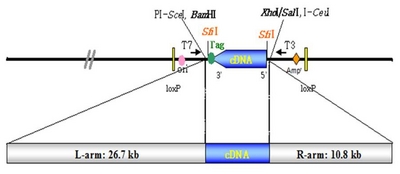
Fig. 2PCR amplification and BLAST analyses of 933 clones. (A) Pattern and numbers of cDNA-containing fragments. (B) frequency of gene according to the length (bp) of FOX cDNA library. (C) NCBI database annotation based on gene function.
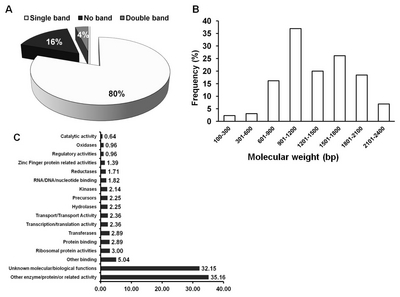
Fig. 3Development, PCR analysis and transformation efficiency in the generation of transgenic rice. (A) Development of selected 250 clones from FL-cDNA through Agrobacterium-mediated transformation (A) callus formation; (B) inoculation; (C) callus selection using hygromycin (D), (E) and (F) shoot formation and elongation; (G) and (H) root formation and elongation; (I) acclimatization; (B) Genomic PCR confirmation of inserted genes using T1 plants by pBigs SfiI (a) and HPT (b) primers in 1% agarose gel. L= Ladder,1 = BR2, 2=BR4, 3=BR7, 4=BR23, 5=BR47, 6=BR61, 7=BR74, 8=BR91, 9=BR129, 10=BR192, 11=BR195, 12=BR174, 13=BR15, 14=BR37, WT= wild type; (C) The efficiency of Agrobacterium-mediated transformation based on the genomic PCR analysis of 1,072 T0 generation rice lines using hpt and pBigs_SfiI primers.
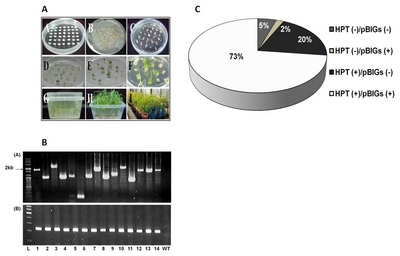
Fig. 4Southern blot analysis and expression pattern of randomly selected FOX rice lines. (A) hpt gene was used as probe using a non-isotope method (dioxiginin). BR 37 (BrCIPK1, CBL-interacting protein kinase 1); BR15 (BrUGE1, UDP-glucose-4-epimerase 1); BR2 (BrCP1, Cysteine protease 1); BR4 (Actin depolymerizing factor 2); BR119 (Unknown function); BR23 (ER auxin-binding protein 2); BR29 (Phospholipase C). (B) RT-PCR of selected rice lines using their respective primer pairs (
Supplemental Table 2. Rice actin was used as loading adjustments (data not shown). Gopum was the wild type.
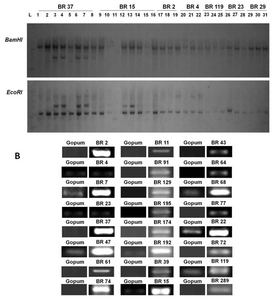
Fig. 5Response of FOX rice lines against cold and salinity (see materials and methods for details).
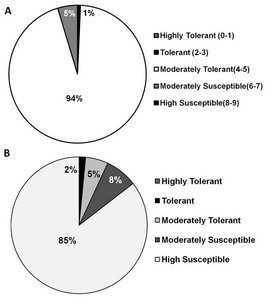
Fig. 6Morpho-agronomic variability of 1,150 transgenic rice lines. Each transgenic rice line was compared to the wild type Gopum to generate significant differences (positive/negative) by two-pair wise Dunnet’s.
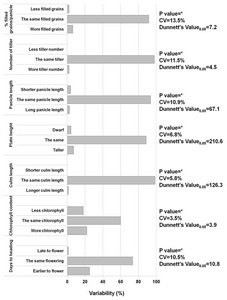
Fig. 7The field evaluation of transgenic rice. A to C, the field plots of the transgenic lines in the vegetative stage arranged in incomplete block design; D, high senescense observed in BR 73; E, mild senescence in BR61; F, observed dwarf in BR122; G, partial sterility observed in BR 452.

Table 1Identified transgenic rice with cold, salinity and drought tolerance.
Table 1
|
Clone |
Tolerance to NaCl stresses (130 mM) |
Tolerance to Cold |
Drought (20% PEG) |
Gene function (based on BLAST) |
|
BR5 |
− |
++ |
− |
Unknown function |
|
BR7 |
+ |
++ |
+ |
DEAD BOX RNA helicase RH15 |
|
BR15 |
− |
+ |
+ |
UDP-glucose 4-epimerase |
|
BR37 |
+ |
++ |
+ |
CBL-interacting protein kinase 1 |
|
BR39 |
− |
++ |
− |
Carbon-monoxide oxygenase (ACYB-1) |
|
BR40 |
+ |
+ |
− |
Phosphatase 2C, putative/PP2C |
|
BR74 |
+ |
+ |
− |
Haloacid dehydrogenase-like hydrolase |
|
BR91 |
+ |
+ |
− |
Unknown protein (AT3G12210) |
|
BR115 |
+ |
++ |
+ |
At5g25810 mRNA |
|
BR119 |
+ |
++ |
+ |
Unknown function |
|
BR503 |
− |
++ |
− |
Unknown function |
|
BR522 |
− |
++ |
− |
Hypothetical protein |
|
BR541 |
− |
++ |
− |
Light harvesting complex PSII |
|
BR547 |
− |
++ |
− |
UDP galactose transporter 3 |
Table 2Composition of media used for tissue culture and transformation of rice.
Table 2
|
Medium |
Composition |
|
2N6 (Embryogenic callus induction) |
4 g/L Chu (N6) medium (Micro and macro elements including vitamins) (Cat. No. C0204, Duchefa Biochemie B.V.), 0.3 g/L Casein hydrolysate, 0.5 g/L Glutamine, 0.5 g/L Proline, 30 g/L Sucrose, 2 mg/L 2.4-D, 2.5 g/L Gelrite (pH 5.8) |
|
AB (Agrobacterium culture) |
5 g/L Glucose, 15 g/L Bacto agar, 3 g/L K2HPO4, 1 g/L NaH2PO4, 1 g/L NH4Cl, 0.3 g/L MgSO4 · 7H2O, 0.15 g/L KCl, 13.25 mg/L CaCl2 · 2H2O, 2.5 mg/L FeSO4 · 7H2O (pH 7.2) |
|
AAM-AS (Agrobacterium suspension and infection) |
0.12 g/L MgSO4, 0.15 g/L CaCl2 · 2H2O, 0.15 g/L NaH2PO4 · 2H2O, 3 g/L KCl, 0.5 g/L Casamino acid, 0.177 g/L L-Arginine, 0.04 g/L EDTA Ferric sodium, 7.6 mg/L MnSO4 · H2O, 2 mg/L ZnSO4· 7H2O, 0.025 mg/L CuSO4 · 5H2O, 0.025 mg/L CoCl2 · 6H2O, 0.75 mg/L KI, 3 mg/L H3BO3, 0.25 mg/L Na2MoO4 · 2H2O, 0.75 mg/L Glycine, 0.09 g/L L-Glutamine, 0.03 g/L L-Aspartic acid, 0.01 g/L Myo-Inositol, 0.1 mg/L Nicotinic acid, 0.1 mg/L Pyridoxine HCl, 1 mg/L Thiamine HCl, 68.5 g/L Sucrose, 36 g/L D-Glucose, 20 mg/L Acetosyringone (pH 5.2) |
|
2N6-AS (Co-culture) |
4 g/L Chu (N6) medium (Micro and macro elements including vitamins) (Cat. No. C0204, Duchefa Biochemie B.V.), 0.3 g/L Casein hydrolysate, 0.5 g/L Glutamine, 0.5 g/L Proline, 30 g/L Sucrose, 10 g/L Glucose, 2 mg/L 2.4-D, 20 mg/L Acetosyringone, 0.15 g/L Dithiothreitol, 0.25 g/L Sodium thiosulfate, 5 mg/L Silver nitrate (pH 5.2) |
|
N6D (Transgenic callus selection) |
4 g/L Chu (N6) medium (Micro and macro elements including vitamins) (Cat. No. C0204, Duchefa Biochemie B.V.), 0.3 g/L Casein hydrolysate, 0.5 g/L Glutamine, 0.5 g/L Proline, 30 g/L Sucrose, 2 mg/L 2.4-D, 2.5 g/L Gelrite, 250 mg/L Cefotaxime, 50 mg/L Hygromycin (pH 5.8) |
|
REIII (Shoot formation) |
4.4 g/L MS medium including vitamins (Cat. No. M0222, Duchefa Biochemie B.V.), 0.5 mg/L NAA, 2 mg/L 6-BA, 20 g/L Sucrose, 30 g/L Sorbitol, 2 g/L Casamino acid, 250 mg/L Cefotaxime, 80 mg/L Hygromycin, 4 g/L Gelrite (pH 5.8) |
|
HF (Root formation) |
4.4 g/L MS medium including vitamins (Cat. No. M0222, Duchefa Biochemie B.V.), 30 g/L Sucrose, 7.5 g/L Plant agar, 250 mg/L Cefotaxime, 80 mg/L Hygromycin (pH 5.8) |
Table 3Primers sequences used in the RT-PCR analysis and annotation of gene function by the NCBI database.
Table 3
|
Gene |
Forward Primer |
Reverse Primer |
Gene Function |
|
BR 2 |
5′-GTGTTGACGGACGTTGTGAC-3′ |
5′-CCGGGTCTAGATCTGTTCCA-3′ |
Cysteine protease 1 |
|
BR 4 |
5′-TGCCGTTATGCCATTTATGA-3′ |
5′-ATCAAGACCCATCTCGGTTG-3′ |
Actin binding/depolymerizing factor 2 |
|
BR 7 |
5′-GCATTTCCTAAAGCCACCAA-3′ |
5′-CCAGGAGTAGGCTCGATCTG-3′ |
DEAD BOX RNA helicase RH15 - like protein |
|
BR 11 |
5′-AACGAGAGAGGAGGGATGAT-3′ |
5′-CCAGAGGAGCTTCAGGGTTG-3′ |
Alcohol dehydrogenase 1 |
|
BR 15 |
5′-GCTTTGCGTGCCTTCTTATC-3′ |
5′-GAAGAACCCACTTGGCAAAA-3′ |
UDP-glucose 4-epimerase 1 |
|
BR 22 |
5′-ACTTGGCGAGCTTGACTCAT-3′ |
5′-GAGCATTTCTCCTGCGTTTC-3′ |
Hydrogen transporting ATPase |
|
BR 23 |
5′-AATGCACGGAATAGAGG-3′ |
5′-CAGGTCCTCATTACCGGTGT-3′ |
Endoplasmic reticulum auxin-binding protein 2 mRNA |
|
BR 37 |
5′-ATCGGAGGCAGCAAGTCAAGCG-3′ |
5′-CTCCATCGTAGCCTCCGTCAT-3′ |
CBL-interacting protein kinase 1 |
|
BR 39 |
5′-CAGGAAAAGAAGCGGATGAG-3′ |
5′-AGTGTGCCTGTCATCGTGAG-3′ |
Carbon-monoxide oxygenase (ACYB-1) |
|
BR 43 |
5′-AACTCCAAGCGTTGGATATG-3′ |
5′-TCTCCAGGTTCGTTTGCTCT-3′ |
Hypothetical protein |
|
BR 47 |
5′-CTAATGCTGACGGTGGGTTT-3′ |
5′-ACGAAAAGGTCGAAAGCTGA-3′ |
Unknown gene function |
|
BR 61 |
5′-CCTCTGATCCCACCACACTT-3′ |
5′-TTAGGAGCAGCACACACTGG-3′ |
Rubredoxin family protein |
|
BR 64 |
5′-AGAAGCTGATGGCTCGGTTA-3′ |
5′-CCTCTTCTCGGCTGAACAAC-3′ |
Beta-lg-H3 domain-containing protein (FLA18) |
|
BR 68 |
5′-CCTTACCGCCGCCACCATGTTT-3′ |
5′-AGGTGTCAAAAGTCGGGTTG-3′ |
Unknown gene function |
|
BR 72 |
5′-ACCAAACACTCACCGGAGTC-3′ |
5′-ATCACCACCCGCAAGTAAAG-3′ |
Unknown protein function |
|
BR 74 |
5′-CGATGTTCGATGTCCTCCTT-3′ |
5′-GGCTCGATCATCGTCATTCT-3′ |
Haloacid dehalogenase-like hydrolase family protein |
|
BR 77 |
5′-CATCTACCGAGCAACAGCAA-3′ |
5′-AATGCCCAATCCATACCAAA-3′ |
Unknown (Zeamays clone 92871) |
|
BR 91 |
5′-TGGAAGCTCTCTCGTGGAA-3′ |
5′-CTCGTCGGACATAAATGGT-3′ |
Unknown protein |
|
BR 119 |
5′-GTATCACTCTCGGGGTTCCA-3′ |
5′-CCAAACAAATGGGACAATCC-3′ |
Unknown gene function |
|
BR 129 |
5′-GTGGAGGGAGTCAACGATGT-3′ |
5′-CCGAGTCCCTGCGATATTTA-3′ |
Dimethylmenaquinone methyltransferase family protein |
|
BR 192 |
5′-CATAAGGTACGCGACGGTTT-3′ |
5′-ACTAAAGCAGCTTCGGTCCA-3′ |
Glycosyltransferase family protein |
|
BR 195 |
5′-CACTGGAGTTGACCCAAC-3′ |
5′-AATGCTAAGGGCACCAACAC-3′ |
Aspartate aminotransferase 2 |
|
BR 289 |
5′-TGATCAGATGGAGTGGTGGA-3′ |
5′-CTCCTCGCTTGTTGGAAGAC-3′ |
Plastid-lipid associated protein PAP/fibrillin family protein |
Table 4Number of transgenic rice lines produced, confirmed gene inserts and its respective generation.
Table 4
|
Generation*
|
Confirmed no. of gene inserts |
No. of lines |
|
T1
|
137 |
470 |
|
T2
|
88 |
408 |
|
T3
|
14 |
46 |
|
T4
|
11 |
226 |
|
|
Total |
250 |
1,150 |
Appendix Table 1List of transgenic rice with full-length cDNA from B. rapa, its annotation function and their morpho-agronomic traits.
Appendix Table 1
|
Code |
Gene Function |
Cold stress scorea
|
Salinity stress scoreb
|
Observed phenotypec
|
|
BR 2 |
cysteine protease 1 |
5.7 |
9.0 |
more green leaves |
|
BR 4 |
ADF2 (Actin depolymerizing factor 2) |
5.0 |
9.0 |
more number of tiller |
|
BR 5 |
PsbQ2 mRNA for oxygen-evolving enhancer protein 3-2 |
3.0 |
9.0 |
more green leaves |
|
BR 6 |
Zea mays clone 93807 mRNA sequence |
5.0 |
9.0 |
more green leaves |
|
BR 7 |
Dead box RNA helicase RH15 - like protein |
3.0 |
3.0 |
same phenotype |
|
BR 9 |
PEC-1 mRNA for Lipid transfer protein |
5.0 |
9.0 |
more green leaves |
|
BR 12 |
Rac GTPase activator |
5.0 |
9.0 |
same phenotype |
|
BR 14 |
CBL-interacting protein kinase 3 |
5.2 |
5.5 |
less green leaves |
|
BR 17 |
mRNA for aluminum-induced |
5.0 |
9.0 |
less green leaves |
|
BR 23 |
endoplasmic reticulum auxin-binding protein 2 |
9.0 |
7.0 |
less green leaves |
|
BR 26 |
Brassica rapa subsp. pekinensis clone KBrH004A06, complete sequence |
5.3 |
9.0 |
same phenotype |
|
BR 29 |
Phospholipase C |
5.2 |
7.0 |
less green leaves |
|
BR 33 |
ABC transporter-like protein |
9.0 |
9.0 |
less green leaves |
|
BR 37 |
CBL-interacting protein kinase 1 |
5.4 |
3.0 |
less green leaves, less plant height |
|
BR 39 |
carbohydrate binding |
4.3 |
9.0 |
less green leaves |
|
BR 40 |
protein phosphatase 2C, putative/PP2C, |
4.7 |
3.0 |
less green leaves, shorter culm length |
|
BR 46 |
plasma membrane intrinsic protein 2 (PIP2) |
5.0 |
9.0 |
less green leaves, more taller |
|
BR 48 |
senescence-associated cysteine protease (CP1) |
4.8 |
5.0 |
more number of tiller |
|
BR 49 |
polypeptide of photosystem II reaction center |
9.0 |
9.0 |
less green leaves |
|
BR 50 |
chlorophyll a/b binding protein |
|
9.0 |
less green leaves |
|
BR 51 |
methyl-CpG-binding domain 4 (MBD4) |
9.0 |
9.0 |
less green leaves, longer panicle length |
|
BR 57 |
putative protein transport protein SEC61 alpha subunit |
5.0 |
7.0 |
less green leaves |
|
BR 60 |
Brassica rapa subsp. pekinensis clone KBrB042D24, complete sequence
|
5.0 |
9.0 |
less green leaves |
|
BR 61 |
rubredoxin family protein |
6.0 |
9.0 |
less green leaves |
|
BR 64 |
GDSL esterase/lipase APG |
5.0 |
9.0 |
less green leaves, longer panicle length, Shorter culm length |
|
BR 72 |
WSCP1 mRNA for water-soluble chlorophyll protein precursor |
5.0 |
9.0 |
same phenotype |
|
BR 73 |
unknown protein |
6.0 |
9.0 |
less green leaves |
|
BR 74 |
haloacid dehalogenase-like hydrolase family protein |
5.0 |
3.0 |
less green leaves, more number of tiller |
|
BR81 |
regulatory subunit of protein kinase CK2 |
5.0 |
7.0 |
less green leaves, shorter culm length |
|
BR 84 |
ATCTIMC (Cytosolic triose phosphate isomerase) |
5.0 |
9.0 |
shorter culm length |
|
BR 89 |
Brassica rapa subsp. pekinensis clone KBrH004P05, complete sequence |
5.7 |
9.0 |
same phenotype |
|
BR 91 |
unknown protein |
5.2 |
3.0 |
less green leaves, longer culm length |
|
BR 92 |
5-enolpyruvylshikimate-3-phosphate synthase (EPSP) |
5.0 |
9.0 |
same phenotype |
|
BR 97 |
G6PD1 (glucose-6-phosphate dehydrogenase 1) |
5.1 |
9.0 |
same phenotype |
|
BR 99 |
40S ribosomal protein S7 (RPS7C) |
5.0 |
9.0 |
same phenotype |
|
BR 104 |
protein binding/zinc ion binding |
5.0 |
9.0 |
same phenotype |
|
BR 105 |
Lhcb6 protein mRNA |
5.0 |
9.0 |
more number of tiller |
|
BR 111 |
protein kinase family protein |
5.0 |
9.0 |
same phenotype |
|
BR 112 |
Arabidopsis thaliana AT5g47540/MNJ7_13 mRNA, complete cds |
5.0 |
9.0 |
same phenotype |
|
BR 113 |
Brassica rapa subsp. pekinensis clone KBrB052L10, complete sequence |
5.8 |
9.0 |
same phenotype |
|
BR 114 |
Zea mays clone 13119 mRNA sequence |
5.0 |
9.0 |
less plant height |
|
BR 115 |
Arabidopsis thaliana At5g25810 mRNA, complete cds |
3.0 |
9.0 |
same phenotype |
|
BR 116 |
Unknown |
5.0 |
9.0 |
same phenotype |
|
BR 118 |
Vitis vinifera, whole genome shotgun sequence, contig VV78X164547.5, clone ENTAV 115 |
6.0 |
9.0 |
same phenotype |
|
BR 119 |
unknown protein |
3.0 |
3.0 |
shorter plant height |
|
BR 120 |
Zea mays clone 14950 mRNA sequence |
5.0 |
6.2 |
same phenotype |
|
BR 122 |
DNAJ heat shock N-terminal domain-containing protein |
5.0 |
9.0 |
same phenotype |
|
BR 124 |
putative thioredoxin reductase |
4.8 |
9.0 |
same phenotype |
|
BR 125 |
Brassica rapa subsp. pekinensis clone KBrS004A14, complete sequence |
5.0 |
9.0 |
less plant height |
|
BR 128 |
Zea mays clone 9139 mRNA sequence |
5.0 |
9.0 |
same phenotype |
|
BR 129 |
dimethylmenaquinone methyltransferase family protein |
5.0 |
9.0 |
same phenotype |
|
BR133 |
prephenate dehydratase family protein |
5.0 |
9.0 |
same phenotype |
|
BR139 |
231 CLA1-like protein mRNA |
5.1 |
9.0 |
same phenotype |
|
BR147 |
231 CLA1-like protein mRNA |
5.0 |
9.0 |
same phenotype |
|
BR148 |
protein kinase family protein/C-type lectin domain-containing protein |
5.0 |
5.2 |
same phenotype |
|
BR 149 |
XTR4 (xyloglucan endotransglycolase 4) |
5.0 |
9.0 |
same phenotype |
|
BR 151 |
clathrin adaptor complexes medium subunit-related |
5.0 |
9.0 |
same phenotype |
|
BR 152 |
unknown mRNA |
5.0 |
9.0 |
same phenotype |
|
BR 158 |
unknown protein (AT1G34010) mRNA |
5.0 |
9.0 |
same phenotype |
|
BR 160 |
SCL5; transcription factor (SCL5) |
5.0 |
5.8 |
same phenotype |
|
BR162 |
Brassica rapa subsp. pekinensis clone KBrB005N03, complete sequence |
5.0 |
9.0 |
same phenotype |
|
BR 165 |
Arabidopsis thaliana genomic DNA, chromosome 5, P1 clone:MZN1 |
5.0 |
9.0 |
more number of tiller |
|
BR 171 |
Brassica rapa subsp. pekinensis clone KBrH004A18, complete sequence |
5.5 |
9.0 |
more number of tiller |
|
BR 173 |
BSD domain-containing protein |
4.7 |
9.0 |
same phenotype |
|
BR 174 |
SEC14 cytosolic factor family protein/phosphoglyceride transfer family protein |
5.5 |
9.0 |
same phenotype |
|
BR 178 |
unknown protein |
4.0 |
9.0 |
same phenotype |
|
BR 179 |
Zea mays clone 8974 mRNA sequence |
5.5 |
5.6 |
same phenotype |
|
BR 192 |
glycosyltransferase family protein |
5.0 |
9.0 |
same phenotype |
|
BR 195 |
ASP2 (aspartate aminotransferase 2 (ASP2) |
5.7 |
9.0 |
same phenotype |
|
BR 225 |
unknown |
5.0 |
9.0 |
same phenotype |
|
BR 240 |
Brassica rapa subsp. pekinensis clone KBrH080C09, complete sequence |
5.0 |
9.0 |
same phenotype |
|
BR 246 |
CTR1 (constitutive triple response 1 |
5.0 |
9.0 |
same phenotype |
|
BR 250 |
unknown protein |
5.0 |
9.0 |
same phenotype |
|
BR 255 |
Indole-3-glycerol phosphate synthase |
5.5 |
9.0 |
same phenotype |
|
BR 256 |
leucine-rich repeat transmembrane protein kinase |
5.0 |
9.0 |
same phenotype |
|
BR 257 |
Unknown |
5.0 |
9.0 |
same phenotype |
|
BR 261 |
Arabidopsis thaliana Full-length cDNA Complete sequence from clone GSLTSIL12ZB10 of Silique of strain col-0 of Arabidopsis thaliana (thale cress) |
5.0 |
9.0 |
same phenotype |
|
BR 262 |
Glycosyl hydrolase family 17 protein |
4.8 |
9.0 |
same phenotype |
|
BR 263 |
CA2 (beta carbonic anhydrase 2); carbonate dehydratase/zinc ion binding (CA2) |
5.0 |
9.0 |
longer panicle length |
|
BR 264 |
Eukaryotic translation initiation factor 1A |
5.0 |
9.0 |
same phenotype |
|
BR 265 |
Zea mays clone 11869 mRNA sequence |
5.0 |
9.0 |
shorter plant height |
|
BR 271 |
rhodanese-like domain-containing protein |
5.0 |
9.0 |
same phenotype |
|
BR 273 |
WD-40 repeat family protein |
5.0 |
9.0 |
same phenotype |
|
BR 274 |
Brassica rapa subsp. pekinensis clone KBrH006E24, complete sequence |
5.0 |
9.0 |
same phenotype |
|
BR 275 |
pentatricopeptide (PPR) repeat-containing protein |
5.0 |
6.0 |
same phenotype |
|
BR 293 |
Zea mays clone 11032 mRNA sequence |
4.0 |
9.0 |
same phenotype |
|
BR 303 |
nodulin family protein |
5.0 |
9.0 |
same phenotype |
|
BR 305 |
Unknown protein |
5.0 |
9.0 |
same phenotype |
|
BR 308 |
Zea mays clone 15053 mRNA sequence |
4.9 |
9.0 |
same phenotype |
|
BR 311 |
Actin (Act1) mRNA |
5.0 |
9.0 |
same phenotype |
|
BR 318 |
unknown protein |
5.4 |
9.0 |
same phenotype |
|
BR 319 |
aldehyde dehydrogenase |
5.0 |
9.0 |
same phenotype |
|
BR 322 |
unknown protein |
5.0 |
9.0 |
same phenotype |
|
BR 336 |
unknown protein |
5.0 |
9.0 |
same phenotype |
|
BR 323 |
unknown protein |
5.0 |
9.0 |
same phenotype |
|
BR 330 |
unknown protein |
5.0 |
9.0 |
same phenotype |
|
BR 336 |
unknown protein |
5.0 |
9.0 |
shorter plant height |
|
BR 339 |
HDA6 (histone deacetylase 6) |
5.0 |
9.0 |
shorter plant height |
|
BR 343 |
ATNUDT2 (Arabidopsis thaliana Nudix hydrolase homolog 2); ADP-ribose diphosphatase/ NAD binding/hydrolase (ATNUDT2) mRNA, complete cds |
5.0 |
9.0 |
same phenotype |
|
BR 348 |
ELO1 (elongate 1) mRNA |
5.0 |
9.0 |
more green leaves, shorter plant height |
|
BR 349 |
unknown protein |
5.0 |
6.0 |
same phenotype |
|
BR 367 |
unknown gene function |
4.8 |
9.0 |
same phenotype |
|
BR 368 |
ribulose 1,5-bisphosphate carboxylase/oxygenase small subunit |
5.0 |
9.0 |
more green leaves |
|
BR 369 |
60S ribosomal protein L10A (RPL10aC) |
5.0 |
9.0 |
same phenotype |
|
BR 370 |
tubulin-specific chaperone C-related |
5.0 |
9.0 |
same phenotype |
|
BR 374 |
ABC1 family protein |
5.0 |
9.0 |
same phenotype |
|
BR 375 |
Zea mays clone 13431 mRNA sequence |
5.5 |
9.0 |
same phenotype |
|
BR 376 |
transcriptional repressor |
5.0 |
9.0 |
same phenotype |
|
BR 377 |
Arabidopsis thaliana clone 37944 mRNA, complete sequence |
5.0 |
9.0 |
same phenotype |
|
BR 383 |
CLA1-like protein mRNA |
5.0 |
9.0 |
same phenotype |
|
BR 384 |
unknown protein |
5.0 |
9.0 |
shorter plant height |
|
BR 386 |
60S ribosomal protein L28 (RPL28C) |
5.0 |
9.0 |
less green leaves, shorter plant height |
|
BR 393 |
unknown protein |
5.5 |
9.0 |
same phenotype |
|
BR 418 |
WNK1 (WITH NO LYSINE (K) 1); kinase (WNK1) mRNA |
5.2 |
9.0 |
same phenotype |
|
BR 425 |
Brassica rapa subsp. pekinensis clone KBrH080C09, complete sequence |
5.5 |
9.0 |
shorter plant height, longer panicle length |
|
BR 436 |
putative nuclear transport factor 2 mRNA |
5.0 |
9.0 |
same phenotype |
|
BR 438 |
60S acidic ribosomal protein P0 (RPP0B) |
5.0 |
9.0 |
same phenotype |
|
BR 445 |
transporter-related |
5.0 |
9.0 |
more green leaves |
|
BR 449 |
FTSH8 (FtsH protease 8); ATP-dependent peptidase/ATPase/ metallopeptidase/ zinc ion binding (FTSH8) mRNA |
5.0 |
6.9 |
same phenotype |
|
BR 450 |
Brassica rapa subsp. pekinensis clone KBrS015K01, complete sequence |
5.0 |
9.0 |
more green leaves |
|
BR 452 |
nuclear transport factor 2 mRNA |
5.2 |
9.0 |
more green leaves |
|
BR 499 |
Brassica rapa subsp. pekinensis clone KBrH003N18, complete sequence |
5.0 |
7.0 |
same phenotypE |
|
BR467 |
Arabidopsis thaliana PTAC16 (PLASTID TRANSCRIPTIONALLY ACTIVE 16); binding/catalytic (PTAC16) mRNA, complete cds |
5.0 |
9.0 |
same phenotype |
|
BR 468 |
Brassica rapa subsp. pekinensis clone KBrB034N10, complete sequence |
5.0 |
9.0 |
more green leaves |
|
BR 471 |
Arabidopsis lyrata subsp. lyrata hypothetical protein, mRNA |
5.0 |
9.0 |
same phenotype |
|
BR 470 |
B-box type zinc finger protein (Bbox1) mRNA |
9.0 |
9.0 |
more number of tiller |
|
BR 472 |
Brassica rapa subsp. pekinensis clone KBrB028I01, complete sequence |
5.0 |
6.4 |
same phenotype |
|
BR 473 |
Zea mays clone 17728 mRNA sequence |
5.0 |
9.0 |
shorter culm length |
|
BR 474 |
ACC oxidase (ACC0x2) |
5.0 |
9.0 |
same phenotype |
|
BR 475 |
Zea mays clone 17728 mRNA sequence |
5.0 |
9.0 |
more taller plant height |
|
BR 478 |
unknown protein |
5.0 |
9.0 |
same phenotype |
|
BR 479 |
Brassica rapa subsp. pekinensis clone KBrB021M22, complete sequence |
5.0 |
9.0 |
same phenotype |
|
BR 480 |
Arabidopsis thaliana unknown protein (AT3G18050) mRNA |
5.0 |
9.0 |
same phenotype |
|
BR 481 |
AL5 (ALFIN-LIKE 5); DNA binding/methylated histone residue binding (AL5) |
5.0 |
9.0 |
same phenotype |
|
BR 482 |
Arabidopsis thaliana LHCB3 (LIGHT-HARVESTING CHLOROPHYLL B-BINDING PROTEIN 3); structural molecule (LHCB3) mRNA, complete cds |
5.0 |
9.0 |
more green leaves, longer panicle length |
|
BR 482 |
unknown protein |
5.0 |
9.0 |
more green leaves |
|
BR 484 |
Zea mays clone 12745 mRNA sequence |
5.0 |
9.0 |
same phenotype |
|
BR 486 |
Arabidopsis thaliana LHCB3 (LIGHT-HARVESTING CHLOROPHYLL B-BINDING PROTEIN 3); structural molecule (LHCB3) mRNA, complete cds |
5.0 |
9.0 |
same phenotype |
|
BR 487 |
Brassica rapa subsp. pekinensis clone KBrB021M22, complete sequence |
5.0 |
9.0 |
same phenotype |
|
BR 488 |
unknown gene function |
5.7 |
9.0 |
more green leaves |
|
BR 489 |
Arabidopsis thaliana 60S ribosomal protein L13 (RPL13D) (AT5G23900) mRNA, complete cds |
5.0 |
9.0 |
more green leaves |
|
BR 490 |
short-chain dehydrogenase/reductase (SDR) family protein |
5.0 |
9.0 |
more green leaves |
|
BR 491 |
transducin family protein/WD-40 repeat family protein (AT2G16405) mRNA |
5.0 |
9.0 |
same phenotype |
|
BR 492 |
Phagocytosis and cell motility protein ELMO1-related (AT3G03610) mRNA |
5.0 |
9.0 |
same phenotype |
|
BR 494 |
HD2C (HISTONE DEACETYLASE 2C) |
5.0 |
9.0 |
same phenotype |
|
BR 495 |
unknown protein |
5.1 |
9.0 |
same phenotype |
|
BR 496 |
unknown gene function |
5.0 |
9.0 |
shorter culm length |
|
BR 497 |
unknown gene function |
5.0 |
9.0 |
same phenotype |
|
BR 498 |
unknown protein |
5.0 |
9.0 |
same phenotype |
|
BR 499 |
Brassica rapa subsp. pekinensis clone KBrH003N18, complete sequence |
5.0 |
9.0 |
same phenotype |
|
BR 485 |
Arabidopsis thaliana unknown protein (At1g15180) mRNA, complete cds/MATE efflux family protein [Arabidopsis thaliana] |
5.0 |
9.0 |
shorter culm length |
|
BR 500 |
Arabidopsis lyrata subsp. lyrata hypothetical protein, mRNA |
5.1 |
9.0 |
same phenotype |
|
BR 501 |
Brassica rapa subsp. pekinensis clone KBrB030F12, complete sequence |
5.0 |
9.0 |
same phenotype |
|
BR 502 |
Brassica oleracea var. botrytis tonoplast intrinsic protein bobTIP26-1 mRNA, complete cds |
5.0 |
9.0 |
same phenotype |
|
BR 503 |
metal tolerance protein 5 (MTP5) |
5.0 |
9.0 |
same phenotype |
|
BR 504 |
Brassica rapa subsp. pekinensis clone KBrH003N18, complete sequence |
5.0 |
9.0 |
same phenotype |
|
BR 505 |
Brassica napus 1433-3 mRNA, complete cds |
5.0 |
9.0 |
same phenotype |
|
BR 506 |
Brassica oleracea var. botrytis tonoplast intrinsic protein bobTIP26-1 mRNA, complete cds |
5.0 |
9.0 |
shorter culm length |
|
BR 507 |
Arabidopsis thaliana 60S ribosomal protein L21 (RPL21C) (AT1G09690) mRNA, complete cds |
5.0 |
9.0 |
shorter culm length |
|
BR 508 |
Arabidopsis thaliana BOU (A BOUT DE SOUFFLE); binding/transporter (BOU) mRNA, complete cds |
5.0 |
9.0 |
same phenotype |
|
BR 509 |
Arabidopsis thaliana ING1 (INHIBITOR OF GROWTH 1); DNA binding/methylated histone residue binding (ING1) mRNA, complete cds |
5.0 |
9.0 |
same phenotype |
|
BR 510 |
hypothetical protein |
5.0 |
9.0 |
shorter culm length |
|
BR 511 |
VHS domain-containing protein/GAT domain-containing protein |
5.3 |
9.0 |
same phenotype |
|
BR 512 |
RsGGT2 mRNA for gamma-glutamyl transferase |
5.0 |
9.0 |
same phenotype |
|
BR 513 |
hypothetical protein |
4.5 |
5.0 |
same phenotype |
|
BR 514 |
Arabidopsis thaliana clone 25991 mRNA, complete sequence |
5.0 |
9.0 |
same phenotype |
|
BR 515 |
Prephenate dehydrogenase family protein |
5.0 |
9.0 |
same phenotype |
|
BR 516 |
Brassica rapa subsp. pekinensis clone KBrB028I01, complete sequence |
5.0 |
9.0 |
same phenotype |
|
BR 517 |
ATP binding/kinase/ protein kinase/ protein serine/threonine kinase/ protein tyrosine kinase (NAK) |
5.0 |
9.0 |
same phenotype |
|
BR 518 |
Arabidopsis thaliana ATS1; glycerol-3-phosphate O-acyltransferase (ATS1) mRNA, complete cds |
3.0 |
9.0 |
same phenotype |
|
BR 519 |
Arabidopsis thaliana HVA22J (HVA22-LIKE PROTEIN J) |
5.0 |
9.0 |
same phenotype |
|
BR 520 |
Arabidopsis thaliana COX6B (CYTOCHROME C OXIDASE 6B); cytochrome-c oxidase (COX6B) mRNA, complete cds |
5.0 |
9.0 |
same phenotype |
|
BR 521 |
Zea mays clone 10882 mRNA sequence |
5.0 |
9.0 |
same phenotype |
|
BR 522 |
hypothetical protein, mRNA |
3.0 |
9.0 |
same phenotype |
|
BR 523 |
Arabidopsis thaliana AtMAPR2 (Arabidopsis thaliana membrane-associated progesterone binding protein 2); heme binding (AtMAPR2) mRNA, complete cds |
5.0 |
9.0 |
same phenotype |
|
BR 524 |
inorganic pyrophosphatase family protein, mRNA |
5.0 |
9.0 |
same phenotype |
|
BR 525 |
unknown gene function |
5.5 |
9.0 |
same phenotype |
|
BR 526 |
PsbQ2 mRNA for oxygen-evolving enhancer protein 3-2 |
5.0 |
9.0 |
same phenotype |
|
BR 527 |
Brassica rapa clone 231 CLA1-like protein mRNA, complete cd |
5.0 |
9.0 |
same phenotype |
|
BR 528 |
NAK; ATP binding/kinase/ protein kinase/ protein serine/threonine kinase/ protein tyrosine kinase (NAK) mRNA |
5.0 |
9.0 |
same phenotype |
|
BR 529 |
unknown gene function |
6.0 |
9.0 |
same phenotype |
|
BR 530 |
Myb family transcription factor |
5.2 |
9.0 |
same phenotype |
|
BR 531 |
Zea mays clone 14041 mRNA sequence |
5.0 |
9.0 |
same phenotype |
|
BR 532 |
Sterol 4-alpha-methyl-oxidase 2 (SMO2-2) |
5.0 |
9.0 |
same phenotype |
|
BR 533 |
hypothetical protein |
5.0 |
9.0 |
more taller plant height |
|
BR 534 |
hypothetical protein |
4.5 |
9.0 |
same phenotype |
|
BR 535 |
mRNA for ACC oxidase (ACC0x2) |
5.0 |
9.0 |
same phenotype |
|
BR 536 |
hypothetical protein |
5.0 |
9.0 |
same phenotype |
|
BR 537 |
Brassica rapa subsp. pekinensis clone KBrB021M22, complete sequence |
5.0 |
9.0 |
same phenotype |
|
BR 538 |
hypothetical protein |
5.0 |
9.0 |
same phenotype |
|
BR 539 |
Raphanus sativus SAUR gene |
5.0 |
9.0 |
same phenotype |
|
BR 541 |
LHCB4.2 (light harvesting complex PSII) |
3.0 |
9.0 |
more taller plant height, longer panicle length |
|
BR 542 |
Brassica rapa subsp. pekinensis clone KBrB048F07, complete sequence |
5.0 |
9.0 |
same phenotype |
|
BR 544 |
Brassica rapa subsp. pekinensis clone KBrH003N18, complete sequence |
5.0 |
9.0 |
more taller plant height |
|
BR 545 |
Arabidopsis thaliana mRNA for 50S ribosomal protein L3, complete cds, clone: RAFL04-10-N10 |
5.0 |
9.0 |
more taller plant height |
|
BR 546 |
Arabidopsis thaliana ATNUDX19 (ARABIDOPSIS THALIANA NUDIX HYDROLASE HOMOLOG 19); hydrolase/ metal ion binding (ATNUDX19) mRNA, complete cds |
4.0 |
9.0 |
more taller plant height |
|
BR 547 |
UDP-galactose transporter 3 (ATUTR3) |
3.0 |
5.0 |
same phenotype |
|
BR 548 |
A. lyrata subsp. lyrata inorganic pyrophosphatase family protein, mRNA |
5.0 |
9.0 |
more taller plant height |
|
BR 549 |
BcRUBP mRNA for ribulose 1,5-bisphosphate carboxylase/oxygenase |
5.0 |
9.0 |
more taller plant height |
|
BR 550 |
Arabidopsis thaliana ATRDH2 (ARABIDOPSIS THALIANA RHODANESE HOMOLOGUE 2); 3-mercaptopyruvate sulfurtransferase/ thiosulfate sulfurtransferase (ATRDH2) mRNA, complete cds |
5.0 |
9.0 |
shorter plant height |
|
BR 551 |
Arabidopsis thaliana RNA binding (AT4G39040) mRNA, complete cds |
5.0 |
5.0 |
same phenotype |
|
BR 552 |
ribosomal protein S6 family protein |
5.0 |
9.0 |
same phenotype |
|
BR 553 |
Brassica rapa PsbQ2 mRNA for oxygen-evolving enhancer protein 3-2, complete cds |
5.0 |
9.0 |
same phenotype |
|
BR 554 |
transducin family protein, mRNA |
5.0 |
9.0 |
same phenotype |
|
BR 555 |
Brassica rapa subsp. pekinensis clone KBrB034A02, complete sequence |
5.0 |
9.0 |
same phenotype |
|
BR 556 |
Hypothetical protein, mRNA |
5.0 |
9.0 |
more taller plant height |
|
BR 557 |
BnD21 mRNA sequence |
5.0 |
5.0 |
more taller plant height |
|
BR 558 |
unknown gene function |
5.0 |
9.0 |
more taller plant height |
|
BR 560 |
B.oleracea mRNA for ACC oxidase (ACC0x2) |
5.0 |
9.0 |
same phenotype |
|
BR 561 |
Arabidopsis thaliana COX6B (CYTOCHROME C OXIDASE 6B); cytochrome-c oxidase (COX6B) mRNA, complete cds |
5.0 |
5.8 |
longer culm length |
|
BR 562 |
Arabidopsis thaliana protein binding/zinc ion binding (AT5G45100) mRNA, complete cds |
5.0 |
9.0 |
same phenotype |
|
BR 563 |
Brassica rapa subsp. pekinensis clone KBrB036G18, complete sequence |
5.0 |
9.0 |
same phenotype |
|
BR 566 |
Arabidopsis thaliana ALB3 (ALBINO 3); P-P-bond-hydrolysis-driven protein transmembrane transporter (ALB3) mRNA, complete cds |
5.0 |
9.0 |
same phenotype |
|
BR 567 |
Zea mays clone 12993 mRNA sequence |
9.0 |
9.0 |
same phenotype |
|
BR 569 |
Zea mays clone 14041 mRNA sequence |
5.0 |
9.0 |
same phenotype |
|
BR 571 |
Arabidopsis thaliana mRNA for putative mrp protein, complete cds, clone: RAFL05-19-J04 |
5.0 |
9.0 |
same phenotype |
|
BR 572 |
Brassica rapa clone 231 CLA1-like protein mRNA, complete cds |
5.0 |
9.0 |
same phenotype |
|
BR 579 |
Zea mays clone 20053 mRNA sequence |
5.0 |
9.0 |
same phenotype |
|
BR 734 |
Arabidopsis thaliana PDCB1 (PLASMODESMATA CALLOSE-BINDING PROTEIN 1); callose binding/polysaccharide binding (PDCB1) mRNA, complete cds |
5.0 |
9.0 |
same phenotype |
|
BR 582 |
isoflavone reductase family protein |
5.0 |
9.0 |
same phenotype |
|
BR 583 |
hypothetical protein |
5.0 |
9.0 |
same phenotype |
|
BR 525 |
disease resistance response protein |
5.0 |
6.8 |
same phenotype |
|
BR 633 |
ribosomal protein L9 family protein |
5.0 |
9.0 |
same phenotype |
|
BR 640 |
TMS membrane family protein/tumour differentially expressed (TDE) family protein |
5.0 |
9.0 |
same phenotype |
|
BR 660 |
translational inhibitor protein like |
5.0 |
9.0 |
same phenotype |
|
BR 670 |
RPL9 (RIBOSOMAL PROTEIN L9); structural constituent of ribosome (RPL9) |
5.0 |
9.0 |
same phenotype |
|
BR 700 |
tonoplast intrinsic protein bobTIP26-1 |
5.0 |
9.0 |
same phenotype |
|
BR 708 |
ATPHB2 (PROHIBITIN 2) (ATPHB2) |
5.0 |
9.0 |
same phenotype |
|
BR 721 |
unknown protein |
5.0 |
5.3 |
same phenotype |
|
BR 725 |
40S ribosomal protein S4 (RPS4A) |
5.0 |
9.0 |
same phenotype |
|
BR 735 |
SPP2 (SUCROSE-6F-PHOSPHATE PHOSPHOHYDROLASE 2); catalytic/ magnesium ion binding/phosphatase/ sucrose-phosphatase (SPP2) |
5.0 |
9.0 |
same phenotype |
|
BR 743 |
mitochondrial pyruvate dehydrogenase kinase (PDK) |
5.0 |
7.0 |
same phenotype |
References
- Abe H, Narusaka Y, Sasaki I, Hatakeyama K, Shin IS, Narusaka M, Fukami-Kobayashi F, Matsumoto S, Kobayashi M. 2011. Development of full-length cDNAs from chinese cabbage (Brassica rapa Subsp. pekinensis) and identification of marker genes for defence. DNA Research. 18: 277-289.
- Alexandrov NN, Brover VV, Freidin S, Troukhan ME, Tatarinova TV, Zhang H, Swaller TJ, Lu YP, Bouck J, Flavell RB, Feldmann KA. 2009. Insights into corn genes derived from large-scale cDNA sequencing. Plant Mol Biol. 69: 179-194.
- Alexandrov NN, Troukhan ME, Brover VV, Tatarinova T, Flavell RB, Feldmann KA. 2006. Features of Arabidopsis genes and genome discovered using full-length cDNAs. Plant Mol Biol. 60: 69-85.
- An G, Jeong DH, Jung KH, Lee S. 2005. Reverse genetic approaches for functional genomics of rice. Plant Mol Biol. 59: 111-123.
- Cheng C, Daigen M, Hirochika H. 2006. Epigenetic regulation of the rice retrotransposon Tos17. Mol Genet Genomics. 276: 378-390.
- Cho YG, Kang HJ, Lee JS, Lee YT, Lim SJ, Gauch H, Eun MY, McCouch SR. 2007. Identification of quantitative trait loci in rice for yield, yield components, and agronomic traits across years and locations. Crop Sci. 47: 2403-2417.
- Choi IS, Kim YG, Cho YC, Hong HC, Baek MK. 2007. A medium-maturing, multi-disease resistant and good eating-quality rice variety “Gopum”. Kor J of Plant Breeding Sci. 39: 586-587.
- Fagard M, Vaucheret H. 2000. (Trans)gene silencing in plants: how many mechanisms? Plant Mol Biol. 51: 167-174.
- Gelvin SB. 2003. Agrobacterium-mediated plant transformation: The biology behind the “gene-jockeying” tool. Microbiol Mol Biol Rev. 67: 16-37.
- Hirochika H, Guiderdoni E, An G, Hsing YI, Eun MY, Han C, Upadhyaya N, Ramachandran S, Zhang Q, Pereira A, Sundaresan V, Leung H. 2004. Rice mutant resources for gene discovery. Plant Mol Biol. 54: 325-334.
- Ichikawa T, Nakazawa M, Kawashima M, Iizumi H, Kuroda H, Kondou Y, Tsuhara Y, Suzuki K, Ishikawa A, Seki M, Fujita M, Motohashi R, Nagata N, Takagi T, Shinozaki K, Matsui M. 2006. The FOX hunting system: an alternative gain-of-function gene hunting technique. Plant J. 48: 974-985.
- International Rice Genome Sequencing Project (IRGSP)2005. The map-based sequence of the rice genome. Nature. 436: 793-800.
- International Rice Research Institute (IRRI).2007. Standard evaluation system for rice. IRRI. Los Baños, Laguna Philippines: pp. 56.
- Jeong DH, An S, Kang HG, Moon S, Han JJ, Park S, Lee HS, An K, An G. 2002. . insertional mutagenesis for activation tagging in rice. Plant Physiol. 130: 1636-1644.
- Jia J, Fu J, Zheng J, Zhou X, Huai J, Wang J, Wang M, Zhang Y, Chen X, Zhang J, Zhao J, Su Z, Lv Y, Wang G. 2006. Annotation and expression profile analysis of 2073 full-length cDNAs from stress-induced maize. Zea mays L. seedlings. Plant J. 48: 710-727.
- Kaeppler SM, Kaeppler HF, Rhee Y. 2000. Epigenetic aspects of somaclonal variation in plants. Plant Mol Biol. 43: 179-188.
- Karlowski WM, Schoof H, Janakiraman V, Stuempflen V, Mayer KF. 2003. MOsDB: an integrated information resource for rice genomics. Nucleic Acids Res. 31: 190-192.
- Kurata N, Yamazaki Y. 2006. Oryzabase: an integrated biological and genome information database for rice. Plant Physiol. 140: 12-17.
- Lee HJ, Abdula SE, Jee MG, Jang DW, Cho YG. 2011. High-efficiency and Rapid Agrobacterium-mediated genetic transformation method using germinating rice seeds. J Plant Biotechnol. 38: 251-257.
- Manigbas NL, Park DS, Park SK, Han SI, Park HM, Hwang WH, Kim SM, Wang HJ, Kang HW, Kim DH, Yun DJ, Yi GW. 2011. Enhanced tolerance of transgenic rice overexpressing Arabidopsis thaliana nucleoside diphosphate kinase (AtNDPK2) against various environmental stresses. Philipp Agric Scientist. 94: 29-37.
- Marathe R, Anandalakshm R, Smith TH, Pruss GJ, Vance VB. 2000. RNA viruses as inducers, suppressors and targets of post-transcriptional gene silencing. Plant Mol Biol. 43: 295-306.
- Miyao A, Iwasaki Y, Kitano H, Itoh J, Maekawa M, Murata K, Yatou O, Nagato Y, Hirochika H. 2007. A large-scale collection of phenotypic data describing an insertional mutant population to facilitate functional analysis of rice genes. Plant Mol Biol. 63: 625-635.
- Nakamura H, Hakata M, Amano K, Miyao A, Toki N, Kajikawa M, Pang J, Higashi N, Ando S, Toki S, Fujita M, Enju A, Seki M, Nakazawa M, Ichikawa T, Shinozaki K, Matsui M, Nagamura Y, Hirochika H, Ichikawa H. 2007. A genome-wide gain-of-function analysis of rice genes using the FOX-hunting system. Plant Mol Biol. 65: 357-371.
- Noro Y, Takano-Shimizu T, Syono K, Kishima Y, Sano Y. 2007. Genetic variations in rice in vitro cultures at the EPSPs-RPS20 region. Theor Appl Genet. 114: 705-711.
- Ohyanagi H, Tanaka T, Sakai H, Shigemoto Y, Yamaguchi K, Habara T, Fujii Y, Antonio BA, Nagamura Y, Imanishi T, Ikeo K, Itoh T, Gojobori T, Sasaki T. 2006. The rice annotation project database (RAP-DB): hub for Oryza sativa ssp. japonica genome information. Nucleic Acids Res. 34: 741-744.
- Ouyang S, Zhu W, Hamilton J, Lin H, Campbell M, Childs K, Thibaud-Nissen F, Malek RL, Lee Y, Zheng L, Orvis J, Haas B, Wortman J, Buell CR. 2007. The TIGR rice genome annotation resource: improvements and new features. Nucleic Acids Res. 35: 883-887.
- Ralph SG, Chun HJE, Cooper D, Kirkpatrick R, Kolosova N, Gunter L, Tuskan GA, Douglas CJ, Holt RA, Jones SJM, Marra MA, Bohlmann J. 2008. Analysis of 4,664 high-quality sequence-finished poplar full length cDNA clones and their utility for the discovery of genes responding to insect feeding. BMC Genomics. 9: 57
- Rice Annotation Project2007. Curated genome annotation of Oryza sativa ssp. japonica and comparative genome analysis with Arabidopsis thaliana. Genome Res. 17: 175-183.
- Sasaki T, Matsumoto T, Antonio BA, Nagamura Y. 2005. From mapping to sequencing, post-sequencing and beyond. Plant Cell Physiol. 46: 3-13.
- Schrider DR, Hahn MW. 2010. Gene copy-number polymorphism in nature. Proc Biol Sci. 277: 3213-3221.
- Seki M, Narusaka M, Kamiya A, Ishida J, Satou M, Sakurai T, Nakajima M, Enju A, Akiyama K, Oono Y, Muramatsu M, Hayashizaki Y, Kawai J, Carninci P, Itoh M, Ishii Y, Arakawa T, Shibata K, Shinagawa A, Shinozaki K. 2002. Functional annotation of a full-length Arabidopsis cDNA collection. Science. 296: 141-145.
- Seki M, Carninci P, Nishiyama Y, Hayashizaki Y, Shinozaki K. 1998. High-efficiency cloning of Arabidopsis full-length cDNA by biotinylated CAP trapper. Plant J. 15: 707-720.
- Seki M, Shinozaki K. 2009. Functional genomics using RIKEN Arabidopsis thaliana full-length cDNAs. J Plant Res. 122: 355-366.
- Sun MM, Abdula SE, Lee HY, Cho YC, Han LZ, Koh HJ, Cho YG. 2011. Molecular aspect of good eating quality formation in japonica rice. PLoS ONE. 6(4): e18385
- Toki S, Hara N, Ono K, Onodera H, Tagiri A, Oka S, Tanaka H. 2006. Early infection of scutellum tissue with Agrobacterium allows high-speed transformation of rice. Plant J. 47: 969-976.
- Town CD, Cheung F, Maiti R, Crabtree J, Haas BJ, Wortman JR, Hine EE, Althoff R, Arbogast TS, Tallon LJ, Vigouroux M, Trick M, Bancroft I. 2006. Comparative genomics of Brassica oleracea and Arabidopsis thaliana reveal gene loss, fragmentation, and dispersal after polyploidy. Plant Cell. 18: 1348-1359.
- Umezawa T, Sakurai T, Totoki Y. 2008. Sequencing and analysis of approximately 40,000 soybean cDNA clones from a full-length-enriched cDNA library. DNA Res. 15: 333-346.
- Vaucheret H, Fagard M. 2001. Transcriptional gene silencing in plants: targets, inducers and regulators. TRENDS in Genetics. 17: 29-34.
- Wu JL, Wu C, Lei C, Baraoidan M, Bordeos A, Madamba MR, Ramos-Pamplona M, Mauleon R, Portugal A, Ulat VJ, Bruskiewich R, Wang G, Leach J, Khush G, Leung H. 2005. Chemical and irradiation-induced mutants of indica rice IR64 for forward and reverse genetics. Plant Mol Biol. 59: 85-97.
- Yang TJ, Kim JS, Kwon SJ, Lim KB, Choi BS, Kim JA, Jin M. 2006. Sequence-level analysis of the diploidization process in the triplicated FLOWERING LOCUS C region of Brassica rapa. Plant Cell. 18: 1339-1347.
- Zhang J, Li C, Wu C, Xiong L, Chen G, Zhang Q, Wang S. 2006. RMD: a rice mutant database for functional analysis of the rice genome. Nucleic Acids Res. 34: 745-748.
- Zhao W, Wang J, He X, Huang X, Jiao Y, Dai M, Wei S, Fu J, Chen Y, Ren X, Zhang Y, Ni P, Zhang J, Li S, Wang J, Wong GK, Zhao H, Yu J, Yang H, Wang J. 2004. BGI-RIS: an integrated information resource and comparative analysis workbench for rice genomics. Nucleic Acids Res. 32: 377-382.