Abstract
Resveratrol, a plant phenolic compound, has potential therapeutic benefits due to its antioxidant properties. This is substantiated by previous studies that show that resveratrol derived from rice grains is an effective treatment agent for metabolic syndrome. Here, we characterized the T-DNA sequence, inserted T-DNA structure, copy number, integrity of the transgene locus, resveratrol synthase gene expression and resveratrol contents in the grains of two resveratrol transgenic rice lines, Iksan515 and Iksan526. The T-DNA transformation vector contained two expression cassettes of the resveratrol synthase gene under the control of the ubiquitin promoter and the bar selection marker gene under the control of the CaMV35S promoter. Flanking sequence analysis indicated that the T-DNAs were inserted into intergenic regions of chromosome 4 for Iksan515 and chromosome 12 for Iksan526. Two T-DNAs connected in an inverted repeat structure at a single locus of the rice genome were identified by whole genome sequencing and Southern blot hybridization in both Iksan515 and Iksan526. No novel open reading frames (ORFs) around insertion sites, sequences encoding allergenic or toxic protein, or other unintended effects by T-DNA insertion were found in either case. In addition, resveratrol synthase gene expression in leaves and resveratrol detection in brown rice grains suggested the successful expression of the inserted foreign resveratrol synthase gene in two transgenic rice lines.
-
Key words: Molecular characterization, Resveratrol, T-DNA, Transgenic rice
INTRODUCTION
The plant phenolic compound resveratrol is a member of the stilbene family and is biosynthesized through the condensation of 1 molecule of
p-coumaroyl CoA and 3 molecules of malonyl CoA under the catalysis of
resveratrol synthase (
Rupprich et al. 1980). Resveratrol is found in over 70 plant species, including peanut (
Arachis hypogaea), grapevine (
Vitis vinifera), pine (
Pinus sylvestris), and Japanese knotweed (
Fallopia japonica); the latter has been used for traditional medicinal plant in China and Japan. Resveratrol acts as a defensive phytoalexin in plant cells as its levels increase in response to fungal infections (biotic stress) or other physical stresses such as UV radiation, ultrasound, and wounding, or in response to chemical exposure (
Langcake and Pryce 1976). The compound is also associated with the “French Paradox” as it is present in red wine, which at certain dosages is associated with cardiovascular benefits and protection (
Jeandet et al. 1995). Growing research suggests that resveratrol plays a role in the prevention of diabetes, cardiovascular and neurodegenerative diseases, and some types of cancers due to its antioxidant properties (
Baur et al. 2006;
Athar et al. 2007;
Saiko et al. 2008). To date, the main market for resveratrol is in nutraceuticals, which are based on raw materials from field cultures of
P. cuspidatum and microorganisms or plants cultured for large-scale production (
Vercauteren et al. 2003).
In the past two decades, many plants have been genetically modified by transferring
stilbene synthase (STS) or
resveratrol synthase (RS) gene to produce resveratrol for its potential roles in health promotion and in the control of disease in plants such as tobacco and tomato (
Fischer et al. 1997;
Thomzik et al. 1997). Resveratrol engineering was used to confer resistance of tobacco to
Botrytis cinerea (
Hain et al. 1993), resistance of rice to
Pyricularia oryzae (
Stark-Lorenzen et al. 1997), resistance of barley and wheat to
Botrytis cinerea (
Leckband et al. 1998), resistance of white poplar to
Melampsora pulcherrima (
Giorcelli et al. 2004), and resistance of papaya to
Phytophthora palmivora (
Zhu et al. 2004). To date, however, few studies on the benefits to human health of resveratrol derived from cereal grains have been performed.
Baek et al. (2013) developed resveratrol transgenic rice lines by introducing the
AhSTS1 (
RS3, GenBank accession no. DQ124938) gene into the commercial rice variety Dongjin. Of the lines tested in that study, RS18 (named Iksan526 in this paper), which contained a single T-DNA insertion resulted in relatively high transgene expression and had good performance of agronomic traits. Thus, RS18 was selected for inclusion in diets of experimental mice, and was found to significantly reduce body weight, blood glucose, triglycerides, total cholesterol, and low-density lipoprotein (LDL)-cholesterol. However, detailed molecular characterization of the resveratrol transgenic rice lines was not performed. Such characterization is required for performing elite event selection, and is necessary for biosafety assessment of genetically modified (GM) crops. In the present study, therefore, we performed comprehensive molecular characterization in two elite transgenic lines (Iksan515 and -526,
Baek et al. 2013), including transformation vector sequencing and the genomic features related to its integration site.
MATERIALS AND METHODS
Plant materials
T6 generation of the two transgenic Dongjin rice lines Iksan515 and Iksan526, expressing resveratrol synthase (RS3) and the selectable marker bar gene were used in this study. Transgenic lines were planted in the genetically modified organism (GMO) experimental fields of RDA, Suwon in 2011. Two lines of isogenic variety Dongjin were also planted per ten transgenic lines with a spacing pattern of 15 cm between plants and 30 cm between rows.
DNA and RNA isolation
Genomic DNAs were isolated from young leaves of rice plants according to the modified cetyltrimethylammonium bromide (CTAB) method (
Chen and Ronald 1999) following RNase treatment. Total RNAs were extracted from rice leaves by using TRIzol reagent (Invitrogen, UK) according to the manufacturer’s protocol. DNA was removed from RNA samples by using TURBO DNA-Free DNase (AMBION INC). Total DNAs and RNAs were quantified by measuring absorbance at 260 nm and 280 nm using the Nanodrop ND 1000-spectrophotometer (Thermo Fisher Scientific Inc., Wilmington, NC, USA) respectively.
Adaptor ligation PCR with a T-DNA-specific primer pair (LB1: 5′-CGGCTATTGGTAATAGGACACTGG-3′; LB2: 5′-CAACCCTCAACTGAAACGGGCCGGA-3′) and an adaptor primer pair (Ada1: 5′-GGATCCTAATACGA CTCACTATAGGGC-3′; Ada2: 5′-CTATAGGGCTCGA GCGGC-3′) were applied for flanking sequence analysis, as described in
Balzergue et al. (2001). For confirmation of insertion site of T-DNA, event-specific primers were designed from the rice genome at which the T-DNA insertion site was determined according to the sequence data from adaptor ligation PCR (see
supplementary table 1). Ex-Taq PCR was performed as the manufacturer’s instructions (
TaKaRa Ex TagTM) and the PCR condition was as follows: 94°C for 5 min, 30 cycles of 94°C for 30 s, 55°C for 30 s and 72°C for 45 s, and final extension at 72°C for 5 min.
Approximately 20 μg of genomic DNA, digested with EcoRI, EcoRV, or SacI was separated on an agarose gel and then blotted onto a Hybond-N+ membrane (Amersham), according to the manufacturer’s instructions. A 500 bp long sequence of the bar or resveratrol synthase gene was DIG-labeled using the PCR DIG probe synthesis kit (Roche). The primers for bar and RS are as follows: bar-forward: GAAGTCCAGCTGCCAGAAAC, bar-reverse: AAGCA CGGTCAACTTCCGTA; RS-forward: GCATTGTTTGC GGATGGAGCTG, RS-reverse: TTATATGGCCACACT GCG. The PCR conditions were 95°C for 2 min, 40 cycles of 95°C for 30 s, 60°C for 30 s, 72°C for 45 s, followed by 72°C for 7 min.
Blotted membranes were incubated with 20 ml of pre-hybridization solution supplemented with 125 ng/ml of denatured salmon sperm DNA at 42°C for 2 h. Afterwards, the membrane containing EcoRI/EcoRV-digested DNA was hybridized with a bar probe, while the membrane containing Sac1/EcoRV-digested DNA was hybridized with an RS probe. The denatured probes were diluted in 4 ml hybridization solution (5 × SSC, 0.1% (w/v) N-lauroylsarcosine, 0.02% (w/v) SDS, 1% (v/v) blocking reagent solution, and 50% (v/v) formamide); hybridization was performed overnight at 42°C. After hybridization, two low-stringency and high-stringency washes (5 min at 42°C and 15 min at 60°C, respectively) were performed. Signals were detected by chemiluminescence following incubation with anti-DIG antibodies and CSPD, according to the manufacturer’s procedure (Roche).
Inserted T-DNA sequence analysis
Event-specific primers were designed according to the flanking sequence information for LA-PCR (see
Supplementary table 2). A total of 50 μl reaction mixture with 0.5 μl LA Taq, 100 ng DNA, 10× buffer, 10× MgCl
2 (25 mM), and 10 pmol primers, as the manufacturer’s suggestions, (
TaKaRa Bio Inc.) was subjected to (95°C for 2 min, 35 cycles of 98°C for 20 s and 68°C for 15 min with 10 min extension time at 72°C). The expected bands of PCR amplification separated on a 1% agarose gel were eluted using a gel extraction kit (QIAGEN) and then cloned into pGEM-T Easy vector for sequencing. Sequence assembly and alignment against transformation vector were achieved using the SeqMan and MegAlign programs from the DNASTAR package, version 8.5 (DNASTAR, Madison, WI).
Genomic DNA samples of Iksan515, Iksan526, and Dongjin were used for whole genome sequencing on the Illumina Sequencing platform according to the user guide (HiSeq 2000 User Guide, Illumina, USA). In brief, genomic DNA samples were fragmented into ~800 bp lengths using a DNA shearing machine Covaris (Covaris, CA, USA) and blunt end-repaired with exonuclease. The repaired DNA fragments were ligated with adaptors (53 bp and 68 bp for both ends). Size selection was performed on a 2% agarose gel. The portion of the gel corresponding to 400- to 600-bp DNA was excised and purified, and then, DNA was amplified through PCR for binding and clustering on the flow cell of the HiSeq. Sequences generated were
de novo assembled to create a contig sequence by using CLC Genomics Workbench (CLC bio, Denmark) and contigs were analyzed and compared with the Rice Reference Genome V7 (
http://rice.plantbiology.msu.edu/index.shtml).
About 5ug RNA of Iksan515 and Iksan526 was used to synthesize each pool of cDNA using the SuperScript III First-Strand Synthesis System (Invitrogen, Carlsbad, CA, USA). Semi-quantitative RT-PCR was performed with 250 ng of cDNA for 28 cycles, with an annealing temperature of 57°C for RS and 60°C for actin. For RS, the primers used were as follows: forward primer, 5′-ATGGTGTCTGTGA GTGGAATTCGC-3′; and reverse primer, 5′-TTATATG GCCACACTGCG-3′. Rice actin1 (Osactin) was used as an internal control, with the forward primer 5′-CTGCTAT GTACGTCGCCATC-3′ and the reverse primer 5′-AGTC TCATGGATACCCGCAG-3.
Analysis for resveratrol contents of transgenic rice grains
300 mg sample from brown rice grains of two resveratrol rice was separately ground (Retsch MM301, Germany) and extracted by 900 μl of 80% (v/v) methanol for 30 min at ambient temperature. After centrifugation (10000 g, 5 min, 4°C), the supernatants were filtered by 0.2 μm nylon membrane filter and then stored in a refrigerator (−20°C) until the injection into a high performance liquid chromatography (HPLC) system for resveratrol analysis.
The 1.0 μl injection volume of extracts obtained were analyzed on an ACQUITY UPLC BEH-C18 1.7 μm column (2.1 mm × 100 mm, Waters) under gradient elution using 0.1% formic acid in acetonitrile (A) and 0.1% formic acid in water (B) as mobile phase (0 min, 10:90 (A:B); 1.54 min, 10:90; 10 min, 15:85; 22 min, 25:75; 22.4 min, 90:10; 25 min, 90:10) with 0.4 mL/min of flow rate. The wavelength used for analyses detection was 308 nm.
RESULTS
Full sequence confirmation for transformation vector
A 5573-bp T-DNA transformation vector containing two expression cassettes was identified by sequencing. The first comprised the resveratrol synthase (
AhSTS1, RS3) gene under the control of the maize
Ubi1 promoter and nopaline synthase (
Nos) terminator. The second contained the
bar gene encoding phosphinothricin ammonia transferase (PAT), which confers resistance to bialaphos. PAT was under the control of the CaMV35S promoter and contained a 35S polyA region (
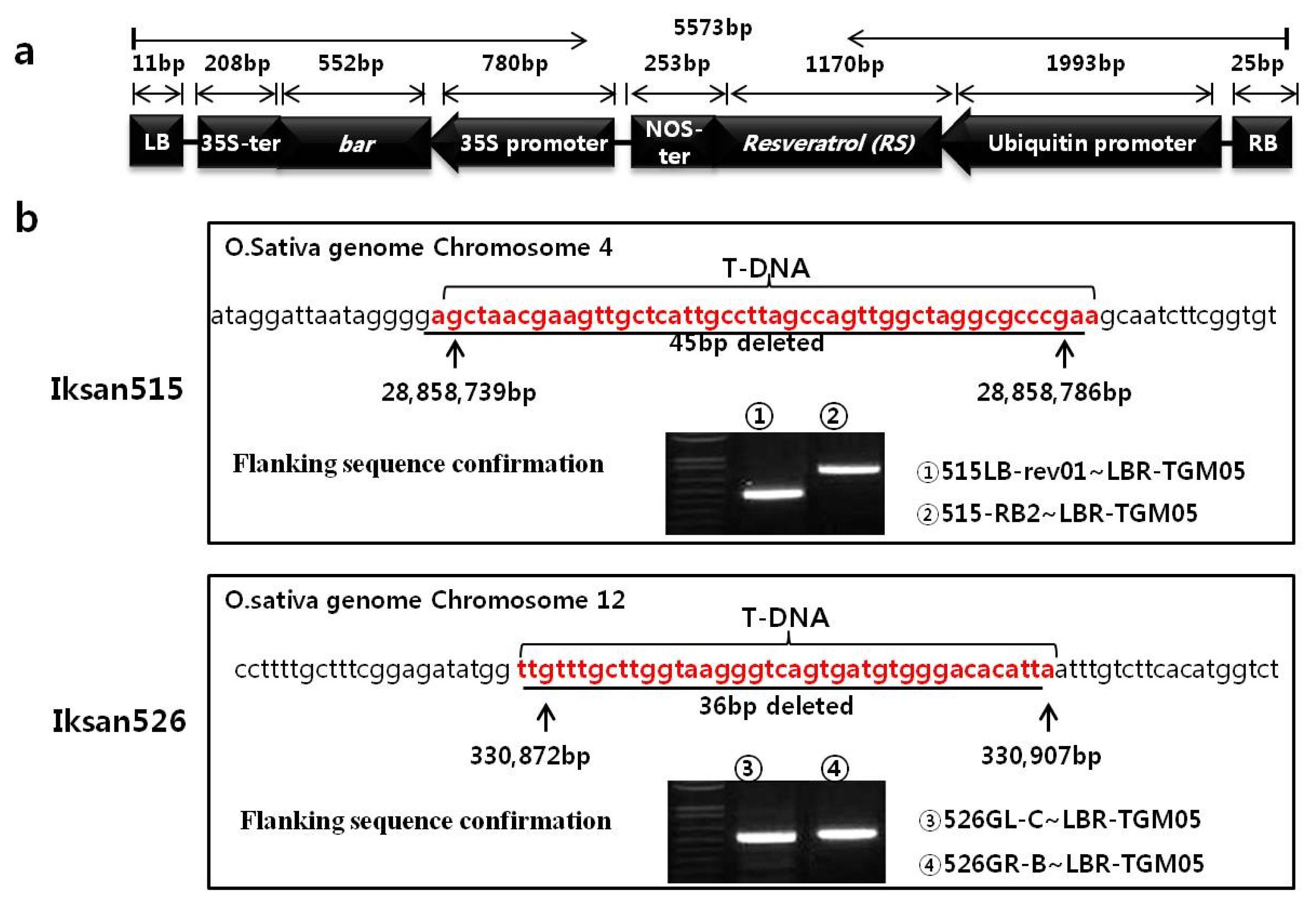
Fig. 1a). Based on the sequence information, structural units of regulatory elements and open reading frames for functional protein (ORFs) were confirmed by blastn, blastx, and ORF finder analysis from NCBI database (
http://www.ncbi.nlm.nih.gov). Moreover, blast analysis was performed to search for any similarity with known toxic proteins, using the nucleotide and/or deduced amino acid sequences of both PAT and RS; and no significant similarity was found. Potential allergenicity is a concern with proteins introduced into GM plants. Therefore, we searched an allergen database (
http://www.allergenonline.org/index.shtml, version 13) for proteins with similarity to PAT and RS. No matches of greater than 35% identity were found, suggesting that these two proteins carry a low risk of causing allergy.
Adaptor ligation PCR was carried out and sequence results revealed that the left border of T-DNA was inserted into an intergenic region from 28,858,739 bp to 28,858,786 bp (between Gene IDs Os04t0572800-00 and Os04t0573000~01) on chromosome 4 for Iksan515. For Iksan526, the insertion was from 330,872 bp to 330,907 bp (between Gene IDs Os12t0106200-01 and Os12t0106300-01) on chromosome 12 (
Fig. 1b). The sequences of expected sizes of 480 bp and 500 bp were amplified by PCR from the combinations of a common primer of LBR-TGM05 and event-specific primers (see
supplementary table 1) for flanking sequence confirmation (
Fig. 1b). Sequencing results confirmed the positive insertions of left border of T-DNAs for both transgenic lines (
Fig. 1b ① and ③). In addition, the insertions are associated with 45-bp and 36-bp deletions of rice genomic sequence.
The combination of the primer LBR-TGM05 and either 515-RB2 or 526GR-B (targeting the sequences downstream of T-DNA right border integration site on chromosome 4 or 12, respectively, see
supplementary table 1) for Ex-PCR amplification yielded a single band on each transgenic line (
Fig. 1b ② and ④). PCR bands sequencing results revealed that the left border of the transformation vector was inserted into the same locus to the first T-DNA, which were analyzed by adaptor ligation PCR for each transgenic line. We thus conclude that at least two inverted repeats of T-DNAs were present at a single genomic insertion site for both transgenic lines, and termed these “second T-DNA”.
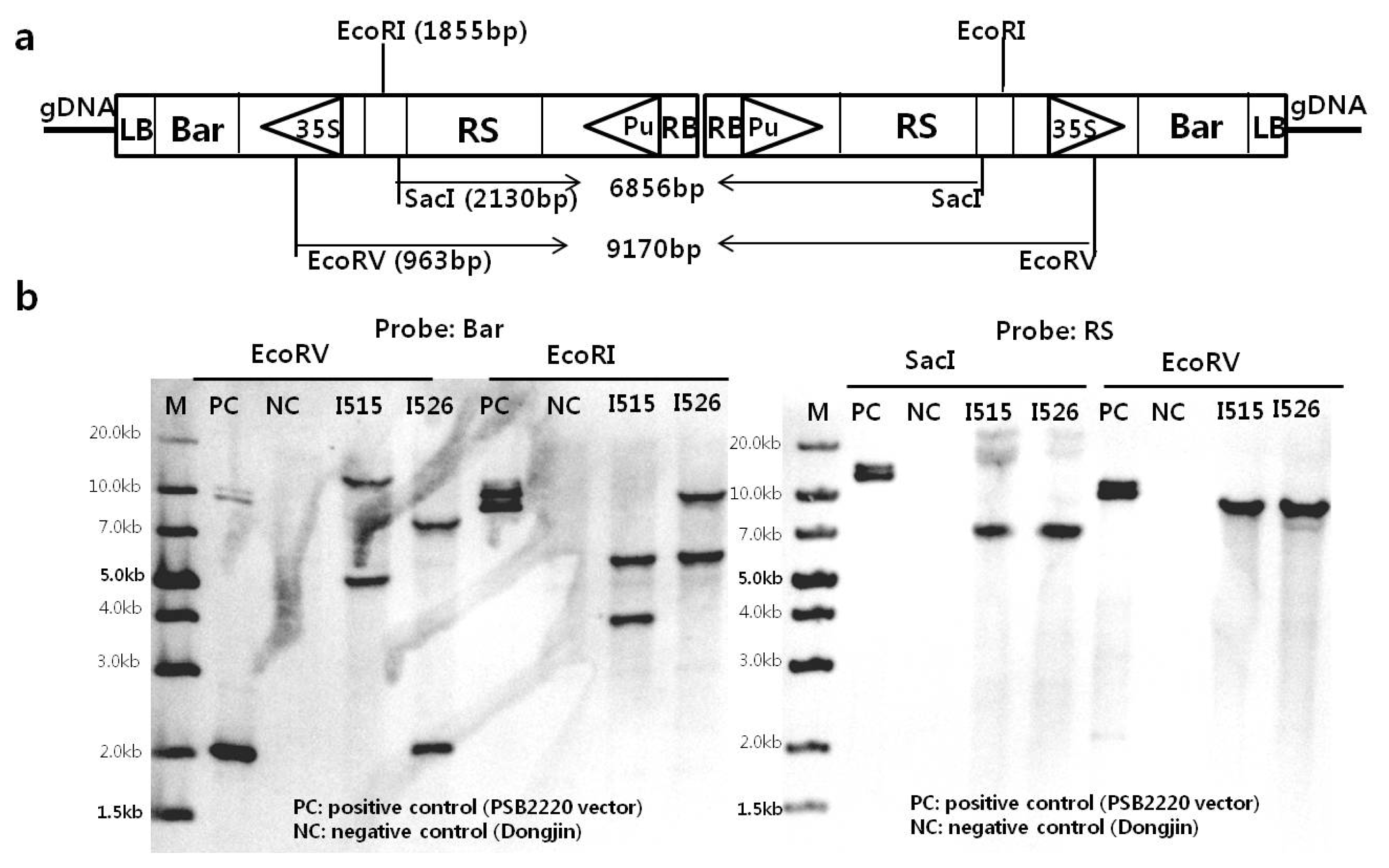
Southern blot hybridization was carried out to confirm the T-DNA copy number. The restriction sites in the construct vector were shown on
Fig. 2a. Based on the presumed inverse repeat structure of the T-DNAs, the size after enzyme digestion was calculated. If there are no additional T-DNAs inserted at the junction between the first and second T-DNAs, single bands should be observed on
EcoRV- or
SacI-digested DNA after RS probe hybridization, with expected band sizes of 6856 bp or 9170 bp for each transgenic strain, respectively.
Fig. 2b reveals this as the case, and that two T-DNAs were inserted into each transgenic line. Furthermore, similar band sizes were observed following
EcoRV or
EcoRI digestion and hybridization with the
bar probe. Likewise, only one band was evident after digestion with either
SacI or
EcoRV and hybridization with the RS probe. In addition, it appears that the structure of the regions at the right border between the two inverts is similar in both Iksan515 and Iksan526, although further evidence will be required to support this prediction.
High GC content of the
ubiquitin promoter region led to the failure of PCR amplification, thus the regions from flanking sequence of rice genome to part of
ubiquitin promoter of T-DNA were amplified by LA-PCR for two independent insertions of each transgenic line. Four primers consisting of 526GL-F02 and 526GL-C from the first T-DNA side and 526GR-B and 526GR-rv04 from the second T-DNA side of rice genomes were designed for Iksan526, respectively. Two primers of Pubi-rev30 and Pubi-rvs13 were designed from 4212 bp and 4302 bp of transformation vector sequence (these hybridize upstream of the 13 mer of G residues, and primer sequence in
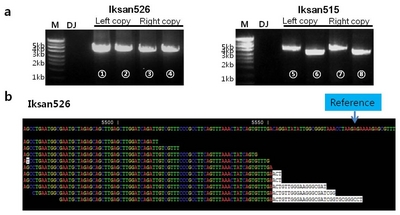
supplementary table 2). Bands of approximately 4~4.5 kb in length were amplified by primer combinations in Iksan526 (
Fig. 3a ①② and ③④), and Iksan515 (
Fig. 3a ⑤⑥ and ⑦⑧). Sequencing results with near-perfect correlation with genomic DNA and vector sequences were achieved, which is consistent with the results of our flanking sequence analysis.
To confirm sequences between ubiquitin promoters (containing right borders) of two T-DNAs, whole genome sequencing of two transgenic rice lines and donor variety Dongjin were performed. After paired-end sequencing, read lengths of 9.6 Gigabase (Gb), 10.9 Gb, and 13.7 Gb sequences corresponding to a coverage depth of 26x, 29x, and 37x were generated from Dongjin, Iksan515, and Iksan 526, respectively (
supplementary table 3).
De novo assembly generated 77,934, 66,428, and 59,860 contigs for Dongjin, Iksan515, and Iksan526, respectively (data not shown). All sequences were blast-analyzed against T-DNA; this resulted in selection of 2,492 matches from Iksan515 and 3,622 matches from Iksan526. Contigs were assembled and mapped to the vector sequence as shown in
Fig. 3b. For Iksan526, sequences of a contig ‘Iksan526_contig_2497’ were mapped to the reference vector. The final 30 bases inversely correlated with sequences in the
ubiquitin promoter region. This strongly suggests that in Ikan526, the two T-DNAs are directly connected in a tail-to-tail configuration and harbor a 58-bp deletion in the first T-DNA and a 146-bp deletion in the second T-DNA. This direct connection of two T-DNAs was confirmed by two expected restriction fragments of ~6.86 kb and ~9.2 kb based on the proposed structure in
Fig. 2a and Southern blot hybridization results in
Fig. 2b. We also observed similar molecular characteristics for the Iksan515 insertion.
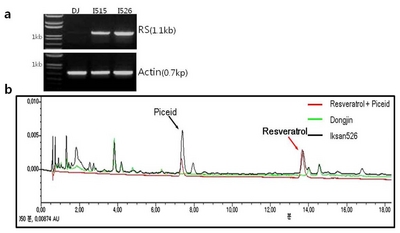
The
resveratrol synthase gene expressions in young leaves of T
6 generations of two transgenic rice lines Iksan515 and Iksan526 were measured by semi-quantitative RT-PCR as shown in
Fig. 4a. Resveratrol contents in brown rice grains of two transgenic rice lines were analyzed by HPLC. A peak at retention time of 14 min indicated the resveratrol (black line) detection in the brown rice grains of Iksan526, however, such was not detected in the donor variety Dongjin (green line) (
Fig. 4b and Iksan515 was shown on
supplementary Fig. 1). Comparing to the donor variety Dongjin, the resveratrol content in brown rice were 1.55 ± 0.03 ug/g for Iksan515 and 2.22 ± 0.08 ug/g for Iksan526, suggesting an effective expression of foreign resveratrol synthase gene.
DISCUSSION
Our detailed molecular characterization provided a new insight into the configuration of inserted T-DNA in two important transgenic rice lines. The tail-to-tail configurations of the T-DNA insertions in Iksan515 and Iksan526 were almost identical. However, the different chromosomal insertion sites and detailed sequence differences at the flanking regions indicate that they are independent transgenic lines.
Iksan515 and Iksan526 were created by
Agrobacterium-mediated transformation using LBA4404 of
A. tumefaciens strain as T-DNA binary system (
Baek et al. 2013). In contrast to other transformation methods,
Agrobacterium-mediated transformation generally results in lower transgene copy numbers (
Kohli et al. 2003;
Shou et al. 2004). However, multiple integrations and complex integration structures frequently occur during
Agrobacterium-mediated transformation (
Jorgensen et al. 1996;
Shou et al. 2004;
De Buck et al. 2009). After investigation of 177 rice T-DNA transformants,
Kim et al. (2003) estimated that approximately 33% of the transformants carried direct repeats, 21% had inverted repeats with the 5′-end junctions, and 7% carried inverted repeats with the 3′-end junctions. In addition,
Galbiati et al. (2000) reported that transformation using
A. tumefaciens LBA4404 resulted in an average transgene copy number of 1.0–1.7 in
Arabidopsis and 2.1–3.0 in maize, which are lower values than those obtained using other strains. Taken together, these results reveal that there is only a 30% probability of selecting single-copy T-DNA insert during LBA4404-mediated transformation, and that direct or invert repeat inserts occur in approximately 50% of all transformants. Despite this, our current results confirm that high level expression of
bar and
resveratrol synthase genes is still maintained in the presence of inverted repeat integrations.
The multiple or incomplete integration events pose technical challenges for characterization of transgenes using PCR-based chromosome-walking techniques (
Tonooka et al. 2009;
Zhang et al. 2012). In this study, we were confronted with a severe challenge for conventional PCR amplification due to the poly G region of the
ubiquitin promoter. Therefore, we performed whole genome sequencing for the two resveratrol transgenic lines and also for the parental isogenic Dongjin line as a reference. This high-throughput next-generation sequencing approach is able to determine the copy number of both the integrated transgene and the backbone of the vector (
Zhang et al. 2012), and is especially capable of detecting complex events, including those with multiple T-DNA insertions and sequence rearrangements (
Kovalic et al. 2012). In this study, the exact integration breakpoints spanning the transgene insertion junctions were finally identified after reference mapping and
de novo assembly, indicating an approximately 204-bp deletion at the two right borders of two T-DNA insertions in the junction parts of inverted repeats as shown in
Fig. 3b. Based on the PCR and whole genome re-sequencing data analysis, we propose an inverted repeat structure of the two T-DNA insertions, which contains some deletions at the right border region of the first T-DNA and the 5′-end
ubiquitin promoter region of the second T-DNA.
Southern blot hybridization is an effective method for identification of copy numbers of inserted T-DNA in transgenic plants. Previous Southern blot experiment studies by
Baek et al. (2013) identified one major band and some trace signal bands in Iksan526 (RS18). To further clarify these results, we combined Southern blot hybridization with whole genome sequencing analysis. This provided unambiguous sequence information of the unknown region from 4302 bp of the transformation vector to the right border of two inserted T-DNAs. Our study also showed that LA-PCR is an effective approach for providing long-fragment amplifications covering both genomic sequence and the entire T-DNA insertion.
Studies in dicot plants have identified vector backbone sequences that reside outside the left and right borders of the integration site at a frequency ranging from 15% to 75% (
Kononov et al. 1997;
De Buck et al. 2000). Furthermore,
Kim et al. (2003) observed that 45% of transformants carrying the left border of the vector backbone are able to integrate into the monocot rice chromosome. Notably, we did not find any novel ORFs within a 10-kb region in either direction of the insertion site in either Iksan515 or Iksan526 by PCR. At the same time, Southern blot hybridization was also performed using full backbone sequence as a probe and results revealed no backbone insertions into the whole rice genome of each transgenic rice line (
Supplementary Fig. 2).
In conclusion, our current results indicate that the insertional events in these transgenic lines pose little risk to human health and nutrition, and that these lines are a valuable resource for future studies of resveratrol-based nutraceuticals. Surely, the stability of the inserted DNA through generations is also a major concern, as this clearly will affect productivity and/or yield of the desired crop. In our case, insertion of two T-DNA copies with an inverted repeat structure may increase the likelihood of gene silencing, and reduce the stability of and expression from the T-DNA element through generations. Therefore, stability of the inserted T-DNA structure, T-DNA expression and phenotypic characteristics through multi-generations will be addressed in further investigation.
Supplementary materials
Supplementary Figure 1
HPLC analysis for resveratrol content of transgenic rice line Iksan515.
Supplementary Figure 2
Southern blot hybridization for two events by using full backbone sequence as a probe with ClaI and BamHI digested DNAs.
Supplementary Table 1Ex-PCR primers for flanking sequence confirmation.
Supplementary Table 1
|
Primers |
Primer sequence |
Primer location |
|
Primers of flanking regions from rice genomes |
|
|
526GL-C |
GTGCTTGTTGTCTTCTCTTACG |
200bp from 1st T-DNA on chr. 12 |
|
526GR-B |
GATATAGACCTAGCACAGAAAGG |
220bp from 2nd T-DNA on chr. 12 |
|
515LB-rev01 |
TTCCAGTTCCGCGCCGGCAAACCATGTCAG |
180bp from 1st T-DNA on chr. 4 |
|
515-RB2 |
GCAGAAGTGGACCAGCCGAAATTAGATC |
660bp from 2nd T-DNA on chr. 4 |
|
|
Common primer from plasmid vector (pSB2220) |
|
|
LBR-TGM05 |
TAGGGTTTCGCTCATGTGTTGAGCA |
300bp from left border |
Supplementary Table 2Primers designed for LA-PCR.
Supplementary Table 2
|
Primers |
Primer sequence |
Primer location |
|
Primers of flanking regions from rice genomes |
|
|
526GL-F02 |
CAATTCGGTCTCATTAGCCTACCTCA |
210bp from 1st T-DNA on chr. 12 |
|
526GL-C |
GTGCTTGTTGTCTTCTCTTACG |
200bp from 1st T-DNA on chr. 12 |
|
526GR-B |
GATATAGACCTAGCACAGAAAGG |
220bp from 2nd T-DNA on chr. 12 |
|
526GR-rv04 |
CAGAAAGGACACATGTCCCGTTGAGAGA |
210bp from 2nd T-DNA on chr. 12 |
|
515LB-rev01 |
TTCCAGTTCCGCGCCGGCAAACCATGTCAG |
180bp from 1st T-DNA on chr. 4 |
|
515-RB2 |
GCAGAAGTGGACCAGCCGAAATTAGATC |
660bp from 2nd T-DNA on chr. 4 |
|
|
Common primer from plasmid vector (pSB2220) |
|
|
Pubi-rvs13 |
CCCCCCTCTCTACCTTCTCTAGATC |
4302bp from left border |
|
Pubi-rev30 |
TTAGATCCGTGCTGCTAGCGTTC |
4212bp from left border |
|
Rs-OsFw01 |
ATGGTGTCTGTGAGTGGAATTCGC |
Start of resveratrol synthase |
|
LBR-TGM05 |
TAGGGTTTCGCTCATGTGTTGAGCA |
300bp from left border |
Supplementary Table 3Summary of whole genome sequencing results from Dongjin, Iksan515, and Iksan526.
Supplementary Table 3
|
Reads No |
Reads Length (bp) |
Coverage (X) |
|
Dongjin |
94,872,502 |
9,582,122,702 |
26 |
|
Iksan515 |
108,331,448 |
10,941,476,248 |
29 |
|
Iksan526 |
135,725,886 |
13,708,314,486 |
37 |
ACKNOWLEDGMENTS
This work was equally supported by grant PJ009528 from the Biogreen 21 R&D Program and grant PJ009652 from the National Academy of Agricultural Science, Rural Development Administration, Republic of Korea.
Fig. 1Transformation vector information and T-DNA flanking sequence confirmation. a. Transformation vector construction; b. T-DNA insertion site in rice genome and putative flanking sequence analysis for transgenic resveratrol rice lines Iksan515 and Iksan526 derived via Ex-PCR.
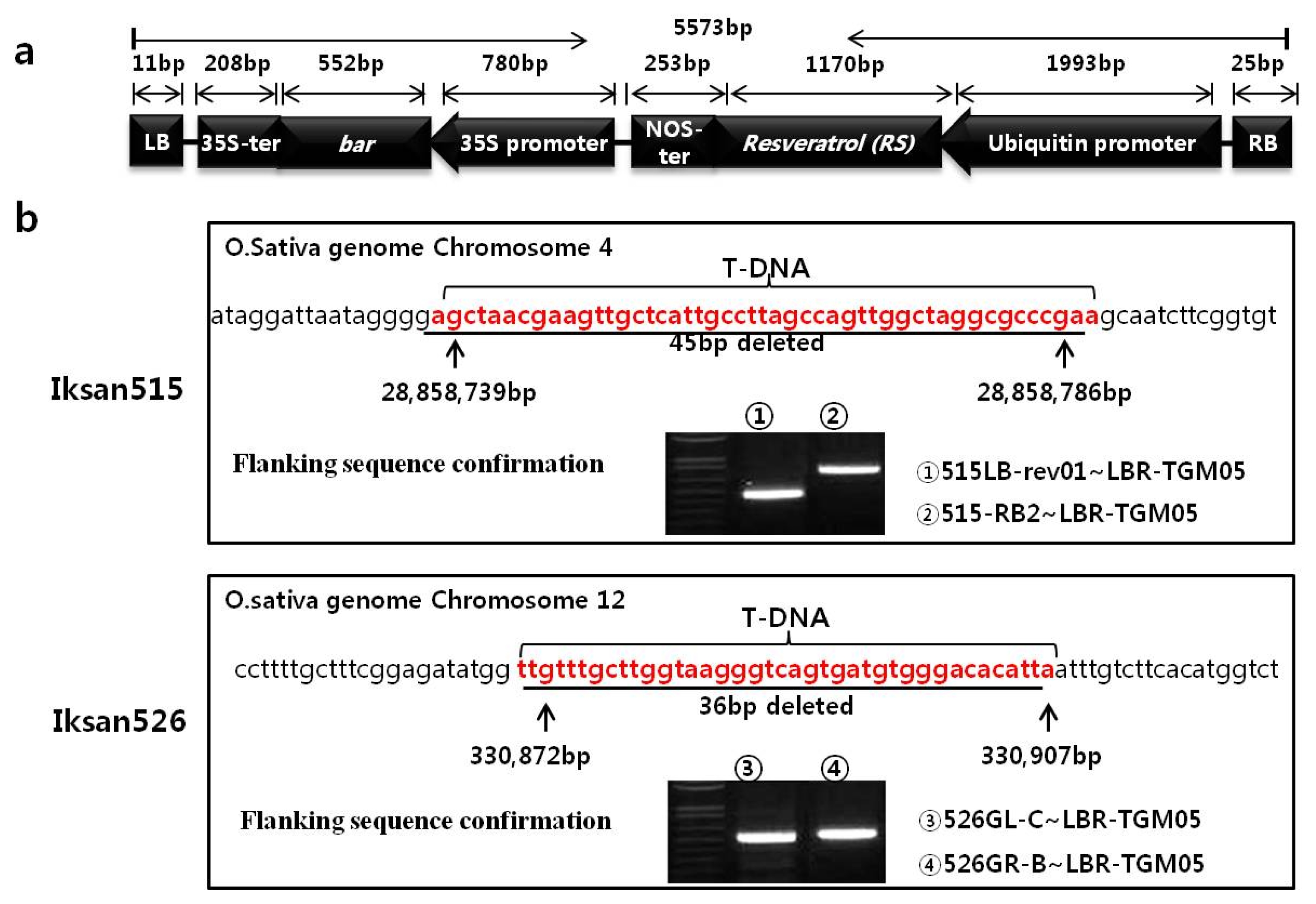
Fig. 2Copy number confirmation for two transgenic lines. a. The expected sizes of enzyme restriction sites between transformation vector and genomic DNAs. b. Confirmation of copy numbers of resveratrol GM rice by Southern blot hybridization using two probes from bar and RS. M: molecular weight size marker GeneRuler 1 kb-Plus DNA ladder; PC: positive control; NC: negative control: Dongjin.
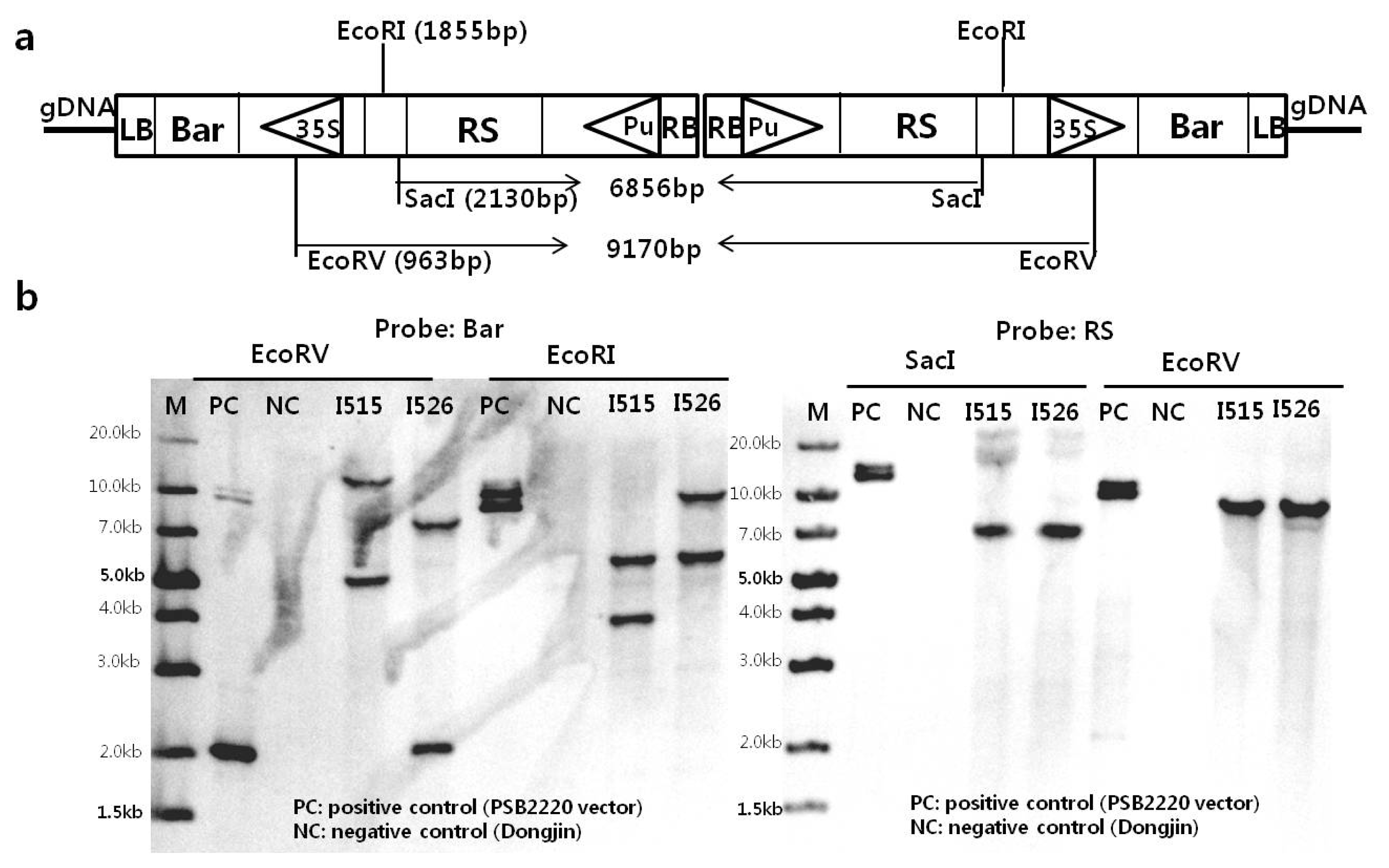
Fig. 3Inserted T-DNA structure confirmation. a. Long fragment amplifications from flanking regions to inserted T-DNA sequences of both events by using LA-PCR. Primer sets as follows: ①526GL-F02~Pubi-rev30; ②526GL-C ~Pubi-rvs13;③526GR-B~Pubi-rev30; ④526GR-rv04~Pubi-rev30; ⑤515LB-rev01~Pubi-rev30; ⑥515LB-rev01 ~Rs-OsFw01; ⑦515-RB2~Pubi-rvs13; ⑧515-RB2~Rs-OsFw01; b. Assembly of sequence reads generated from pair-end sequencing by a HiSeq2000 sequencer and reference mapping to transformation vector sequence. Reference: T-DNA parts of vector sequence; Colored letters (ATGC type, bases): sequences of individual reads from sequencer; black letters in white bracket (ATGC type, bases): mismatched sequences to reference vector.
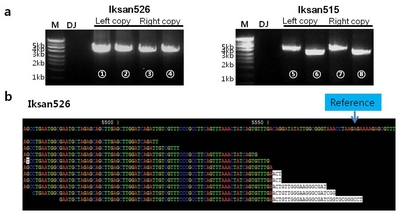
Fig. 4Resveratrol synthase gene expression in transgenic rice lines. a. Semi-quantitative RT-PCR for resveratrol synthase gene expression; b. HPLC analysis for resveratrol content in brown rice grains of the transgenic line Iksan526.
References
- Athar M, Back JH, Tang X, Kim KH, Kopelovich L, Bickers DR, Kim AL. 2007. Resveratrol: a review of preclinical studies for human cancer protection. Toxicol Appl Pharmacol. 224: 274-283.
- Baek SH, Shin WC, Ryu HS, Lee DW, Moon EJ, Seo CS, Hwang E, Lee HS, Ahn MH, Jeon YJ, Kang HJ, Lee SW, Kim SY, DSouza R, Kim HJ, Hong ST, Jeon JS. 2013. Creation of Resveratrol-Enriched Rice for the Treatment of Metabolic Syndrome and Related Diseases. PLoS One. 8: e57930
- Balzergue S, Dubreucq B, Chauvin S, Le-Clainche I, Le Boulaire F, de Rose R, Samson F, Biaudet V, Lecharny A, Cruaud C, Weissenbach J, Caboche M. 2001. Improved PCR-walking for large-scale isolation of plant T-DNA borders. Biotechniques. 30: 496-504.
- Baur JA, Sinclair DA. 2006. Therapeutic potential of resveratrol: the in vivo evidence. Nat Rev Drug Discov. 5: 493-506.
- Chen DH, Ronald PC. 1999. A rapid DNA minipreparation method suitable for AFLP and other PCR applications. Plant Mol Biol Rep. 17: 53-57.
- De Buck S, De Wilde C, Van Montagu M, Depicker A. 2000. T-DNA vector backbone sequences are frequently integrated into the genome of transgenic plants obtained by Agrobacterium-mediated transformation. Mol Breed. 6: 459-468.
- De Buck S, Podevin N, Nolf J, Jacobs A, Depicker A. 2009. The T-DNA integration pattern in Arabidopsis transformants is highly determined by the transformed target cell. Plant J. 60: 134-145.
- Fischer R, Budde I, Hain R. 1997. Stilbene synthase gene expression causes changes in flower color and male sterility in tobacco. Plant J. 11: 489-498.
- Galbiati M, Moreno MA, Nadzan G, Zourelidou M, Dellaporta SL. 2000. Large-scale T-DNA mutagenesis in Arabidoposis for functional genomic analysis. Funct Intergr Genomics. 1: 25-34.
- Giorcelli A, Sparvoli F, Mattivi F, Tava A, Balestrazzi A, Vrhovsek U, Calligari P, Bollini R, Confalonieri M. 2004. Expression of the stilbene synthase (StSy) gene from grapevine in transgenic white poplar results in high accumulation of the antioxidant resveratrol glucosides. Transgenic Res. 13: 203-214.
- Hain R, Reif HJ, Krause E, Langebartels R, Kindl H. 1993. Disease resistance results from foreign phytoalexin expression in a novel plant. Nature. 361: 153-156.
- Halls C, Yu O. 2008. Potential for metabolic engineering of resveratrol biosynthesis. Trends Biotechnol. 26: 77-81.
- Jeandet P, Bessis R, Maume BF, Meunier P, Peyron D, Trollat P. 1995. Effect of enological practices on the resveratrol isomer content of wine. J Agric Food Chem. 43: 316-319.
- Jorgensen RA, Cluster PD, English J, Que Q, Napoli CA. 1996. Chalcone synthase co-suppression phenotypes in petunia flowers: comparison of sense vs. antisense constructs and single-copy vs. complex T-DNA sequences. Plant Mol Biol. 31: 957-973.
- Kim SR, Lee J, Jun SH, Park S, Kang HG, Kwon S, An GH. 2003. Transgene structures in T-DNA-inserted rice plants. Plant Mol Biol. 52: 761-773.
- Kohli A, Twyman RM, Abranches R, Wegel E, Stoger E, Christou P. 2003. Transgene integration, organization and interaction in plants. Plant Mol Biol. 52: 247-258.
- Kononov ME, Bassuner B, Gelvin SB. 1997. Integration of T-DNA binary vector ‘backbone’ sequences into the tobacco genome: evidence for multiple complex patterns of integration. Plant J. 11: 945-957.
- Kovalic D, Garnaat C, Guo L, Yan Y, Groat J, Silvanovich A, Ralston L, Huang M, Tian Q, Christian A, Cheikh N, Hjelle J, Padgette S, Bannon G. 2012. The use of next generation sequencing and junction sequence analysis bioinformatics to achieve molecular characterization of crops improved through modern biotechnology. The Plant Genome. 5: 149-163.
- Langcake P, Pryce CA. 1976. The production of resveratrol by Vitis vinifera and other members of Vitaceae as a response to infection or injury. Physiol Plant Pathol. 9: 77-85.
- Leckband G, Lörz H. 1998. Transformation and expression of a stilbene synthase gene of Vitis vinifera L. in barley and wheat for increased fungal resistance. Theor Appl Genet. 96: 1004-1012.
- Rupprich N, Hildebrand H, Kindl H. 1980. Substrate specificity in vivo and in vitro in the formation of stilbenes biosynthesis of rhaponticin. Arch Biochem Biophys. 200: 72-78.
- Saiko P, Szakmary A, Jaeger W, Szekeres T. 2008. Resveratrol and its analogs: defense against cancer, coronary disease and neurodegenerative maladies or just a fad? Mutat Res. 658: 68-94.
- Shou H, Frame BR, Whitham SA, Wang K. 2004. Assessment of transgenic maize events produced by particle bombardment or Agrobacterium-mediated transformation. Mol Breed. 13: 201-208.
- Stark-Lorenzen P, Nelke B, Hänbler G, Mühlbach HP, Thomzik JE. 1997. Transfer of a grapevine stilbene synthase gene to rice (Oryza sativa L.). Plant Cell Rep. 16: 668-673.
- Thomzik JE, Stenzel K, Stöcker R, Schreier PH, Hain R, Stahl DJ. 1997. Synthesis of a grapevine phytoalexin in transgenic tomatoes (Lycopersicon esculentum Mill.) conditions resistance against Phytophthora infestans. Physiol Mol Plant Pathol. 51: 265-278.
- Tonooka Y, Fujishima M. 2009. Comparison and critical evaluation of PCR mediated methods to walk along the sequence of genomic DNA. Appl Microbiol Biotechnol. 85: 37-43.
- Vercauteren J, Castagnino C, Delaunay JC. 2003. Compositions based on Resveratrol. US Patent 6572882. B1.
- Zhang R, Yin Y, Zhang Y, Li K, Zhu H, Gong Q, Wang J, Hu X, Li N. 2012. Molecular characterization of transgene integration by next-generation sequencing in transgenic cattle. PLoS One. 7: e50348
- Zhu YJ, Agbayani R, Jazckson MC, Tang CS, Moore PH. 2004. Expression of the grapevine stilbene synthase gene VST1, in papaya provides increased resistance against diseases caused by Phytophthora palmivora. Planta. 12: 807-812.