Abstract
Confirmation of T-DNA inheritance and stability requires testing of several parameters such as phenotypic performance of transgenic lines, transgene integrity structure and expression stability for at least three consecutive generations, and Mendelian inheritance analysis, which are very important to transgenic breeding and commercialization of genetically modified crops (GMOs). In the present study, we measured nine agronomic and yield-related traits of Iksan526 and confirmed stable phenotypic performance comparing to the donor variety Dongjin over three years. Transgene integrity structure, flanking sequence, functional element copy numbers, absence of backbone sequence and transgene expression were all verified in Iksan526 over three consecutive generations, indicating a stable inheritance of the transgene. In addition, phenotypic and genotypic segregation were analyzed on F2 and F3 populations derived from a cross of Dongjin/Iksan526, and results complied with Mendel’s law of segregation based on single transgene locus. This study demonstrated detailed analyses and confirmation of transgene event Iksan526 which is crucial for commercialization of GMO.
-
Key words: Generations, Mendelian inheritance, Resveratrol, Stability, Transgenic
INTRODUCTION
As one of the most important staple crops, rice is widely consumed for a large part of the world’s human population, especially in Asia. In recent years, most efforts had been initiated to improve rice yields, disease resistance, drought or salinity tolerance and nutritional values by genetic engineering in order to meet population growth, economic benefits and health needs (
Portrykus 2001;
Vasconcelos et al. 2003;
Chen et al. 2009;
Gu et al. 2013).
Baek et al. (2013) developed resveratrol-enriched rice and revealed that its consumption can significantly reduce body weight, blood glucose, triglycerides, total cholesterol and LDL- cholesterol. Resveratrol rice breeding shared the same breeding process to the other transgenic crops which started through
RS3 gene transformation, evaluation and selection, along with complete molecular characterization. Specifically, analysis of the transgenic line event Iksan526 was initiated for commercialization (
Qin et al. 2013).
Submission of biosafety assessment of GM crops is an important requirement for commercialization of GMOs. The general principles for the risk assessment of GM plants and GM-derived food and feeds are described in the EFSA guidance document of the EFSA Scientific Panel on Genetically Modified Organisms and addressed the details of the different components of the risk assessment. One of them is molecular characterization which provides information on the structure and expression of the insert(s) and on the stability of the intended trait(s) (
European Food Safety Authority 2011). In addition, transgene inheritance patterns, whether or not follows Mendelian segregation, need to be observed due to a frequency of 10% non-Mendelian inheritance recorded from transgenic plants produced by
Agrobacterium-mediated transformation (
Yin et al. 2004). In the previous study, the resveratrol transgenic rice line Iksan526 was identified as one intergenic insertion locus by two copies with inverted repeats conjunction structure and expected mRNA expression and resveratrol production (
Qin et al. 2013). However, the complex T-DNA integrity structure of inverted repeats whether it will lead to a instable inheritance from generation to generation not only for the agronomic and yield-related traits, but also T-DNA integrity structure and gene expression, need to be proved. In addition, Mendelian segregation patterns of the hybrid progenies of transgenic line Iksan526 should be analyzed. The results of this study do not only show vital significance to transgene breeding but also as a document to biosafety assessment of the GM rice event Iksan526.
MATERIALS AND METHODS
Agronomic and yield traits measurement and statistical analysis
Nine agronomic and yield traits consisting of culm length, panicle length, panicle number, 100-grain weight, weight per plant, unfilled grain number, filled grain number, grain number per panicle and seed set were measured for 100 samples of transgenic line Iksan526 and the donor variety Dongjin from 2011 to 2013 at Suwon GMO experimental field. Mean values, standard deviations and t-test were performed by means of Microsoft Excel.
Resveratrol analysis by HPLC
A total of 100 mg grain powder from the brown rice of transgenic lines and control Dongjin were obtained every generation for three years. The samples were shake- incubated at 1200 rpm for 30 min at 30°C mixed with 500 ul of 80% (v/v) methanol, and centrifuged at 14000 rpm at 4°C for 5 min. Then the supernants were filtered by 0.2 μm nylon membrane filter for HPLC analysis. Resveratrol standard was obtained from Sigma-Aldrich (#R5010). The separations were carried out using Waters Symmetry C18 column (250 × 4.6 mm, 5μm with guard column) and Shimadzu HPLC system (Kyoto, Japan) with PDA detector. The injection volume, flow rate, column temperature and UV detection are 10 ul, 0.3 ml/min, 35°C and 308 nm, separately. The mobile phase were A: 0.1% formic acid in H2O, and B: 0.1% formic acid in acetonitrile with programs (20min 70:30, 40min 10:90, 50min 10:90, 52min 90:10 and 60min 90:10). Standard calibration with 1.25 ug, 2.5 ug, 5 ug, and 10 ug showed a linear regression with a correlation >0.99.
DNA and RNA sample collection and isolation
T
6, T
7 and T
8 generations of the transgenic resveratrol rice line Iksan526 and its isogenic variety Dongjin were planted at the GMO experimental fields of RDA, Suwon in the rice growing season of 2013. Two lines of isogenic variety ‘Dongjin’ for every three transgenic lines were established with a spacing pattern of 15 cm between plants and 30 cm between rows. Genomic DNAs were isolated from young leaves of rice plants according to the modified cetyltrimethylammonium bromide (CTAB) method (
Chen and Ronald 1999).
For gene expression analysis in different growing stages, leaves from T6, T7, and T8 generations and the donor variety Dongjin were sampled during seedling stage (June 5th), tillering (July 12th), and heading (August 20th). For gene expression analysis in different organs and tissues, the leaf, stem and spikelet were separately sampled from three generations of transgenic lines and the donor variety Dongjin on August 20th, and the seeds were sampled on November 1st. Root samples were also collected at seedling stage. All samples for expression analysis were collected with three repeats. The total RNAs were then extracted by using TRIzol reagent (Invitrogen, UK) according to the manufacturer’s protocol.
Molecular analysis
LA-PCR, southern blot hybridization and RT-PCR were performed to detect T-DNA integrity structure, flanking sequence, functional elements, backbone absence and gene expression following the description in
Qin et al. 2013. Event-specific primers were designed for LA-PCR and RT-PCR as shown in
supplementary Table 1. Specific probes for southern blot hybridization from functional elements and backbone sequence were designed as shown in
supplementary Table 2 and
supplementary Table 3.
Phenotypic and genotypic analyses were performed on F
2 and F
3 populations in 2013 and 2014. A total of 450 F
2 four-leaf old plants were treated with 0.3% of herbicide Basta, and the surviving plants were counted after one week. Plants that have survived were transplanted at Suwon GMO experimental field. For F
3 generation, 9486 heterozygous lines were used for Basta treatment in 2014 (
Supplementary Fig. 4D). For genotypic separation analysis, a total of 300 F
2 and 352 F
3 that have survived from herbicide tolerance screening were PCR using specific primers (
Supplemental Fig. 4A and C). The PCR conditions include a predenaturation step at 94°C for 5 min, followed by 30 cycles of denaturation at 94 °C for 30 s, annealing at 55°C for 30 s, extension for 1 min at 72°C, and final extension for 5 min at 72°C.
RESULTS
Phenotypic traits stability of Iksan526 over generations Agronomic and yield traits stability
Nine agronomic and yield traits were measured over three years. T-test analysis revealed significant difference in 100-grain weight (t = 2.45*) in 2012 and seed set (t = 3.58**) in 2013 between Iksan526 lines and the donor variety, Dongjin (
Table 1). Analysis of three-year combined agronomic and yield data, however, did not show significant difference between the transgenic lines and Dongjin indicating that Iksan526 lines have stable phenotypic performance and yield production over three generations.
Resveratrol, as the end product of T-DNA insertion and expression, was detected in the brown rice grains of three generations of transgenic lines by HPLC analysis while it was undetected in Dongjin. The average values of resveratrol content in Iksan526 were 4.34±0.11 ug/g in T
6, 4.27±0.19 ug/g in T
7, and 4.48±0.03 ug/g in T
8 generations (
Supplemental Fig. 1), illustrating relatively stable inheritance and resveratrol production over generations.
Inserted T-DNA integrity and structure stability
The whole genome sequencing and southern blot hybridization analysis of Iksan526 showed two T-DNAs with head-to-head inverted insertion into the chromosome 12 of Dongjin genome in the previous studies (
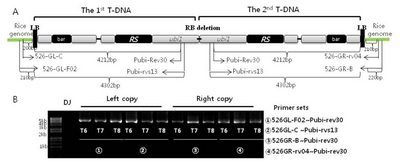
Qin et al. 2013). To illustrate the inserted T-DNA structure through generations, a long fragment of two T-DNAs (4.4–4.5kb) was amplified by LA-PCR. Four primers consisting of 526GL-F02 and 526GL-C from the first T-DNA side and 526GR-B and 526GR-rv04 from the second T-DNA side of rice genomes at insertion site were designed for Iksan526. Two primers of pubi-rev30 and pubi-rvs13 were designed from 4212 and 4302 bp of transformation vector sequence of inserted T-DNA (upstream of the 13 mer of G residues) as shown in
Fig. 1A. By using the four primer sets, LA-PCR was performed for the 1
st and the 2
nd inserted T-DNA of Iksan526 through T
6, T
7 and T
8 generations (
Fig. 1B). The same amplicon size of each primer set ① (4422 bp) and ② (4502bp) observed on the left copy of three generations confirmed the 1
st T-DNA integrity structure and insertion site implying stable inheritance. Similar result was noted for the 2
nd T-DNA (the amplicon size of primer sets ③ was 4412 bp and ④ was 4422 bp). Sequencing analysis of LA-PCR amplicons ② and ③ verified the same flanking sequences at insertion sites of two T-DNAs over T
6, T
7 and T
8 generations (
Supplemental Fig. 2).
T-DNA functional elements insertion stability
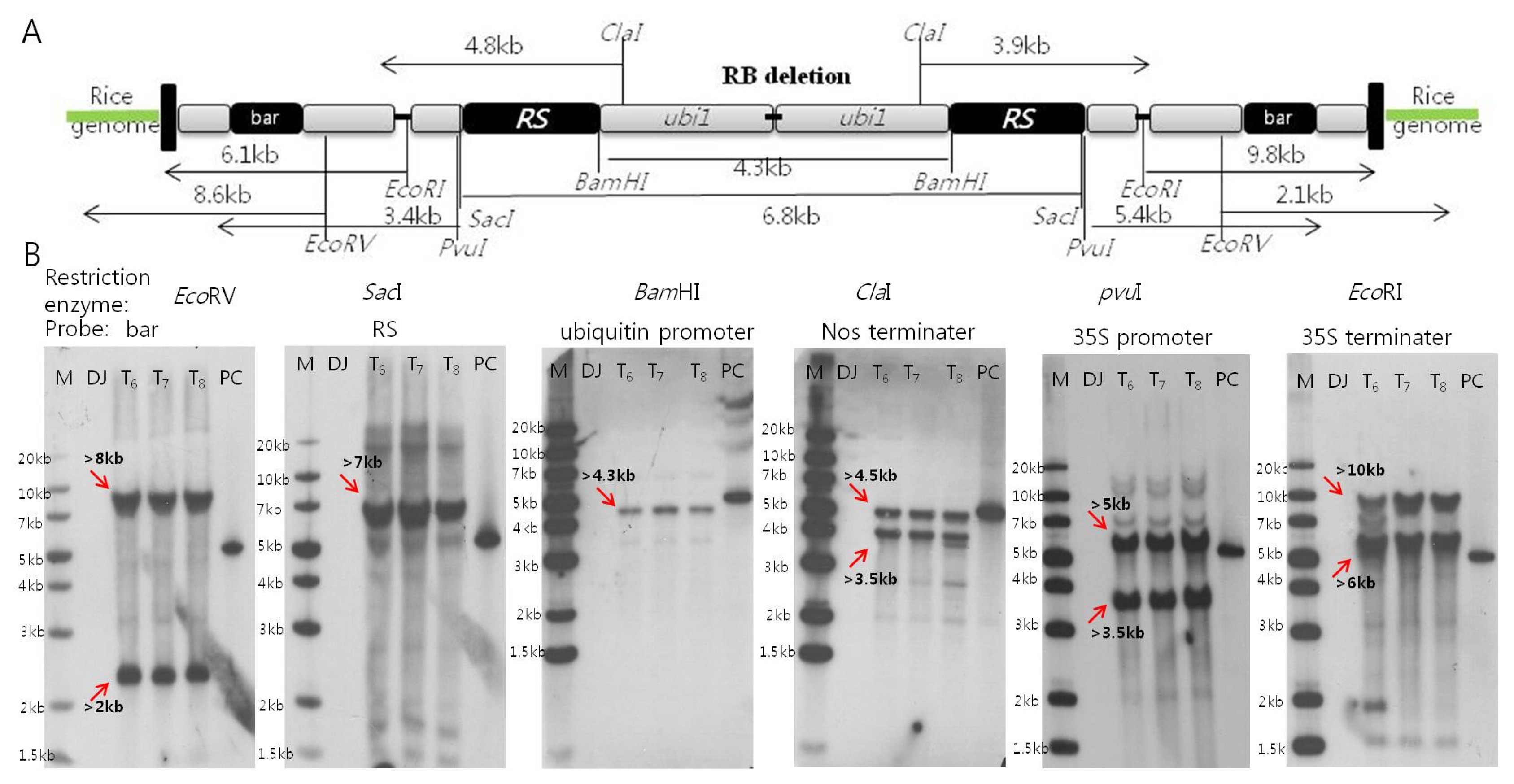
The enzyme restriction site for each functional elements, and the expected band numbers and sizes for inserted two DNAs of Iksan526 were shown in
Fig. 2A. 100 pg positive control derived from the gel elutions from
ScaI digested T-DNA vector (from left border to the upstream about 299 bp of ubiquitin promoter) with a length of 5.04 kb was obtained. The observed band numbers and sizes from southern blot hybridizations were depicted in
Fig. 2B. As expected, two bands were observed for each membrane hybridization: 2 kb/8 kb for
bar, 3.5 kb/4.5 kb for Nos terminator, 3.5 kb/5 kb for 35S promoter, and 6 kb/10 kb for 35S terminator. Each band was observed on the membrane after hybridization with ubiquitin promoter and
RS gene. These results are attributed to the two copies T-DNA insertion with inverted head- to-head structure which is consistent with the expected results. In comparison, there is no signal detected on the negative control Dongjin and the 5.04 kb band on the positive control.
Stability of backbone exclusion
Two kinds of enzymes,
ClaI and
BamHI, were used to digest the DNAs of three generations of Iksan526 (
Supplemental Fig. 3A). If backbone sequences were indeed inserted into rice genomes, two bands should be detected on the membrane after the hybridization of
ClaI digested DNAs with backbone specific probes which overlapped at 6.226 kb backbone sequence. Moreover, one band should be detected on membrane after hybridization of
BamHI digested DNAs with corresponding specific backbone probes. However, none of the bands produced indicates the backbone in rice genome sequence of Iksan526 as shown in
Supplemental Fig. 3C. The same performances of three generations suggested that genome of Iksan526 did not carry the backbone sequence, while it stably passed on the transgene over three generations.
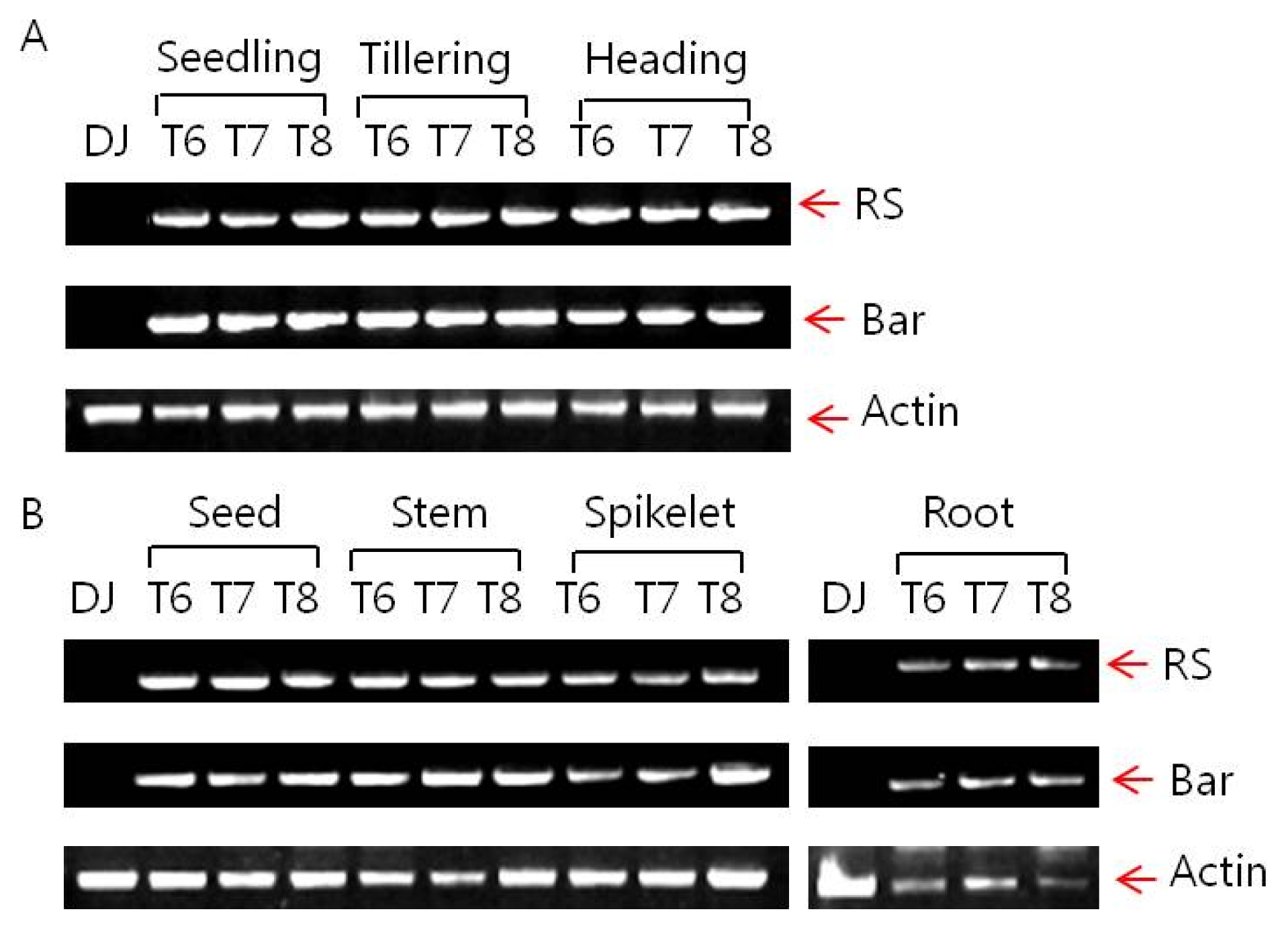
Stability of foreign gene expression
The inserted T-DNA is composed of two expression cassettes of Ubi:
RS and 35S:
bar. Expressions of
RS and
bar genes were determined by reverse-transcript PCR as shown in
Fig. 3. Stability of expressions of these genes were verified regardless of growing stages such as seedling, tillering and heading, or generations (T
6, T
7 and T
8).
Mendelian inheritance analysis for transgene
Phenotypic and genotypic screening were carried out on two consecutive populations F
2 and F
3 derived from a cross between Dongjin and Iksan526 (T
6), to confirm one locus T-DNA insertion in accordance with Mendel’s law of segregation in hybrid progeny population. A phenotypic segregation ratio of 3:1 was observed in F
2 population after treatment with Basta where 321 plants survived and 129 died, and in F
3 population where 7100 plants survived and 2386 died (
Table 2). PCR assay revealed 92 homologous and 208 heterozygous in F
2 population and 120 homologous and 232 heterozygous in F
3 population (
Supplemental Fig. 4B and E) which fit the 1:2 segregation ratio. Results of genetic analysis of F
2 and F
3 populations indicated that the phenotypic and genotypic segregation ratios of transgene were consistent to Mendel’s law of segregation on single gene locus.
DISCUSSION
Whether or not a transgenic trait has a predicted inheritance and expression pattern over multi-generations, conformation to Mendelian segregation is crucial to transgene breeding and GMOs commercialization (
Zhong 2001). In addition, GMOs with inverted repeated structure transgenes usually show the instability of DNA integrity leading to transgene inactivation or silencing during the development from generation to generation (
Muskens et al. 2000;
Lechtenberg et al. 2003). Thus, based on the prerequisite of a characterization of two head-to-head copies of one locus by intergenic insertion and high gene expression in the previous study (
Qin et al. 2013), the confirmation and document of phenotypic performance, integrity structure, functional expression for at least three generations (years), and Mendelian inheritance patterns through two hybrid progeny population is the main goal in the present study.
In summary, agronomic and yield-related traits of Iksan526 were evaluated and compared to those of Dongjin for three years and stable phenotypic performance was confirmed. Moreover, stability of integrity structure stability and expression were verified through analyses of flanking sequence, inserted T-DNA sequence, functional element copy numbers and size, backbone absence confirmation, mRNA expression, and end product of resveratrol content over three successive generations. A similar research was carried out on PAC rice (
Oh et al. 2012), but our study presented more detailed information especially on the complex T-DNA integrity structure of the event Iksan526.
Transgenes may not always follow Mendelian segregation in some cases due to endogenous locus disruption and interference with the endogenous biochemical pathways by biochemical product (
Pawlowski and somers 1996;
Hood et al. 1997;
Yin et al. 2004), however, such is not the case in Iksan526. Mendelian inheritance analysis revealed that transgenes of Iksan526 followed Mendelian segregation. Taken together, the event Iksan526 is stably inherited and thus fits into biosafety assessment of GMOs.
Supplementary materials
Supplemental Table 1The primer sequence information of LA-PCR.
Supplemental Table 1
|
Primers |
Primer sequence |
Primer location |
|
Primers of flanking regions from rice genomes |
|
|
526GL-F02 |
CAATTCGGTCTCATTAGCCTACCTCA |
210bp from 1st T-DNA on chr. 12 |
|
526GL-C |
GTGCTTGTTGTCTTCTCTTACG |
200bp from 1st T-DNA on chr. 12 |
|
526GR-B |
GATATAGACCTAGCACAGAAAGG |
220bp from 2nd T-DNA on chr. 12 |
|
526GR-rv04 |
CAGAAAGGACACATGTCCCGTTGAGAGA |
210bp from 2nd T-DNA on chr. 12 |
|
|
Common primer from plasmid vector (pSB2220) |
|
|
Pubi-rvs13 |
CCCCCCTCTCTACCTTCTCTAGATC |
4302bp from left border |
|
Pubi-rev30 |
TTAGATCCGTGCTGCTAGCGTTC |
4212bp from left border |
Supplemental Table 2The primer sequence information of specific probes from six functional elements and inner control actin for RT-PCR.
Supplemental Table 2
|
Functional elements |
Primer names |
Primer sequences |
Expected size |
Specific enzymes |
|
35S terminater |
35s-ter-f |
ATTCGGGGGATCTGGATTTTAGTACT |
187bp |
EcoRI |
|
35s-ter-r |
CGAGTTTCTCCATAATAATGTGTGAG |
|
35S promoter |
35s-pro-f |
GGTAGACTCGAGAGAGATAGATTTG |
152bp |
PvuI |
|
35s-pro-r |
TGATGTGATATCTCCACTGACGTA |
|
Nos-terminater |
Nos-ter-f |
GCGATAATTTATCCTAGTTTGCGCGCTA |
244bp |
ClaI |
|
Nos-ter-r |
CGGAGCTCGAATTTCCCCGATCGTTC |
|
Ubiquitin promoter |
ubi-pro-f |
GTCCTGTTGTCAAAATACTCAATTGTC |
276bp |
BamHI |
|
ubi-pro-r |
AGTGCCAAGCTTGCATGCCTGCAGTGC |
|
Bar |
bar-for |
GAAGTCCAGCTGCCAGAAAC |
400bp |
EcoRV |
|
bar-rev |
AAGCACGGTCAACTTCCGTA |
|
Resveratrol synthase |
Rs-for |
GCATTGTTTGCGGATGGAGCTG |
500bp |
SacI |
|
Rs-rev |
TTATATGGCCACACTGCG |
|
|
Actin |
fw |
CTGCTATGTACGTCGCCATC |
440bp |
- |
|
rv |
GTCTCATGGATACCCGCAG |
Supplemental Table 3The primer sets for confirmation on absence of T-DNA vector backbone sequence in transgenic rice line Iksan526.
Supplemental Table 3
|
Backbone |
Primer name |
Primer sequence |
Expected size |
|
Pair1 |
rsbcbn-1-fw |
GATACAGGCAGCCCATCAGTC |
753bp |
|
rsbcbn-1-rv |
TTTCAAGGGCTTTGTTCATCTTCA |
|
pair2 |
rsbcbn-2-fw |
CACTTTGAACGGCATGATGG |
797bp |
|
rsbcbn-2-rv |
CAGTATTTGTATCTGCGCTCTG |
|
pair3 |
rsbcbn-3-fw |
GCGGTGGTTTGTTTGCCGGATCAAGAGC |
809bp |
|
rsbcbn-3-rv |
GATTGTACTGAGAGTGCACCATA |
|
pair4 |
rsbcbn-4-fw |
GCGAGTCAGTGAGCGAGGAAGC |
944bp |
|
rsbcbn-4-rv |
GTGGCAGCGCCGGAGAGTTC |
|
pair5 |
rsbcbn-5-fw |
CCGCCTCCTCCTTCAAATCGTA |
794bp |
|
rsbcbn-5-rv |
CGCCCAAGGGCGACGAGCAA |
|
pair6 |
rsbcbn-6-fw |
CATGATGCTGCGACTATCG |
781bp |
|
rsbcbn-6-rv |
AACCGTTCTTGAATCAGAACC |
|
pair7 |
rsbcbn-7-fw |
TAACTCAAATGAGTTTTGATTTG |
769bp |
|
rsbcbn-7-rv |
GCCAAGCTGGCGGCCTGG |
|
pair8 |
rsbcbn-8-fw |
CATCACCTTTTTAGACGG |
771bp |
|
rsbcbn-8-rv |
GCACCAGGCGCGACCGC |
|
pair9 |
rsbcbn-9-fw |
CACTGTCACAACGTCGCCAG |
562bp |
|
rsbcbn-9-rv |
GGTTTATCCGTTCGTCCA |
Supplemental Fig. 1
Resveratrol contents analysis for the brown rice grains of three generations for transgenic rice line Iksan526 and their non-detect control Dongjin. A. Resveratrol standard of 10ug; B. A mixture of 5ug resveratrol standard and the resveratrol extracts from T6 seed of Iksan526; C. The donor variety Dongjin as a non-detect negative control; D, E and F indicated the resveratrol detection from T6, T7 and T8 brown rice grains of Iksan526, respectively.
Supplemental Fig. 2
Flanking sequence of T-DNA insertion over three generations by LA-PCR.
Supplemental Fig. 3
Backbone sequence absence confirmation for three generations of Iksan526; A. The restriction enzyme sites of plasmid construction vector; B. The expected and observed band numbers by two enzymes after hybridization; C. Confirmation on absence of plasmid vector backbone sequence of three generations of transgenic rice line Iksan526 by using southern blot hybridization. * M: Generuler 1kb plus ladder; NC: Dongjin (negative control); T6~8: generation of Iksan526; PC: Positive control (psb2220-5.04kb).
Supplemental Fig. 4
Phenotypic and genotypic separation analysis for F3 population derived from a cross of Dongjin/Iksan526 (T6). A. A strategy of primer design for genotypic separation analysis; B. PCR amplification to differentiate heterozygous and homozygous genotypes; C. Specific primer sequence and location on rice chromosome; D. 4-leaf old plants before and after basta treatment; E. Genotypic separation of 352 F3 population.
ACKNOWLEDGMENTS
This work was supported by grant PJ009528 from the Biogreen 21 R&D Program, Rural Development Administration, Republic of Korea.
Fig. 1Confirmation of T-DNA integrity structure and generational stability of Iksan526. A. The basic molecular information of inserted T-DNA and primer design for LA-PCR; B. T-DNA flanking sequence analysis through three generations by LA-PCR.
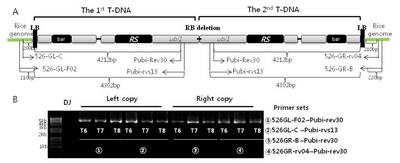
Fig. 2Confirmation on copy numbers of six functional elements for Iksan526 using Southern blot hybridization. A. The restriction enzyme sites of inserted T-DNA and their expected band sizes; B. Copy numbers of six functional elements through three generations. * M: Generuler 1 kb plus ladder; NC: Dongjin (negative control); T6~8: generation of Iksan526; PC: Positive control (psb2220–5.04kb)
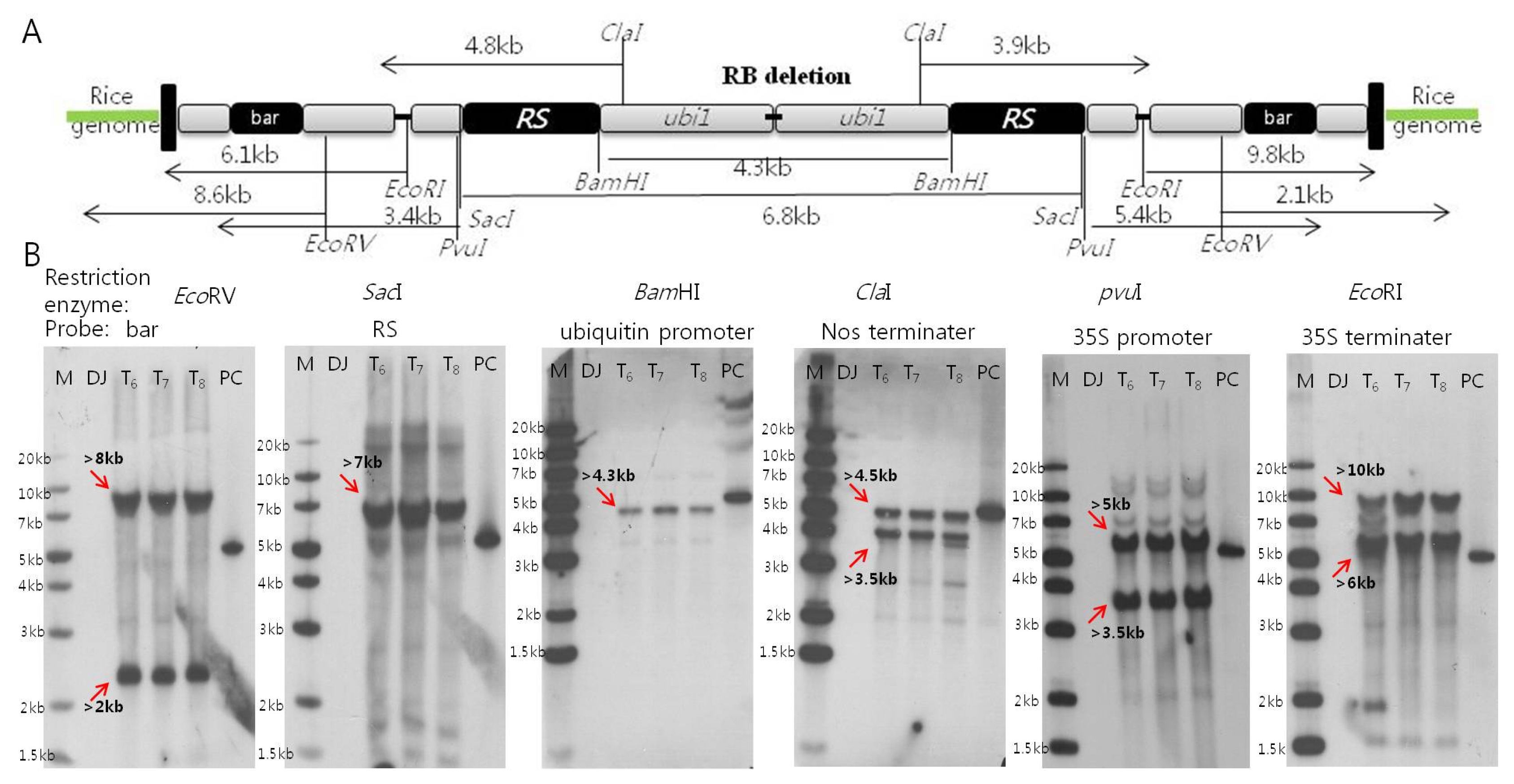
Fig. 3Inserted resveratrol synthase gene and bar gene expression of Iksan526 by RT-PCR. A. Gene expression in different growing stage; B. Gene expression in different tissues.
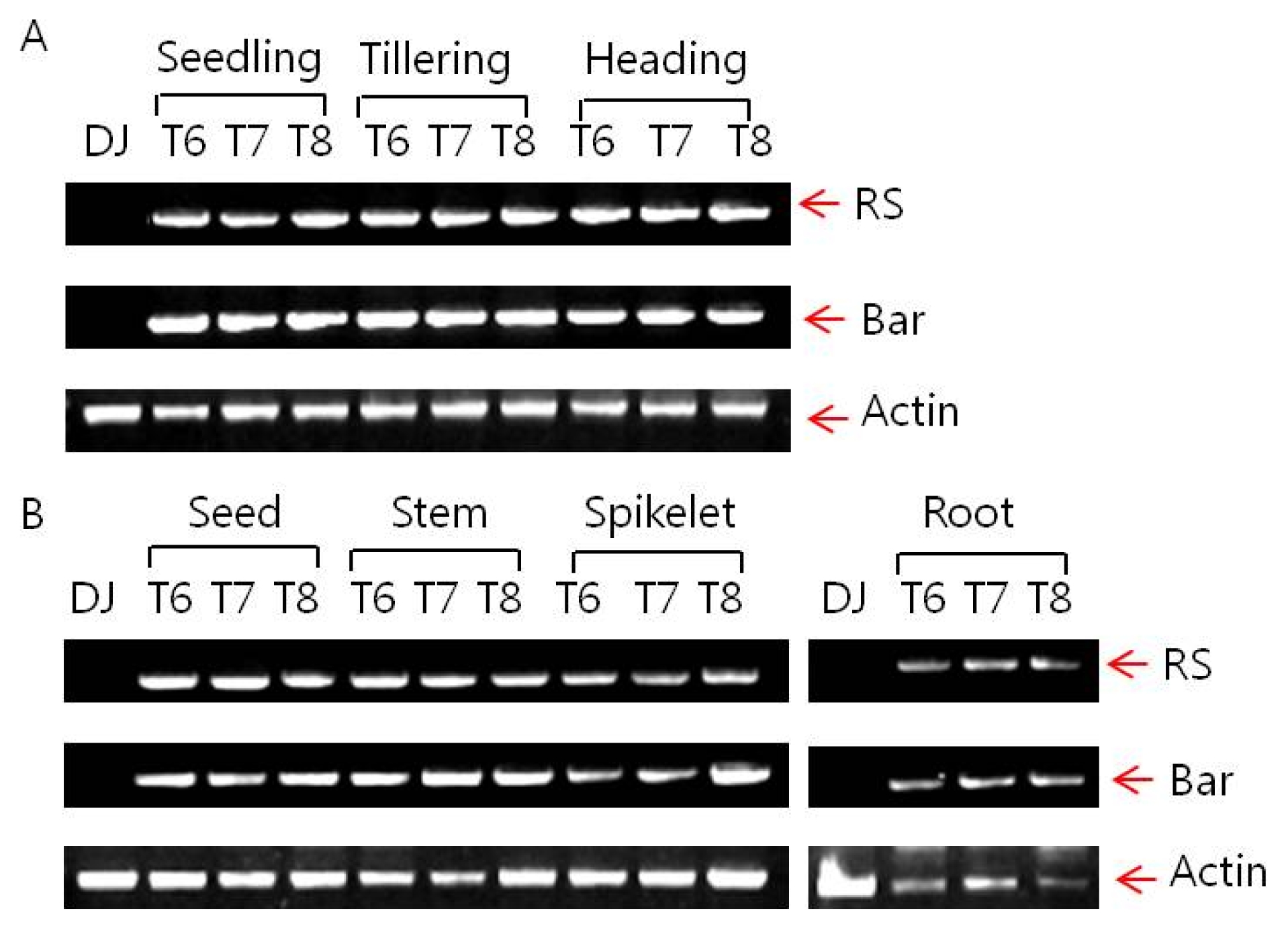
Table 1Comparison and t-test analysis of agronomic and yield traits over three years (generations) of resveratrol transgenic rice Iksan526 and its donor variety Dongjin.
Table 1
|
Variety |
Years |
Culm length |
Panicle length |
Panicle number |
Weight/plant (g) |
100-grain weight (g) |
|
Iksan526 |
2011 |
94.15±4.69 |
20.74±1.85 |
11.17±2.13 |
30.63±7.16 |
2.73±0.17 |
|
Dongjin |
96.60±4.27 |
20.30±1.57 |
13.10±1.85 |
34.28±7.93 |
2.61±0.10 |
|
t value |
0.30 |
0.24 |
0.85 |
0.14 |
0.77 |
|
|
Iksan526 |
2012 |
83.24±3.89 |
22.02±1.71 |
8.84±2.33 |
26.31±8.98 |
2.99±0.07 |
|
Dongjin |
86.32±2.38 |
21.26±1.10 |
8.89±2.40 |
27.53±8.26 |
2.83±0.09 |
|
t value |
0.83 |
0.47 |
0.02 |
0.14 |
2.15* z)
|
|
|
Iksan526 |
2013 |
81.66±2.64 |
20.59±1.28 |
7.80±2.15 |
15.36±4.81 |
2.79±0.06 |
|
Dongjin |
83.89±2.03 |
21.30±0.95 |
7.80±2.12 |
18.85±5.37 |
2.74±0.07 |
|
t value |
0.92 |
0.60 |
0.00 |
0.74 |
0.73 |
|
|
Iksan526 |
3Y |
85.60±6.30 |
21.37±1.78 |
8.92±2.53 |
23.10±9.55 |
2.86±0.15 |
|
Dongjin |
87.78±5.09 |
21.08±1.26 |
9.11±2.75 |
24.57±8.27 |
2.76±0.11 |
|
t value |
0.36 |
0.17 |
0.07 |
0.16 |
0.71 |
|
Variety |
Years |
Unfilled grain number |
Filled grain number |
Seed set (%) |
Grain number/panicle |
Resveratrol (+ Piceid) (ug/g) |
|
Iksan526 |
2011 |
102.35±48.88 |
1121.04±248.18 |
91.52±3.84 |
101.28±17.52 |
4.34±0.11 |
|
Dongjin |
111.10±45.82 |
1313.96±311.99 |
92.12±3.18 |
100.69±19.07 |
- |
|
t value |
0.01 |
0.01 |
0.18 |
0.00 |
- |
|
|
Iksan526 |
2012 |
180.46±81.28 |
879.91±298.24 |
83.03±4.67 |
119.16±17.95 |
4.27±0.19 |
|
Dongjin |
89.42±33.71 |
974.79±298.06 |
91.46±2.80 |
119.71±13.49 |
- |
|
t value |
1.20 |
0.32 |
1.90 |
0.03 |
- |
|
|
Iksan526 |
2013 |
309.57±94.91 |
552.23±176.02 |
63.75±5.37 |
70.64±10.37 |
4.48±0.03 |
|
Dongjin |
145.30±71.19 |
687.68±199.60 |
82.67±6.20 |
87.88±7.23 |
- |
|
t value |
1.88 |
0.78 |
3.58** y)
|
1.82 |
- |
|
|
Iksan526 |
3Y |
212.72±114.19 |
806.77±328.59 |
59.14±31.03 |
97.43±26.64 |
4.36±0.11 |
|
Dongjin |
116.98±58.63 |
892.45±304.80 |
72.31±33.22 |
101.94±18.97 |
- |
|
t value |
0.90 |
0.26 |
0.42 |
0.18 |
- |
Table 2Phenotypic and genotypic separation ratio at F2 and F3 populations from a cross Dongjin/Iksan526.
Table 2
|
Population |
Phenotype/genotype |
Treatment |
Expected separation ratio |
Observed separation ratio |
Possibility |
|
Dongjin/Iksan526 F2 population |
Phenotypic separation |
Basta |
3 : 1 |
Resistant 321 |
Susceptible 129 |
>0.90 |
|
Genotypic separation |
PCR |
1 : 2 |
Homologous 92 |
Heterozygous 208 z)
|
>0.90 |
|
Dongjin/Iksan526 F3 population |
Phenotypic separation |
Basta |
3 : 1 |
Resistant 7100 |
Susceptible 2386 |
>0.90 |
|
Genotypic separation |
PCR |
1 : 2 |
Homologous 120 |
Heterozygous 232 |
>0.90 |
References
- Baek SH, Shin WC, Ryu HS, Lee DW, Moon EJ, Seo CS, Hwang E, Lee HS, Ahn MH, Jeon YJ, Kang HJ, Lee SW, Kim SY, DSouza R, Kim HJ, Hong ST, Jeon JS. 2013. Creation of resveratrol-enriched rice for the treatment of metabolic syndrome and related diseases. PLoS One. 8: e57930
- Chen H, Lin YJ, Zhang QF. 2009. Review and prospect of transgenic rice research. Chinese Sci Bull. 54: 4049-4068.
- Chen DH, Ronald PC. 1999. A rapid DNA minipreparation method suitable for AFLP and other PCR applications. Plant Mol Biol Rep. 17: 53-57.
- European Food Safety Authority (EFSA)2011. EFSA Panel on Genetically Modified Organisms (GMO). Science Opinion on Guidance for risk assessment of food and feed from genetically modified plants. EFSA J. 9: 2150.
- Gu JF, Qiu M, Yang JC. 2013. Enhanced tolerance to drought in transgenic rice plants overexpressing C4 photosynthesis enzymes. Crop J. 1: 105-114.
- Hood E, Witcher D, Maddock S, Meyer T, Baszczynski C, Bailey M, Flynn P, Register J, Marshall L, Bond D, Kulisek E, Kusnadi A, Evangelista R, Nikolov Z, Wooge C, Mehigh R, Herman R, Kappel W, Ritland D, Li CP, Howard J. 1997. Commercial production of avidin from transgenic maize: characterization of transformant, production, processing, extracting and purification. Mol Breed. 3: 291-306.
- Lechtenberg B, Schubert D, Forsbach A, Gils M, Scjmidt R. 2003. Neither inverted repeat T-DNA configurations nor arrangements of tandemly repeated transgenes are sufficient to trigger transgene silencing. Plant J. 34: 507-517.
- Muskens MWM, Vissers APA, Mol JNM, Kooter JM. 2000. Role of inverted DNA repeats in transcriptional and post-transcriptional gene silencing. Plant Mol Biol. 43: 243-260.
- Oh SD, Park SY, Sohn SI, Kim JK, Park JS, Park SK, Cho HS, Ahn BO, Lee KJ. 2012. Evaluation and Assessment of Biosafety Carotenoid-biofortified Rice (PAC): Progeny Stability Study for Four Generations. Korean J Breed Sci. 44: 579-587.
- Pawlowski WP, Somers DA. 1996. Transgene inheritance in plants genetically engineered by microprojectile bombardment. Mol Biotechnol. 6: 17-30.
- Potrykus I. 2001. Gold rice and beyond. Plant Physiol. 125: 1157-1161.
- Qin Y, Ahn HI, Kweon SJ, Baek SH, Shin KS, Woo HJ, Cho HS, Lee JH, Lim MH. 2013. Molecular characterization of transgenic rice producing resveratrol. Plant Breed Biotech. 1: 406-415.
- Vasconcelos M, Datta K, Oliva N, Khalekuzzaman M, Torrizo L, Krishnan S, Oliveira M, Goto F, Datta SK. 2003. Enhanced iron and zinc accumulation in transgenic rice with the ferritin gene. Plant Sci. 164: 371-378.
- Yin ZM, Plader W, Malepszy S. 2004. Transgene inheritance in plants. J Appl Genet. 45: 127-144.
- Zhong GY. 2001. Genetic issues and pitfalls in transgenic plant breeding. Euphytica. 118: 137-144.