Abstract
Ascorbate peroxidases (APXs) are enzymes that detoxify peroxides such as hydrogen peroxide using ascorbate. They are distributed as isoenzymes in distinct cellular compartments, such as the cytosol, mitochondria and peroxisomes, where they play essential roles in scavenging reactive oxygen species (ROS) and protecting cells against the toxic effects of these species in higher plants, algae, euglena and other organisms. APXs also respond to environmental stresses, such as salinity and drought, and rice ascorbate peroxidase 1 (OsAPx1) participates in salinity tolerance. However, it is still unclear how OsAPx1 is involved in growth and development before and after flowering. Here, we show that OsAPx1 plays an important function in seed development, including fertilization. Proteomic analysis and quantitative RT-PCR showed that protein and mRNA levels of OsAPx1 were much higher in immature seeds than in mature seeds. Plant height and grain size in an Osapx1 mutant were almost the same as those in wild-type plants. However, about 58% of the seeds were aborted in the Osapx1 mutant, although the mutant was capable of normal flowering. Our results suggest that rice ascorbate peroxidase functions both as a regulator of seed development and as a scavenger of ROS.
-
Key words: Ascorbate peroxidase, OsAPx1, Seed development, Rice
INTRODUCTION
Rice (
Oryza sativa) is a model grain crop and its grain shape plays a major role in consumer preferences in different countries and populations. It is known that the filled grain ratio is strongly affected by biotic and abiotic stresses, whereas the total grain weight is mostly or completely governed by genetic factors (Sakamoto and Matsuoka 2008). Seed size is a major determinant of seed yield, cooking and eating quality, and also affects milling and processing quality to some extent (
Fan et al. 2006;
Bai et al. 2010;
Song et al. 2007). In addition to these direct components, traits such as plant height and tillering are also crucial for seed yield in modern agriculture.
Seed development is affected by various factors. One is genetic or epigenetic. It has been reported that grain length and width may be regulated by multiple genes (
Kesavan et al. 2013). For example, GS3 (grain size), encoding a trans-membrane protein, is a major QTL linked to grain length and weight. It is also a minor QTL for grain width and thickness in rice (Takano-Kai
et al. 2009). The loss-of-function mutant
GW2 (grain width and weight) leads to increased cell number, a wider spikelet hull, and an accelerated grain milk-filling rate, which increases grain width, weight, and yield (
Song et al. 2007). QTL
SW5 (seed width) or
GW-5 (grain width) mutants result in defects in cell division during seed development and also increases the spikelet volume and cell number in the outer glume (
Shomura et al. 2008;
Weng et al. 2008). Grain incomplete filling 1 (
GIF1) encodes a cell-wall invertase that is required for carbon partitioning during early grain filling (
Wang et al. 2008).
Epigenetic modifications of regulated genes are necessary to control seed development and seed size (Kesavan
et al. 2012). Evidence from
Arabidopsis interploidy crosses suggest that maternal methylation makes viable seeds larger than normal, whereas paternal methylation results in smaller seeds. Similar results were obtained by crossing wild-type and transgenic DNA methyltransferase (
MET1) RNAi plants (
Xiao et al. 2006). Therefore, mutations in imprinting genes usually lead to seed abortion and ultimately result in a smaller seed size (
Kohler et al. 2005).
The second factor is hormones. The effects that plant hormones exert on seed size development can be seen in mutants defective in plant hormone biosynthesis or perception. Arabidopsis
ARF2 (
Auxin Response Factor 2) may have similar functions to rice
GW2 in terms of seed size and weight (
Schruff et al. 2005). The BR biosynthesis pathway also affects crop grain size (
Tanabe et al. 2005). For example, brassinosteroid (BR) biosynthesis-defective rice dwarf mutants, i.e.,
dwarf1 (d1), dwarf2 (d2) dwarf11 (d11),
BR-deficient dwarf1, and
BR-deficient dwarf2 share a similar seed phenotype, producing grains with a significantly reduced seed length. The phenotype of the
dwarf1 mutant is gibberellic acid deficient, while
DWARF1 is the only rice gene encoding a prototypical heterotrimeric G protein α-subunit, which probably functions in the GA signal transduction pathway (Hong
et al. 2005). Cytokinin may also have an important role in seed size and weight. For example,
Arabidopsis transgenic plants that overexpress cytokinin oxidase-dehydrogenase genes,
AtCKX1 and
AtCKX3, also have a reduced seed set, but the seeds that form are enlarged and about twice the weight of wild-type seeds. Abscisic acid (ABA) is also involved in seed development. In barley (
Hordeum vulgare), the malting quality-related genes
HvFIE and
HvE(Z) are induced by ABA (
Kapazoglou et al. 2010). Unlike other hormones, the newly identified strigolactone inhibits shoot branching, thereby reducing spikelet number and final grain yield.
The third factor is abiotic stress. It is well accepted that increased temperature and CO
2 regulate the grain-filling process via genetic and environmental pathways (
Yang and Zhang 2006). An increase in the optimum mean temperature during grain filling may result in infertility and thence poor seed set, and also results in decreased grain size and weight (
Yang and Zhang 2006). In addition, it gives rice grains a chalky appearance, which is a serious problem because chalky grains have an inferior polishing capacity and lower palatability (Sakamoto and Matsuoka 2008;
Peng et al. 2004). Furthermore, high temperature inhibits dry matter accumulation, resulting in lower grain size and weight in cereals (
Peng et al. 2004).
ROS, such as the superoxide radical (O
2−), H
2O
2, and the hydroxyl radical, are produced as a consequence of the univalent reduction of molecular oxygen in metabolic reactions, an intrinsic feature of aerobic life. In plant cells, the intracellular level of ROS must be tightly regulated since ROS also act as signaling molecules in several physiological processes, such as cell cycle, stress perception, gene regulation, programmed cell death (PCD), hypersensitive response, and senescence (
Dat et al. 2000;
Orozco-Cárdenas et al. 2001;
Mullineaux and Karpinski 2002;
Vandenabeele et al. 2003). Control of intracellular ROS levels is a very complex process and involves a large network of genes, whose principal function is to avoid cellular damage that could result in oxidative stress and disturbances in cellular redox homeostasis (
Mittler et al. 2004). The ROS-scavenging system is responsible for removing free radicals and keeping the cellular steady-state level of ROS under tight control. This system consists of low-molecular weight compounds, including ascorbic acid, glutathione, tocoferol, and carotenoids, and several enzymes such as superoxide dismutases (SOD), ascorbate peroxidases (APx), catalases (CAT), glutathione peroxidases (GPx), and peroxidases (Prx) (
Halliwell and Gutteridge 1999;
Scandalios 2002;
Mittler et al. 2004).
APx has a key role in scavenging ROS and in protecting cells against the toxic effects of ROS in higher plants, algae, and Euglena (
Shigeoka et al. 1980,
2002). The expression of APx encoding genes is modulated by diverse environmental stimuli such as water and salt stresses, high temperatures, freezing, pathogen attack, H
2O
2 treatment, and abscisic acid (
Zhang et al. 1997;
Mittler et al. 1998;
Yoshimura et al. 2000;
Sato et al. 2001;
Shigeoka et al. 2002;
Agrawal et al. 2003;
Menezes-Benavente et al. 2004a,
b). Despite its obvious importance as a model cereal plant, little is known about the function of rice APx during development, in particular during seed development.
Here, we show that OsAPx1 is highly expressed in developing immature seed and that its mutant produces abnormal panicles and aborted seeds. Our data provide valuable information that ascorbate peroxidase plays an important function in not only protecting cells from ROS but also in seed development.
MATERIALS AND METHODS
Plant materials
The T-DNA insertion mutant rice, PFG_2B-70163, was kindly provided by Dr. Gynheung An at POSTECH (Pohang, Korea). T-DNA insertion mutant rice was generated from the cultivar Dongjinbyeo (wild type). To identify T-DNA insertion sites within the
OsAPx1 locus, primers were designed for T-DNA border sequences and the flanking region of the
OsAPx1. The loci of inserted T-DNA in the mutant line were identified by TAIL-PCR according to previously described methods (
Liu and Whittier 1995). The insertion sites were amplified by PCR using primers that specifically bind to the border sequence and flanking region of the T-DNA. Selected homozygote lines were used to produce progeny for this study.
To examine the phenotypes of the Osapx1 mutant and wild-type, plants were grown in the field for 90 days and then whole plants were photographed. To check the weight of the panicle and the number of normal and aborted seeds of the Osapx1mutant, panicles and grains were harvested from wild-type and Osapx1 mutant plants at 55 days after flowering.
Two-dimensional Gel Electrophoresis and Imaging Analysis
Seed proteins were extracted from immature (15 DAF) and mature (50 DAF) seeds of the wild type and separated by two-dimensional gel electrophoresis (2-DE). Sample preparation was performed by the following method. Whole seeds chilled in liquid nitrogen were homogenized using a mortar and pestle. The extraction buffer (7M urea, 2 M thiourea, 4% (w/v) 3-[(3-cholamidopropy) dimethyammonio]-1-propanesulfonate (CHAPS), 1% (w/v) dithiothreitol (DTT), 2% (v/v) pharmalyte, and 1 mM benzamidine) was added to powdered samples and incubated for 1 h at room temperature. After centrifugation at 15,000 6g for 1 h at 15 uC, the soluble fraction was withdrawn using 2-DE. The proteins were loaded onto an IPG dry strip equilibrated with another buffer (7 M urea, 2 M thiourea, 2% CHAPS, 1% DTT, and 1% pharmalyte). Isoelectric focusing was performed at 20 uC using a Multiphor II electrophoresis unit and an EPS 3,500 XL power supply (Amersham Biosciences) under the following conditions: linear increase from 150 to 3,500 V for 3 h; constant 3,500 V to reach 96 kV/h. Then, the strips were immersed for 10 min in solution (50 mM Tris-Cl (pH 6.8), 6 M urea, 2% SDS, and 30% glycerol) and transferred to SDS-polyacrylamide gels (20,624 cm, 10–16%). SDS-PAGE was carried out at 20°C with 1,700 V/h using a Hoefer DALT 2D system (Amersham Biosciences) according to the manufacturer’s instructions. After electrophoresis, gels were stained with silver nitrate as previously described (
Oakley et al. 1980). From the stained gels, digitized images were acquired and quantitatively analyzed using the PDQuest (version 7.0, Bio-Rad) software. The quantity of each spot was normalized by total valid spot intensity. Protein spots to analyze with MALDI-TOF were selected based on a more than two-fold intensity increase or decrease as compared with control samples. Enzymatic Digestion of Proteins in Gels and MALDI-TOF Analysis Protein spots were enzymatically digested in-gel using modified porcine trypsin as previously described (
Shevchenko et al. 1996). Gel pieces were cleaned with 50% acetonitrile followed by rehydration with trypsin (8–10 ng/mL) and incubated for 8–10 h at 37°C. The reaction was terminated by addition of 5 mL of 0.5% trifluoroacetic acid. Tryptic peptides were combined from the aqueous phase of gel extractions with 50% aqueous acetonitrile through several rounds. After concentration, the peptides were desalted using C18ZipTips (Millipore, MA, USA) and eluted in 1–5 mL of acetonitrile. Purified peptides were mixed with an equal volume of α-cyano-4-hydroxycinnamic acid in 50% aqueous acetonitrile, and 1 mL was spotted onto a target plate. Protein analyses were performed using an Ettan MALDI-TOF (Amersham Biosciences). Peptides were evaporated with an N2 laser at 337 nm and accelerated with a 20 kV injection pulse for time-of-flight analysis. Each spectrum was a cumulative average of 300 laser shots. The database search program ProFound, developed by Rockefeller University (
http://129.85.19.192/profound_bin/WebProFound.exe), was used to identify proteins by peptide mass fingerprinting. To determine the confidence of identified proteins, the Z-score, MASCOT score, and sequence coverage were used as the determining criteria. The proteins with a Z-score $1.28 were considered to be successfully identified (Z-score $1.28 indicates that the database match is nonrandom with a 90% probability). Proteins that were matched with a MASCOT score of greater than 64 were considered to be significant (p=0.05). The sequence coverage of matching peptides should not be less than 15%. Spectra were calibrated against trypsin auto-digestion ion peaks m/z (842.510, 2211.1046), which serve as internal standards.
Total RNA was extracted from plants at different developmental stages and from different tissues to investigate the spatial and temporal expression patterns of OsAPX1 according to a method previously described (
Altenbach and Howell 1981). Total RNA was quantified and first-strand cDNA synthesis was performed using 5 μg of total RNA and the iScriptTM cDNA Synthesis Kit (Bio-Rad). Then, an equal volume of cDNA was amplified by quantitative real-time PCR (MyiQ, Bio-Rad) according to the manufacturer’s protocol. Specific primers and template cDNA were combined with 25 μL iQTM SYBRH Green Super Mix (Bio-Rad), and reactions were performed as follows: 50°C for 2 min; 95°C for 10 min; and 40 cycles of 95°C for 15 sec and 60°C for 1 min. The CT values of the target genes were normalized to the CT value of OsActin (Accession No. X16280) and analyzed using iCycler IQTM software (Bio-Rad). RT-PCR primers were designed using Primer3 (
http://frodo.wi.mit.edu/cgi-bin/primer3/primer3.cgi), and their specificity was verified by cloning into the pGEM T-Easy vector (Promega) and sequencing with an ABI 3730xl DNA Analyzer (Applied Biosystems).
The protein sequences of ascorbate peroxidases of various species that were homologous to OsAPx1 were aligned by using the CLUSTALW2 program (
http://www.ebi.ac.uk/Tools/msa/clustalw2/) and then manually adjusted to minimize gaps by using MEGA5 software (
Tamura et al. 2011). A taxonomic lineage was generated by the neighbor-joining method with 1,000 bootstrap replicates using MEGA5 software.
RESULTS
OsAPX1 accumulates in mature seed
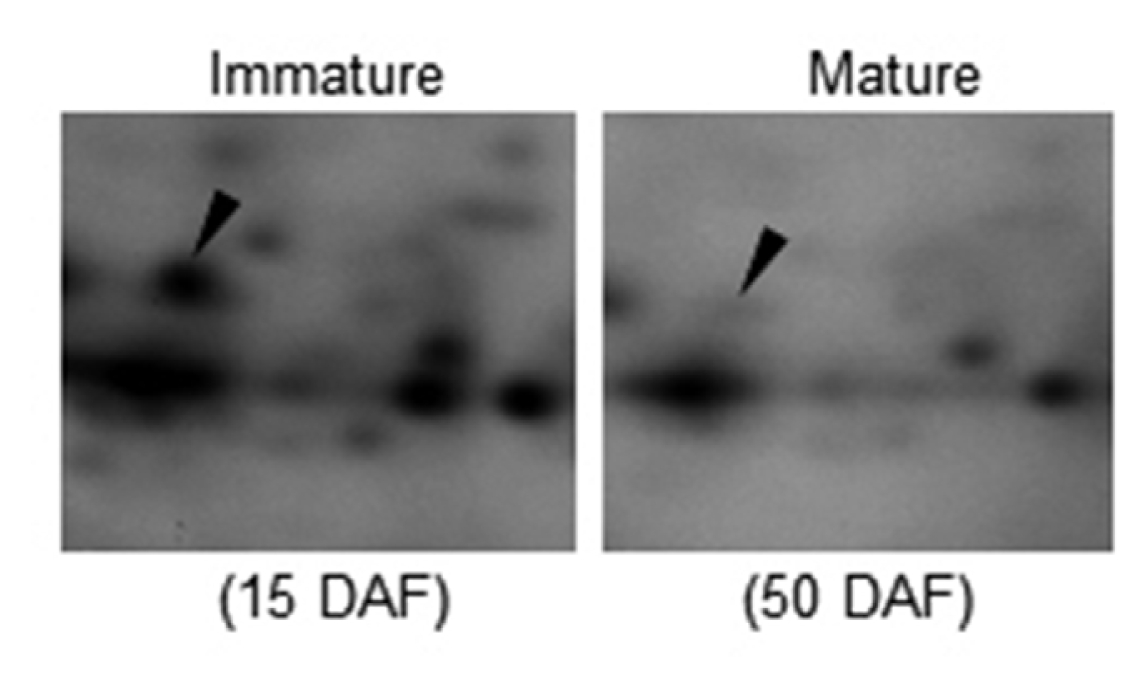
To understand the mechanism of seed development in rice, we tried to isolate proteins that are highly expressed during seed development. To this end, we employed a proteomic method. We first harvested the seeds from field-grown rice. Each seed was collected at 15 and 50 days after flowering (DAF). Total proteins were isolated from the collected seeds and then used for electrophoresis. As a result, we found that many kinds of proteins accumulated during seed development. We identified the protein spots using MALDI-TOF mass spectrometry. The level of OsAPx1 was significantly higher at 15 DAF than at 50 DAF (
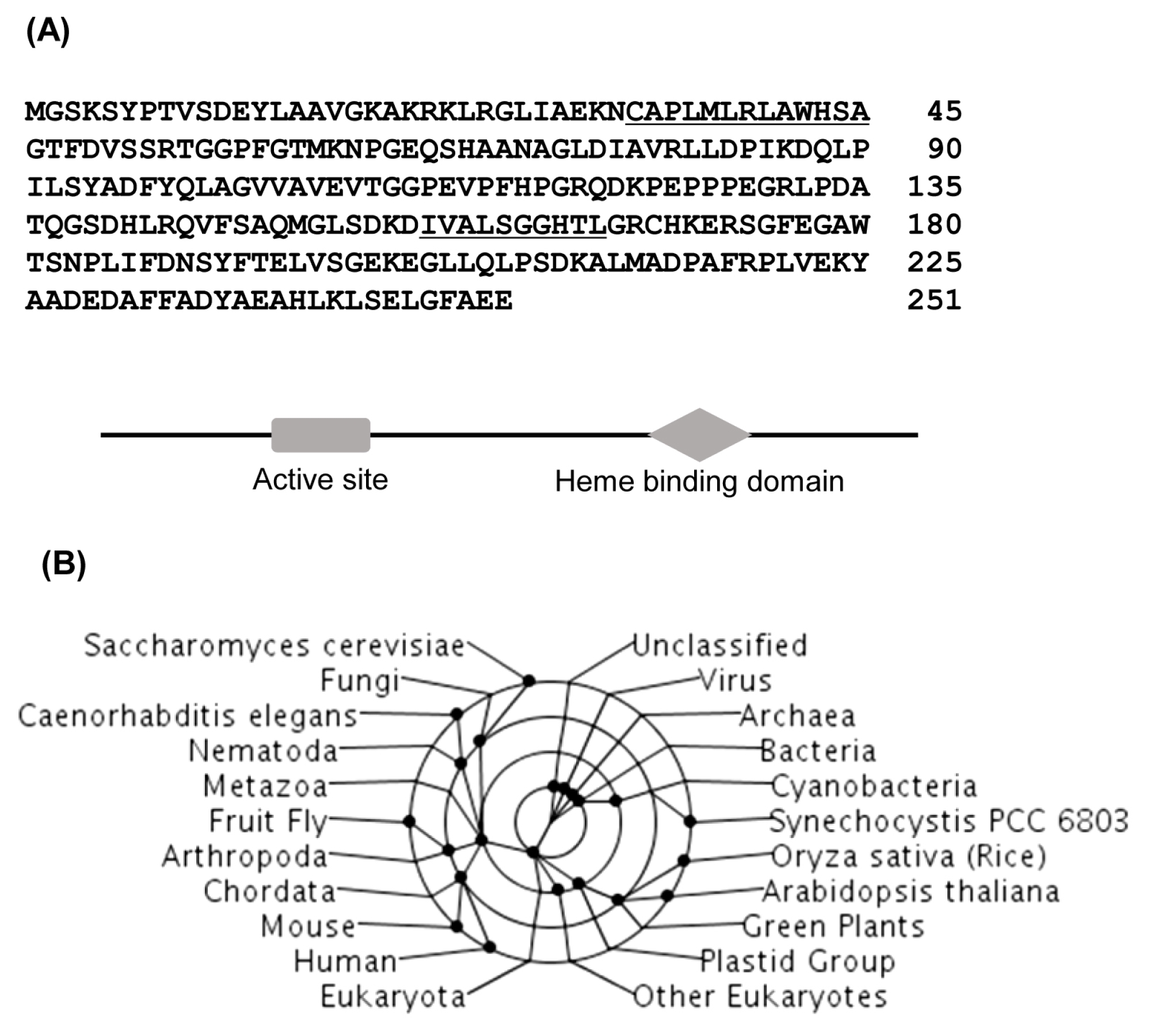
Fig. 1). We chose OsAPx1 for further experiments. It has been reported that APx has a key role in scavenging ROS and in protecting cells in various eukaryotes. An analysis of amino acid sequence of OsAPx1 showed it was a typical APx comprised of 251 amino acids with a heme binding domain (
Fig. 2A). Texanomic analysis revealed that OsAPx1 is very close to the APx of green plants including Arabidopsis (
Fig. 2B).
Next, we investigated the growth and phenotypes of the
Osapx1 mutants. For this experiment, we isolated a T-DNA insertion mutant of
OsAPx1 by performing PCR with
OsAPx1gene-specific and T-DNA-specific primers and examined its phenotype. The results showed that the growth of the
Osapx1mutant was similar to that of the wild type (
Fig. 3A). However, seed development of the
Osapx1 mutant was totally different from that of the wild type (
Fig. 3A and B,
Table 1). Weight per panicle was about 5.05 for the wild type and 2.01 for the
Osapx1mutant, although the number of total seeds was similar (147 in wild type and 139.67 in
Osapx1mutant). Over 58% of seeds were aborted in the
Osapx1 mutant whereas only 3% of seeds were aborted in the wild type.
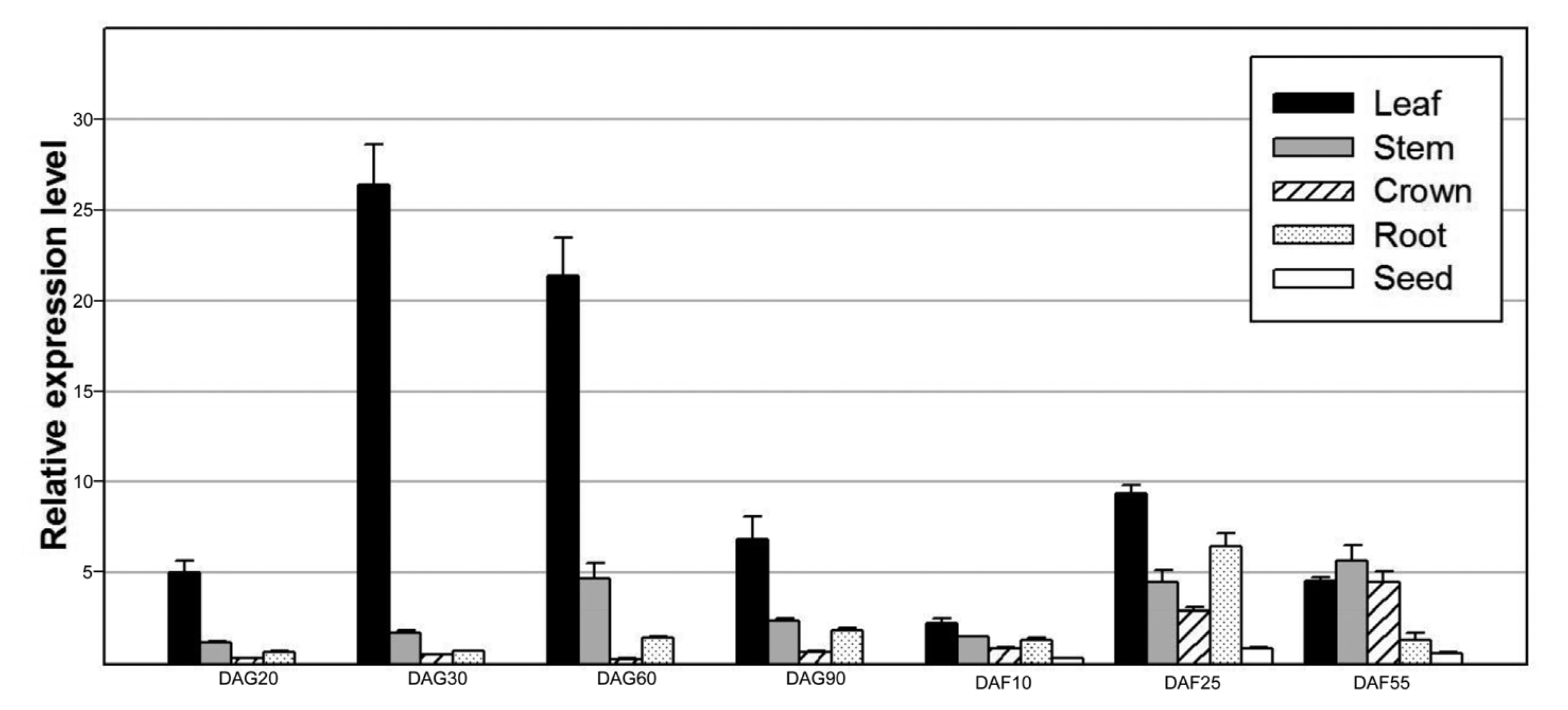
Because OsAPX1 accumulated to high levels in developing seed, we further examined its expression over the entire growth period. We collected samples from DAG 20 to DAF 55 and then used them for quantitative real time RT-PCR. As a result, we found that
OsAPX1 is highly expressed in leaf during development even if it is also expressed in stem, crown, root, and seed (
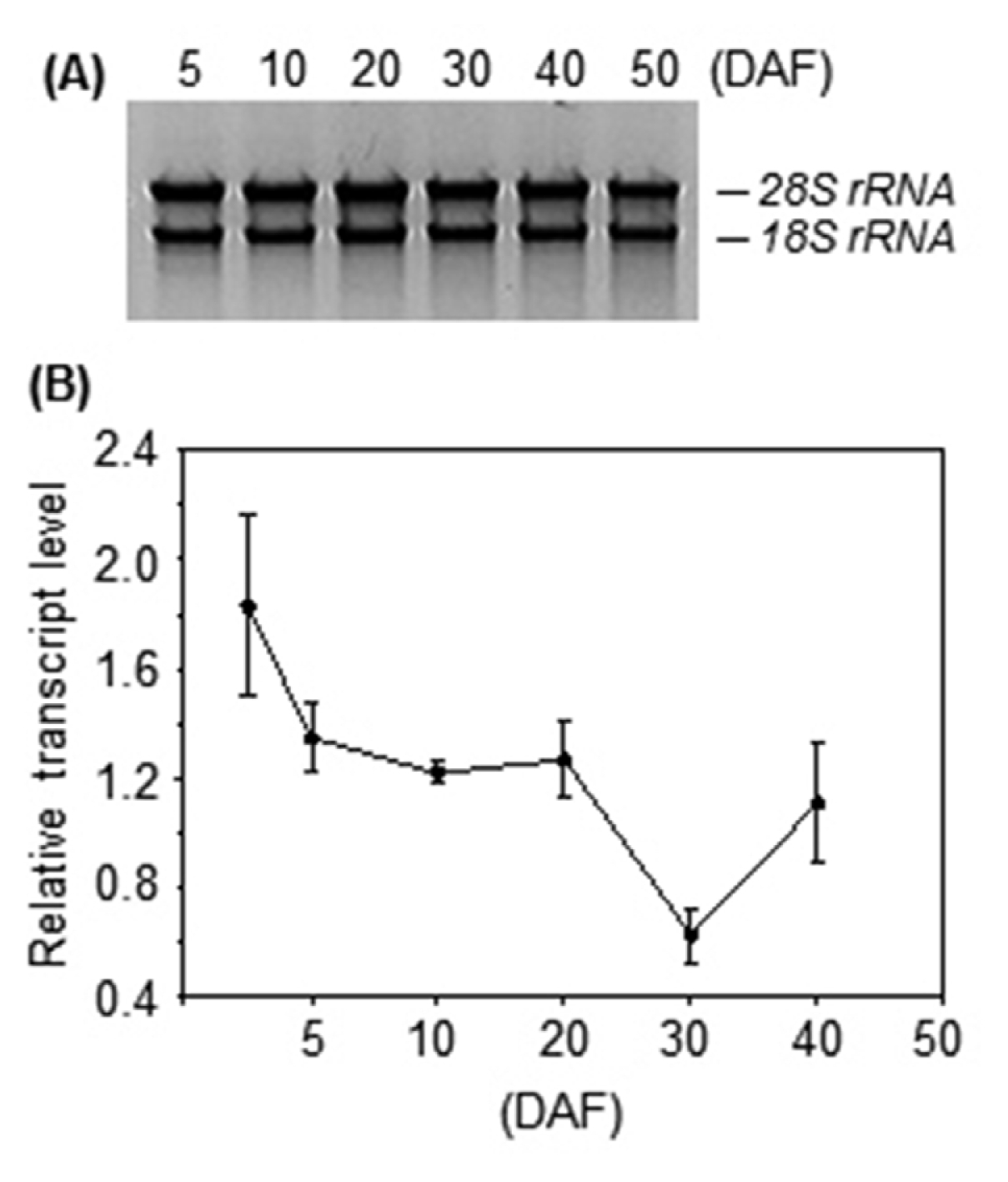
Fig. 4). We also examined its expression during seed development. The results showed that its transcript level gradually decreased after flowering (
Fig. 5). Proteomic analysis showed that the level of OsAPx1 was also much higher in immature seed than in mature seed (
Fig. 1). These results strongly suggest that OsAPx1 is not modified by either small or large molecules after translation, indicating that its function and stability is probably not modulated by post-translational modifications.
DISCUSSION
Here, we report that rice ascorbate peroxidase plays a pivotal role in seed development, although its primary function is a scavenger of ROS.
A recent study reported the identification of a multigene family encoding eight ascorbate peroxidases in rice: two cytosol (OsAPx1 and OsAPx2), two putative peroxisome (OsAPx3 and OsAPx4), and four putative chloroplast isoforms (OsAPx5 to OsAPx8), of which OSAPX8 is a putative thylakoid-bound isoform (Teixeira et al. 2004). Nevertheless, little is known about the function of rice APx, although its role has been well identified in various eukaryotes.
We isolated a T-DNA insertion mutant and characterized the growth and phenotype of the mutant. We expected its growth pattern and morphology to be different from those of the wild type because this mutant is known to accumulate detrimental levels of ROS. However, the growth and development pattern of
Osapx1 mutant was similar to that of the wild type before flowering (
Fig. 3A). For example, there were no differences in germination, growth of leaf and root, or tillering between the wild type and
Osapx1 mutant. We therefore focused on the possible role of OsAPx1 in seed set and development using the
Osapx1 mutant. To this end, we examined the size and number of panicles, number of seeds per panicle, and the abortion ratio in the
Osapx1 mutant. The panicle size of the
Osapx1 mutant was much shorter than that of the wild type (
Fig. 3), and the weight of each panicle was approximately 5.05 and 2.01 in the wild type and
Osapx1 mutant (
Table 1), respectively, suggesting that
Osapx1 controls panicle development. The number of seeds per panicle was 147 and 140 in the wild type and
Osapx1mutant, respectively (
Table 1). Finally, we examined seed abortion and found that approximately 58% of seeds of the
Osapx1 mutant were aborted (
Table 1). Taken together, the results strongly suggest that
Osapx1 plays an important role in seed set and rice maturation.
On the basis of the phenotype analyses, we speculated that its phenotype would correlate with the expression pattern of
Osapx1 gene during development. To check the relationship between the phenotypes of the
Osapx1 mutant and the expression of the
OsAPx1 gene, we examined the expression pattern of the
OsAPx1 gene in various rice organs during whole development by quantitative real-time RT-PCR. The results showed that the transcript level of
OsAPx1 was highest in young leaves during vegetative growth (
Fig. 4), indicating that OsAPx1 is required for leaf development. Notably, its transcript level in seeds was dramatically decreased during maturation (
Fig. 5). The protein level of OsAPx1 was also high in immature seeds, implying that the function and stability of OsAPx1 is not controlled by post-translational modifications and that OsAPx1 plays an important role in not only leaf development during the vegetative stage but also in seed development during the filling and ripening stages. These results indicate that the morphological features and abnormal seed yields of the OsAPx1 mutant strictly correlate with stage- and organ-specific expression of the
OsAPx1 gene.
It is well known that tillers are lateral organs that develop during the vegetative stage in rice. However, panicle branches can also act as lateral organs during the reproductive stages in rice (
Wang and Li 2011). In addition, in several cases, these two kinds of lateral organs are regulated by common mechanisms and change coordinately, although the number of tillers or panicle branches do not always undergo a similar change in most cases (
Wang and Li 2011). The OsAPx1 mutant showed reduced panicle size, panicle weight, and grain yield compared with the wild type, but had a similar number of tillers, indicating that the two organs are not coordinately regulated by OsAPx1 and that OsAPx1 activity is necessary for seed development.
In conclusion, the results of this study demonstrate that panicle development in the OsAPx1 mutant results in abnormal seed development, which in turn results in abnormal seeds and low grain yield. Further expression analysis of OsAPx1 using reporter genes, such as GUS and GFP, could provide helpful information about how OsAPx1 regulates panicle development. Moreover, identification of OsAPx1-regulating enzymes may provide crucial clues for understanding the role of OsAPx1 in the control of rice panicle and grain development
ACKNOWLEDGEMENTS
This work was supported by a grant from the Next-Generation BioGreen 21 Program (Plant Molecular Breeding Center no. PJ01108701), Rural Development Administration, Republic of Korea.
Fig. 1Relative intensity of ascorbate peroxidase 1 (OsAPx1) in immature and mature seeds. Total seed proteins were extracted from the wild type and Osapx1 mutant and then analyzed by two dimensional gel electrophoresis. Protein spot 3106 was identified as an OsAPx1 by peptide mapping. The OsAPx1 level is 11.28 folds higher at 15 DAF (Day after flowering, immature seed) than at 50 DAF (mature seed). Arrow heads indicate the protein spots of OsAPx1.
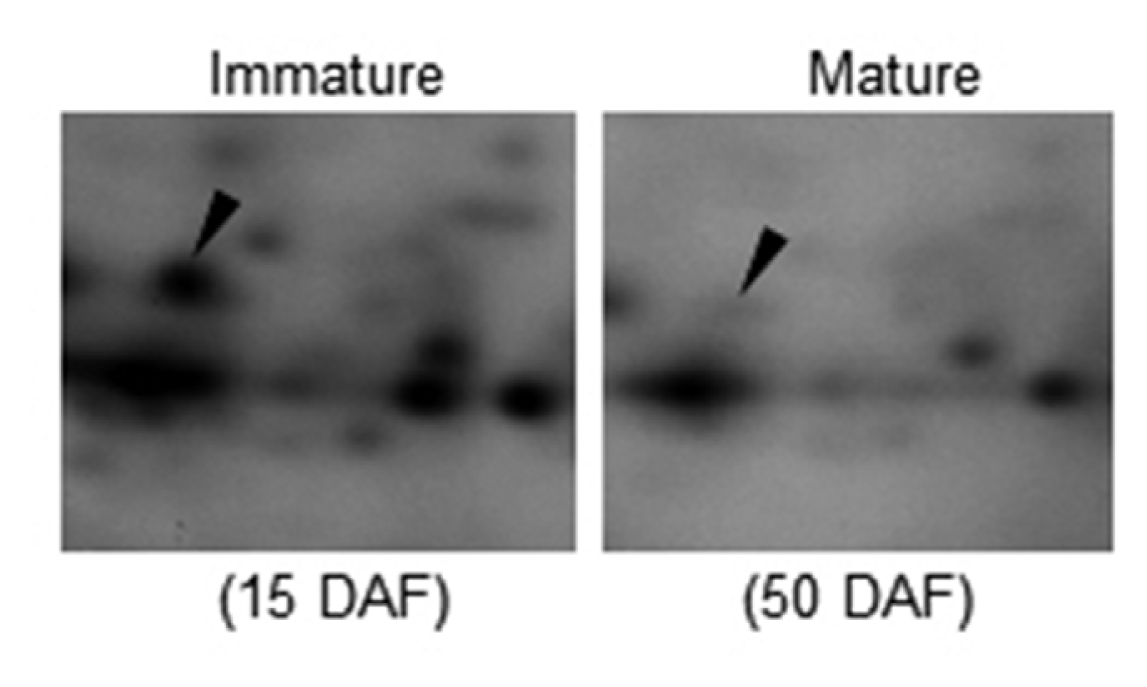
Fig. 2Deduced amino acid sequences of OsAPx1 (Accession number: AB050724). (A) Active site (34th to 45th aa) and heme binding motif (157th to 166th aa) are underlined (upper panel). The positions of the active site and the heme binding domain are also shown as a schematic diagram (lower panel). (B) Taxonomic lineage of rice ascorbate peroxidase (OsAPx1) amino acid sequences with those of its homologs in various species. The sequences of the proteins were aligned using the CLUSTALW2 software program and a taxonomic lineage was generated using the MEGA5 software program.
Fig. 3Phenotypes of the Osapx1 mutant. Wild-type and Osapx1 mutant rice were grown in the field and photographed (A, left). Panicles were collected from wild type and the Osapx1 mutant and then photographed (A, right). The weight of the panicle in the Osapx1 mutant is significantly low in the mutant, leading to abnormal seeds. (B) The seeds were collected from the wild type and the Osapx1 mutant. The hulls of the seeds were opened and then photographed.

Fig. 4Expression profile of OsAPx1 during development. Total RNA was isolated from various tissues of field-grown wild-type plants at the indicated stages and the transcript level of OsAPx1 was analyzed by quantitative real-time RT-PCR using gene-specific primers. OsAPx1 expression was highest in young leaves during vegetative development, although it was also expressed in other organs, including seeds. DAG, day after germination; DAF, day after flowering.
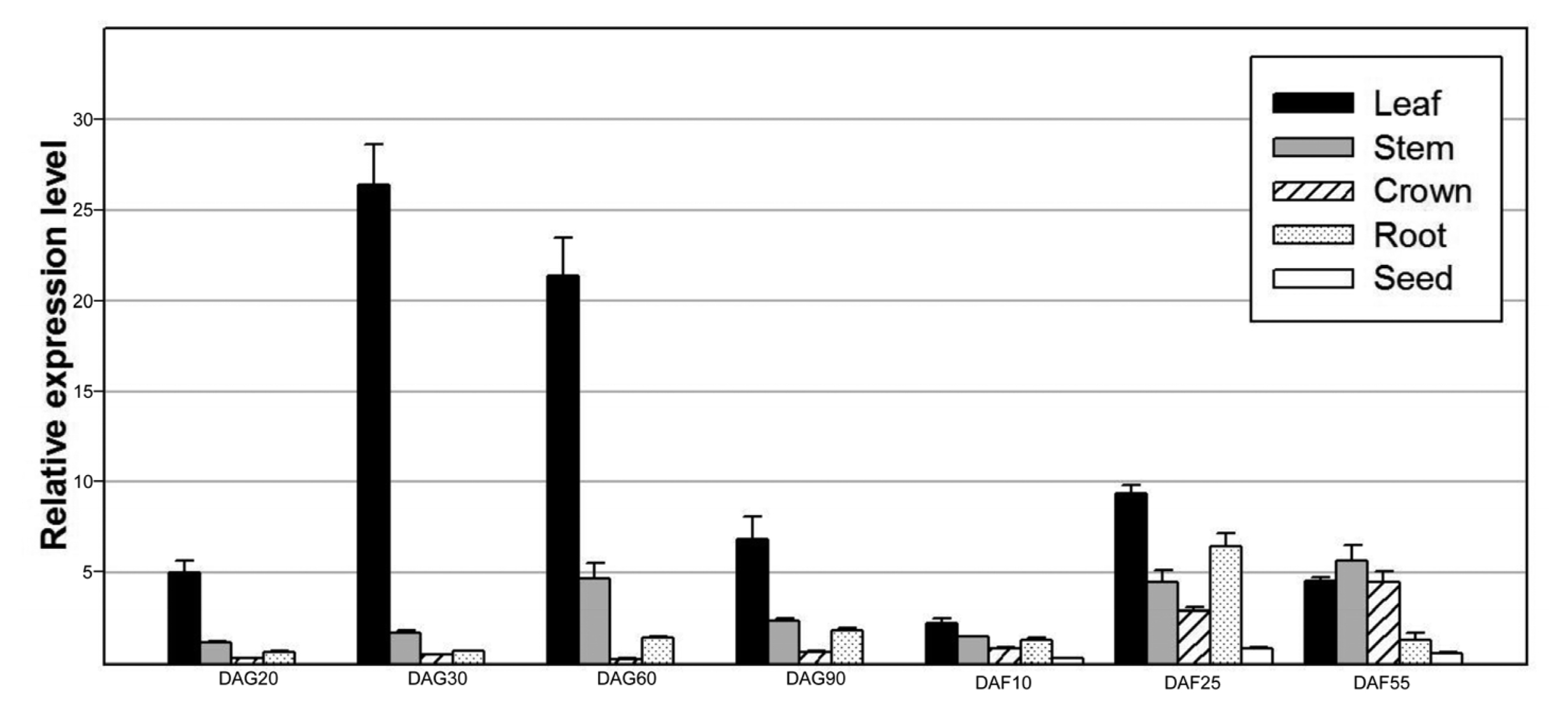
Fig. 5Transcript profile of OsAPx1 during seed development. (A) Total RNA was isolated from harvested seeds at the indicated time-points and then separated on a formaldehyde–agarose gel. (B) First-strand cDNA was synthesized from equal quantities of total RNA and then analyzed by quantitative real-time PCR using the gene-specific primers. The amplified cDNA from the total RNA sample extracted at 5 DAF was used as a standard to compare relative quantities.
Table 1Comparison of panicle weight, and number of normal and aborted seeds between wild-type and Osapx1 mutant.
Table 1
|
Weight of per panicle |
No. of total seed per panicle |
No. of aborted seed per panicle (%) |
|
WT |
5.05 ± 0.67 |
147 ± 23.73 |
4 ± 2.98 (2.7) |
|
Osapx1
|
4.51 ± 0.44 |
139 ± 21.43 |
81 ± 13.10 (58.3) |
References
- Agrawal GK, Jwa N, Iwahashi H, Rakwal R. 2003. Importance of ascorbate peroxidases OsAPx1 and OsAPx2 in the rice pathogen response pathway and growth and reproduction revealed by their transcriptional profiling. Gene. 322: 93-103.
- Altenbach SB, Howell SH. 1981. Identification of a satellite RNA associated with turnip crinkle virus. Virology. 112: 25-33.
- Bai X, Luo L, Yan W, Kovi Mallikarjuna Rao, Zhan W, Xing Y. 2010. Genetic dissection of rice grain shape using a recombinant inbred line population derived from two contrasting parents and fine mapping a pleiotropic quantitative trait locus qGL7. BMC Genet. 11: 1-11.
- Dat J, Vandenabeele S, Vranová E, Van Montagu M, Inzé D, Van Breusegem F. 2000. Dual action of the active oxygen species during plant stress response. Cell Mol Life Sci. 57: 779-795.
- Fan C, Xing Y, Mao H, Lu T, Han B, Xu C, Li X, Zhang Q. 2006. GS3, a major QTL for grain length and weight and minor QTL for grain width and thickness in rice, encodes a putative transmembrane protein. Theor Appl Genet. 112: 1164-1171.
- Halliwell B, Gutteridge JM. 1999. Free radicals in biology and medicine. Oxford University Press. New York: pp. 936.
- Kapazoglou A, Tondelli A, Papaefthimiou D, Ampatzidou H, Francia E, Stanca MA, Bladenopoulos K, Tsaftaris AS. 2010. Epigenetic chromatin modifiers in barley: IV. The study of barley polycomb group (PcG) genes during seed development and in response to external ABA. BMC Plant Biol. 10: 1-19.
- Kesavan M, Song JT, Seo HS. 2013. Seed size: a priority trait in cereal crops. Physiol Plantarum. 147: 113-120.
- Khush G. 2003. Productivity improvement in rice. Nutr Rev. 61: 114-116.
- Kohler C, Page DR, Gagliardini V, Grossniklaus U. 2005. The Arabidopsis thaliana MEDEA Polycomb group protein controls expression of PHERES1 by parental imprinting. Nat Genet. 37: 28-30.
- Liu Y, Whittier R. 1995. Thermal asymmetric interlaced PCR: automatable amplification and sequencing of insert end fragments from P1 and YAC clones for chromosome walking. Genomics. 25: 674-681.
- Menezes-Benavente L, Kernodle SP, Margis-Pinheiro M, Scandalios JG. 2004a. Salt-induced antioxidant metabolism defenses in maize (Zea mays L.) seedlings. Redox Rep. 9: 29-36.
- Menezes-Benavente L, Teixeira FK, Kamei CL, Margis-Pinheiro M. 2004b. Salt stress induces expression of genes encoding antioxidant enzymes in seedlings of a Brazilian indica rice (Oryza sativa L.). Plant Sci. 166: 323-331.
- Mittler R, Feng X, Cohen M. 1998. Post-transcriptional suppression of cytosolic ascorbate peroxidase expression during pathogen-induced programmed cell death in tobacco. Plant Cell. 10: 461-473.
- Mittler R, Vanderauwera S, Gallery M, Van Breusegem F. 2004. Reactive oxygen gene network of plants. Trends Plant Sci. 9: 490-498.
- Mullineaux P, Karpinski S. 2002. Signal transduction in response to excess light: getting out of the chloroplast. Curr Opin Plant Biol. 5: 43-48.
- Oakley BR, Kirsch DR, Morris NR. 1980. A simplified ultrasensitive silver stain for detecting proteins in polyacrylamide gels. Anal Biochem. 105: 361-363.
- Orozco-Cárdenas ML, Narváez-Vasquéz J, Ryan CA. 2001. Hydrogen peroxide acts as a second messenger for the induction of defense genes in tomato plants in response to wounding, systemin, and methyl-jasmonate. Plant Cell. 13: 179-191.
- Peng S, Huang J, Sheehy John E, Laza Rebecca C, Visperas Romeo M, Zhong X, Centeno Grace S, Khush Gurdev S, Cassman Kenneth G. 2004. Rice yields decline with higher night temperature from global warming. Proc Natl Acad Sci USA. 101: 9971-9975.
- Sato Y, Murakami T, Funatsuki H, Matsuba S, Saruyama H, Tanida M. 2001. Heat shock-mediated APX gene expression and protection against chilling injury in rice seedlings. J Exp Bot. 52: 145-151.
- Scandalios JG. 2002. The rise of ROS. Trends Biochem Sci. 27: 483-486.
- Schruff MC, Spielman M, Tiwari S, Adams S, Fenby N, Scott RJ. 2005. The AUXIN RESPONSE FACTOR 2 gene of Arabidopsis links auxin signalling, cell division, and the size of seeds and other organs. Development. 133: 251-261.
- Shevchenko A, Wilm M, Vorm O, Mann M. 1996. Mass spectrometric sequencing of proteins from silver-stained polyacrylamide gels. Anal Chem. 68: 850-858.
- Shomura A, Izawa T, Ebana K, Ebitani T, Kanegae H, Konishi S, Yano M. 2008. Deletion in a gene associated with grain size increased yields during rice domestication. Nat Genet. 40: 1023-1028.
- Shigeoka S, Nakano Y, Kitaoka S. 1980. Metabolism of hydrogen peroxide in Euglena gracilis Z by L-ascorbic acid peroxidase. Biochem J. 186: 377-380.
- Shigeoka S, Ishikawa T, Tamoi M, Miyagawa Y, Takeda T, Yabuta Y, Yoshimura K. 2002. Regulation and function of ascorbate peroxidase isoenzymes. J Exp Bot. 53: 1305-1319.
- Song XJ, Huang W, Shi M, Zhu MZ, Lin HX. 2007. QTL for rice grain width and weight encodes a previously unknown RING-type E3 ubiquitin ligase. Nat Genet. 39: 623-630.
- Tamura K, Peterson D, Peterson N, Stecher G, Nei M, et al. 2011. MEGA5: Molecular evolutionary genetics analysis using Maximum Likelihood, Evolutionary Distance, and Maximum Parsimony methods. Mol Biol Evol. 28: 2731-2739.
- Tanabe S, Ashikari M, Fujioka S, Takatsuto S, Yoshida S, Yano M. 2005. A novel cytochrome P450 is implicated in brassinosteroid biosynthesis via the characterization of a rice dwarf mutant (dwarf11) with reduced seed length. Plant Cell. 17: 776-790.
- Vandenabeele S, Van Der Kelen K, Dat J, Gadjev I, Boonefaes T, Morsa S, Rottiers P, Slooten L, Van Montagu M, Zabeau M, Inzé D, Van Breusegem F. 2003. A comprehensive analysis of hydrogen peroxide-induced gene expression in tobacco. Proc Natl Acad Sci USA. 100: 16113-16118.
- Wang Y, Li J. 2011. Branching in rice. Curr Opin Plant Biol. 14: 94-99.
- Wang E, Wang J, Zhu X, Hao W, Wang L. 2008. Control of rice grain-filling and yield by a gene with a potential signature of domestication. Nat Genet. 40: 1370-1374.
- Weng JF, Gu SH, Wan XY, Gao H, Guo T, Su N, Lei CL, Zhang X, Cheng ZJ, Guo XP, Wang JL, Jiang L, Zhai HQ, Wan JM. 2008. Isolation and initial characterization of GW5, a major QTL associated with rice grain width and weight. Cell Res. 18: 1199-1209.
- Xiao W, Brown Roy C, Lemmon Betty E, Harada John J, Goldberg Robert B, Fischer Robert L. 2006. Regulation of seed size by hypomethylation of maternal and paternal genomes. Plant Physiol. 142: 1160-1168.
- Yang J, Zhang J. 2006. Grain filling of cereals under soil drying. New Phytol. 169: 223-236.
- Yoshimura K, Yabuta Y, Ishikawa T, Shigeoka S. 2000. Expression of spinach ascorbate peroxidase isoenzymes in response to oxidative stresses. Plant Physiol. 123: 223-233.
- Zhang H, Wang J, Nickel U, Allen RD, Goodman HM. 1997. Cloning and expression of an Arabidopsis gene encoding a putative peroxisomal ascorbate peroxidase. Plant Mol Biol. 34: 967-971.