Abstract
Most Bt transgenic rice events have an underlying grain cost depending on the insect pressure in the field. This study was conducted (1) to determine if the cry1Ac/bar genetically modified (GM) rice (designated Agb0101) based on a Korean japonica rice cultivar has any underlying grain cost and (2) to develop cry1Ac/bar GM rice lines with yield performance similar to those of Korean elite cultivars via crossbreeding. A three-year field experiment showed that Agb0101 had a high underlying grain cost equivalent to 18% grain yield loss, and this cost was mainly due to the grain number per panicle. Moreover, it was found that the cry1Ac/bar transgene was inherited as a single dominant gene. BC1F4―F5 crossbred progeny derived from an anther culture of Agb0101―showed phenotypic resemblance to non-transgenic Korean elite cultivars, resulting in a decrease in the grain yield cost of Agb0101 from 27% to 10%. These results suggest that Bt-GM rice-oriented cross-fertilization with a non-GM elite cultivar followed by two- or threefold selection is a feasible method to minimize or remove the underlying grain cost of the Bt transgenic insect-resistant rice line.
-
Key words: cry1Ac, Bt transgenic rice, Underlying grain cost, Crossbreeding
INTRODUCTION
More than half of the world’s six billion people eat rice as their staple food, particularly in Asia where stable rice production is of national interest to countries (
Garris et al. 2005;
Lu and Snow 2005;
Yu et al. 2002). Asian countries such as Korea, China, and Japan have been investing in rice research to develop high-yielding rice varieties resistant to pests and diseases in order to stabilize and increase rice production. However, these pest-resistant rice varieties have been insufficient to protect rice plants from pests. Thus, pesticides have been widely applied to control pests in rice fields. In addition, recently developed pest-resistant transgenic rice has attracted considerable attention as its use can increase plant resistance to insects and reduce the need for pesticides (
Kim et al. 2009).
Many countries such as China, India, Vietnam, and Iran have invested in the development of
Bt transgenic rice; numerous
Bt rice lines have been produced and are in the pipeline for commercialization (
Lu and Snow 2005;
Wang and Johnston 2007). Although many
Bt transgenic rice lines have been developed and evaluated (
Fujimoto et al. 1993;
High et al. 2004;
Jiang et al. 2013;
Kim et al. 2008;
Shu et al. 2000;
Tu et al. 2000;
Wang et al. 2002;
Wang et al. 2010;
Wang et al. 2012a,
2012b;
Wu et al. 2002;
Xia et al. 2011;
Yea et al. 2001), only three insect-resistant
Bt transgenic rice lines carrying
cyr1Ab and/or
cry1Ac have been approved in China and Iran (ISAAA’s GM Approval Database:
http://www.isaaa.org/gmapprovaldatabase/, as of January 16, 2014). Among the crystal proteins (
Bt proteins) from
Bacillus thuringiensis that produce δ-endotoxin,
cry1Ab,
cry1Ac, and their synthetic proteins are highly specific to lepidopteran insects such as
Chilo suppressalis (striped stem borer),
Scirpophaga incertulas (yellow stem borer), and
Cnaphalocrocis medinalis (leaf folder), which cause significant rice yield losses annually (
Kim et al. 2009;
Lee et al. 2006;
Nathan 2006;
Shu et al. 2000). In addition, almost half of the insecticides used in the rice fields in Asia are targeted at lepidopteran insect species (
Heong et al. 1994). Transgenic
Bt rice allows farmers to reduce pesticide use by nearly 80% compared to that in the case of conventional rice varieties (
Huang et al. 2005).
Several conflicting reports are available on the yield performance of
Bt transgenic rice in the field; results presented in these reports vary depending on the transgenic line and insect infestation conditions (
Huang et al. 2005;
Kim et al. 2008;
Wang et al. 2012a,
2012b;
Xia et al. 2010,
2011). However, all of these reports indicate that the underlying cost of the grain yield of
Bt transgene(s) may be usually incurred in the field. Therefore, well-designed transgenic breeding measures are necessary to minimize the potential cost of
Bt transgene(s) before commercial production of genetically modified (GM) rice varieties can be realized.
In Korea, the Rural Development Administration (RDA) created three
Bt transgenic rice lines carrying a synthetic truncated
cry1Ac gene harboring a bar gene, and selected a homozygous event called C7-1 as the breeding material. C7-1 was selected as it has a single intact copy of T-DNA targeted to the chloroplast (
Lee et al. 2009) and high resistance to insects such as the rice leaf folder, rice green caterpillar, and rice skipper under laboratory conditions, and to the rice leaf folder under field conditions (
Kim et al. 2009).
The present study was conducted to (1) determine if cry1Ac/bar transgenic rice Agb0101 has any underlying grain cost, (2) analyze the genetic action of transgenes transferred from Agb0101 to non-GM elite rice cultivars via crossing, and (3) develop Agb0101 crossbred lines with yield performance similar to that of Korean elite cultivars by using crossbreeding measures.
MATERIALS AND METHODS
Field growth conditions for and phenotypic evaluation of Agb0101
The synthetic truncated
cry1Ac transgenic T
3 line (Agb0101), derived from the C7-1 transgenic rice event developed by the RDA, South Korea (
Lee et al. 2009), and its non-transgenic counterpart cv. Nakdongbyeo, a
japonica type, were used in three years of field experiments from 2006 to 2008 to evaluate their field performance. These experiments were conducted in the LMO paddy field of the National Institute of Crop Science (35°56′N, 126°55′E; Iksan, South Korea). Treatments were arranged in a randomized complete block design with three replications and a plot size of 9.45 m
2. Thirty-day seedlings were transplanted in a transplanting space of 30 cm × 15 cm with three seedlings per hill. The stem length (cm), panicle length (cm), and panicle number per hill (no. hill
−1) were measured from 10 hills in each plot. The grain number per panicle (no. panicle
−1) was counted from the 10 sampled hills after hand-threshing; the numbers of filled and unfilled grains were counted after separating them by submerging three 30-g grain subsamples from each plot in tap water. The filled grain percentage (%) was calculated by dividing the number of filled grains by the total grain number for each subsample. The 1000-grain weight (g) based on brown rice and grain yield (kg 10a
−1) based on white rice were determined for each plot and adjusted to a moisture content of 15% on a dry basis.
Data are presented as mean ± standard error or standard deviation calculated by the SAS 9.2 PROC MEANS procedure (SAS Institute, Cary, NC, USA). The recorded data were then analyzed by analysis of variance (ANOVA), and differences between means of the lines were tested by employing the least significant difference (LSD) test at the 0.05 probability level by using the PROC ANOVA procedure.
Genetic analysis of crossbred progenies between Agb0101 and Korean elite cultivars
Agb0101 and cv. Dongjinbyeo (a Korean elite
japonica cultivar) were crossed, and their F
1 hybrid seeds were harvested. Anthers were cultured from these F
1 hybrid plants as described by
Won et al. (2004). Fifteen anther derived green plants were obtained after chromosome doubling performed using colchicine; these doubled-haploid (DH) plants were then planted in a greenhouse. At the three-leaf stage, a 0.3% (v/v) glufosinate ammonium solution (Basta®; Bayer CropScience Korea, Seoul, South Korea) was sprayed evenly on the plants, and the surviving six
bar-resistant plants were transplanted at one seedling per hill. Twenty seeds of each of the six DH lines were planted; all the plants were resistant to Basta®. Genomic DNA was extracted from mature leaves of three plants from each of the six DH lines by using the CTAB extraction method to examine the T-DNA insertion site harboring the
cyr1Ac gene (
Chen and Ronald 1999). The analysis was performed as described by
Lee et al. (2009) and was followed by NCBI nucleotide blast search.
In total, 1,111 plants from 9 F2 populations derived from the cross between Agb0101 and cv. Nakdongbyeo were used for an inheritance analysis of the cry1Ac/bar transgene. To analyze its inheritance in the backcross population, 321 plants from each of the seven BC1F1 populations derived from crosses between Agb0101 and seven non-GM Korean cultivars, namely, cv. Odaebyeo, cv. Taeseongbyeo, cv. Gounbyeo, cv. Hwaseongbyeo, cv. Ilpumbyeo, cv. Hopyeongbyeo, and cv. Gopumbyeo, were used. Basta® solution was sprayed evenly on the plants at the three-leaf stage, and the Basta® resistance of the plants was evaluated 14 days after spraying to investigate the segregation ratio of the cry1Ac/bar transgenes in the F2 and BC1F1 populations. Plants with no damage were classified as Basta®-resistant, whereas damaged ones were classified as Basta®-susceptible. A segregation analysis was performed using the chi-square test according to Mendelian segregation hypotheses. The chi-square test was performed using R package ver. 3.0.1 (R software, Vienna, Austria).
Development of crossbred cry1Ac/bar transgenic lines with field performance similar to that of Korean elite cultivars
In 2009, 41 cry1Ac/bar transgenic breeding lines derived from 17 cross-combinations of Agb0101 and 10 Korean elite japonica cultivars provided by the National Institute of Crop Science were used to select elite Bt transgenic lines that showed the best field performance. Among the 41 crossbred lines, 13 were F3 progeny of the DHs originating from the anther culture of F1 hybrids from 5 cross-combinations; another other 13 were self-pollinated F4 progeny from 7 cross-combinations. The rest 15 were self-pollinated BC1F3 progeny from F1 hybrids backcrossed with their non-transgenic elite parent.
In 2010, 57 lines were selected from the 2009 entry according to the pedigree breeding method. Thirty seeds of each pedigree were planted, and Basta® solution was sprayed evenly on the plants at the three-leaf stage. Expression of the cry1Ac and bar genes in the leaves was investigated using the ImmunoStrip® assay kit (Agdia, Elkhart, IN, USA) and Trait LL test strips (Strategic Diagnostics Inc, Newark, DE, USA) in accordance with the manufacturers’ instructions. Twenty seedlings of each pedigree resistant to Basta® were transplanted. Field experiments were conducted at the LMO paddy field of the National Academy of Agricultural Sciences (37°15′N, 126°59′E; Suwon, South Korea). Three plants from each line showing the best field performance in each plot were selected; seven traits of these plants related to growth and yield (i.e., stem length, panicle length, panicle number per plant, grain number per panicle, filled grain percentage, 1000-grain weight, and grain yield per plant) were measured.
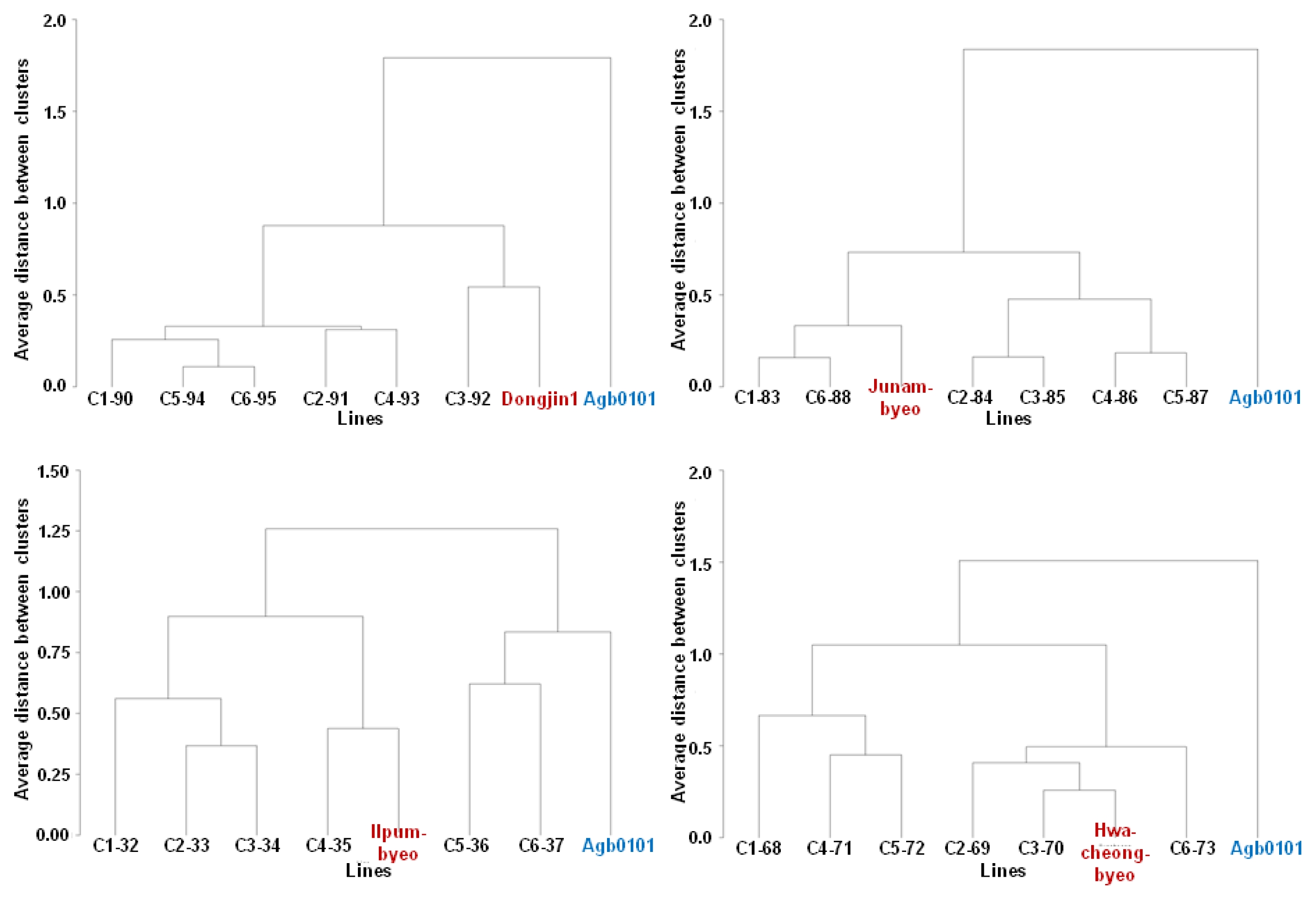
Data are presented as mean ± standard error or standard deviation calculated by the PROC MEANS procedure (SAS 9.2). Cluster analysis was then performed on the basis of the aforementioned seven traits to identify breeding lines that were similar to each non-transgenic elite cultivar parent. The crossbred lines and their parents were grouped by cluster analysis based on the average linkage method carried out using the PROC cluster procedure (SAS 9.2).
Fertilization method and pest management in experimental fields
Phosphorus (fused superphosphate) was applied at 4.5 kg·P2O5 10a−1 as the base. Potassium (potassium chloride) at 5.7 kg·K2O 10a−1 was applied with a split of 70% at the base and 30% at the panicle initiation stage. Nitrogen (urea) at 9.0 kg·N 10a−1 was applied as follows: 40% at base, 30% at mid-tillering, and 30% at panicle initiation. Pests (including Bt target pests), diseases, and weeds were intensively controlled to avoid yield loss.
RESULTS
Field performance of cry1Ac/bar transgenic rice Agb0101
Agb0101 had a significantly smaller stem length, panicle length, and yield components than those of its non-transgenic counterpart, cv. Nakdongbyeo. However, their 1,000-grain weights were very similar. Further, the panicle length and grain number per panicle of Agb0101 were ~20% less than those of cv. Nakdongbyeo. However, the trend observed for the panicle number per plant was opposite to those observed for other traits: Agb0101 had ~5% more panicles than cv. Nakdongbyeo, although the difference was not significant (
Table 1). This field performance eventually resulted in a significant grain yield difference of 79.3 kg 10a
−1 between Agb0101 and cv. Nakdongbyeo: the grain yield of Agb0101 was approximately 18% less than that of cv. Nakdongbyeo. This result suggests that the incorporation of
cry1Ac/bar in rice may cause a yield reduction in the field (
Table 1).
Considering the abovementioned results, it can be concluded that the underlying cost of grain yield was mainly due to the grain number per panicle in Agb0101. Therefore, minimizing the underlying cost of grain yield is necessary via breeding measures.
Genetic stability of cry1Ac/bar transgene in crossbred populations between Agb0101 and Korean elite cultivars
The chromosomal location of the cry1Ac/bar transgene was determined in the six DH lines derived from the anther culture of F1 hybrids (Agb0101 and cv. Dongjin1). All six DH lines had the flanking genomic T-DNA sequence carrying the cry1Ac/bar transgene at the same site on chromosome 1 as in Agb0101. This observation indicated that the transgene of Agb0101 can be stably delivered to other Korean elite cultivars from a chromosome-doubled anther culture via artificial crossing.
The inheritance of the Agb0101-originated
cry1Ac/bar transgene was analyzed on the basis of the herbicide response to Basta® of nine F
2 populations derived from crossing between Agb0101 and cv. Nakdongbyeo. Among the 1,111 F
2 plants evaluated, 823 were resistant and 288 were susceptible to Basta®, suggesting that the transgene in the F
2 progeny segregated at a resistant:susceptible ratio of 3:1, which is suitable for the single gene model (
Table 2). This result shows that the transgene was stably inherited as a single dominant gene.
Further, the inheritance of the transgene in the backcross populations was analyzed using seven BC
1F
1 populations derived from crossing between Agb0101 and seven Korean elite cultivars. Among the 321 BC
1F
1 plants evaluated, 153 F
2 plants were resistant and 168 were susceptible to Basta®, suggesting that the transgene in the BC
1F
1 progeny segregated at a resistant:susceptible ratio of 1:1, with a good fit to the single gene model (
Table 2). This result too supports the theory of the inheritance of the transgene as a single dominant gene in the backcross populations.
Twenty-three of 41 pedigrees planted in 2009 and 42 of 57 pedigrees planted in 2010 were resistant to Basta®, indicating that all these pedigrees had the homogeneous
cry1Ac/bar transgene. In particular, all the pedigrees derived from the DHs were resistant to Basta®, suggesting that the anther culture of the F
1 hybrid is a quick method of fixing recombinants of the
cry1Ac/bar transgenes (
Table 3). All plants of every pedigree resistant to Basta® showed positive reactions on each immunostrip assay for both
cry1Ac and
bar proteins (data not shown), confirming that the
cry1Ac/bar transgenes were stably inherited and expressed in the crossbred progeny. Of these, three plants showing the best yield performance were selected from each crossbred line by considering the resemblance to their non-transgenic elite cultivar obtained via cluster analysis by using seven agronomic traits (including grain yield and homogeneity) of
cry1Ac/bar transgenes based on Basta® resistance. As a result, 57 plants in 2009 and 69 in 2010 were selected and advanced to the next generation (
Table 3).
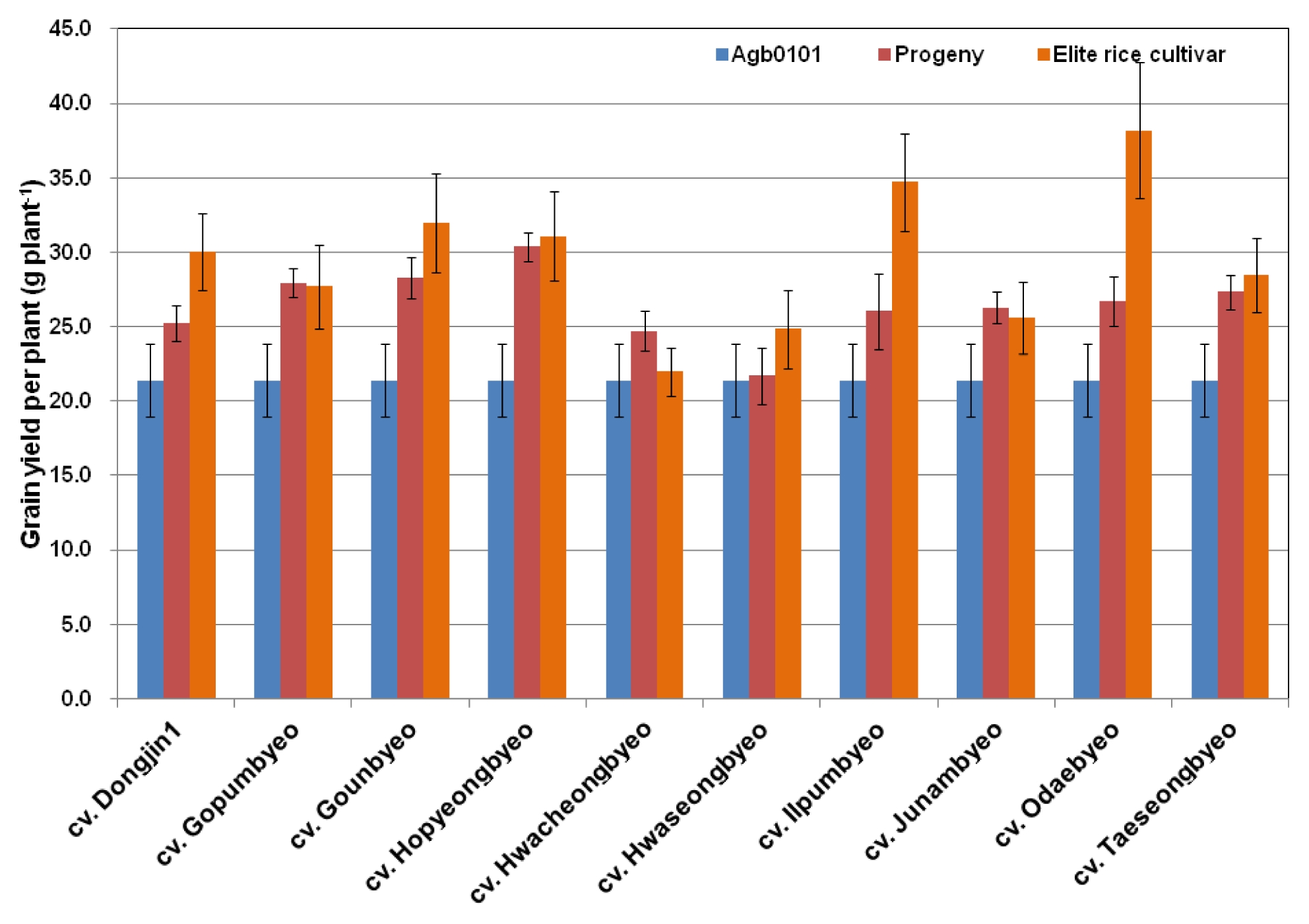
For the 57 lines planted in 2010, the stem length, panicle length, and grain yield components such as the filled grain percentage, panicle number plant, grain number per panicle, and 1,000-grain weight of Agb0101, the 10 elite cultivars, and all of their crossbred progeny were compared. The stem length, filled grain percentage, and 1,000-grain weight of all crossbred progeny were not significantly different from those of their parents. The Agb0101 plants had ~15% shorter panicle length, ~31% fewer grain numbers per panicle, and ~18% more panicles than those of the 10 elite cultivars. All crossbred progeny lines had a 2–3% difference in the panicle length, grain number per panicle, and panicle number per plant from their elite cultivar parents (
Fig. 1). These results show that the principal traits of the crossbred progeny lines related to their growth and yield changed from those of Agb0101 to those of their elite cultivar parents. Moreover, Agb0101 had approximately 27% lower grain yield, but its 57 crossbred progeny lines showed approximately 10% lower grain yield than their elite cultivar parents (
Fig. 1).
The crossbred progeny lines were grouped by cluster analysis based on the stem length, panicle length, yield, and yield components such as filled grain percentage, panicle number per plant, grain number per panicle, and 1,000-grain weight. After the cluster analysis, all the crossbred progeny lines were grouped with their elite cultivar parents (
Fig. 2) except for those derived from crossing between Agb0101 and cv. Hwaseongbyeo (data not shown), cv. Ilpumbyeo (
Fig. 2), and cv. Odaebyeo (data not shown).
Furthermore, the grain yield of the crossbred progeny lines was compared with that of their parents, Agb0101, and each of the 10 elite cultivars. The progeny lines showed grain yields similar to those of their elite cultivar parents except for those derived from crossing with cv. Dongjin1, cv. Ilpumbyeo, and cv. Odaebyeo (
Fig. 3), which had plants with a greater yield than their elite cultivar parent. This observation suggests that further selection will improve the grain yield of the crossbred progeny lines.
DISCUSSION
In this study, a three-year field experiment showed that Agb0101 had a high underlying grain cost equivalent to 18% grain yield loss; this cost was mainly due to the grain number per panicle. Therefore, efforts for minimizing the underlying cost of grain yield are necessary via breeding measures.
Xia et al. (2010,
2011) suggested that effective biotechnology and breeding measures are needed to minimize the potential underlying cost of an insect-resistant transgene(s) before commercial production of GM crops can be realized. Other studies on F
1 hybrid rice lines showed that the
Bt transgenic F
1 hybrid obtained by crossbreeding
Bt transgenic rice with non-transgenic rice had a higher grain yield than its non-transgenic hybrid counterpart. In other words,
cry1C* transgenic Minghui-63 (MH63, an elite
indica CMS restorer line) has a significantly lower grain yield than its non-transgenic counterpart MH63 (
Jiang et al. 2013;
Wang et al. 2012b). However, the
cry1C* transgenic hybrid obtained by crossbreeding
cry1C* transgenic MH63 with Zhenshan 97A (an elite
indica CMS line) had a higher grain yield than its non-transgenic hybrid counterpart (non-transgenic MH63 × Zhenshan97A) (
Wang et al. 2012a,
2012b).
Likewise, we adopted a crossbreeding method to develop lines showing field performance similar to that of Korean elite japonica cultivars, with a view to eliminating or minimizing the underlying grain yield cost of cry1Ac/bar transgenic Agb0101. As a result, the grain yield cost of Agb0101 decreased from 27% to 10% of that of the elite cultivars. In particular, the grain yield of the progeny lines derived from crossing with seven elite rice cultivars was statistically equal to that of their non-transgenic counterpart parent.
In conclusion, this is the first report showing that crossbred Bt transgenic japonica rice lines with elite japonica cultivars can be used to minimize the underlying grain cost of transgenic insect-resistant lines. This reduction in the underlying grain cost would contribute to realizing the commercial production of GM rice varieties.
ACKNOWLEDGMENTS
This study was supported by a grant from the National Academy of Agricultural Science (PJ008634) and in part by a grant from the Next-Generation BioGreen 21 Program (Plant Molecular Breeding Center No. PJ011024) funded by the Rural Development Administration, Republic of Korea. The authors have no conflict of interest to declare.
Fig. 1Comparison of growth and yield components of Agb0101, 10 elite cultivars, and their crossbred progeny lines planted in 2010. Values are each ratio of Agb0101, counterpart elite cultivar, or transgenic crossbred progeny to counterpart elite cultivar.
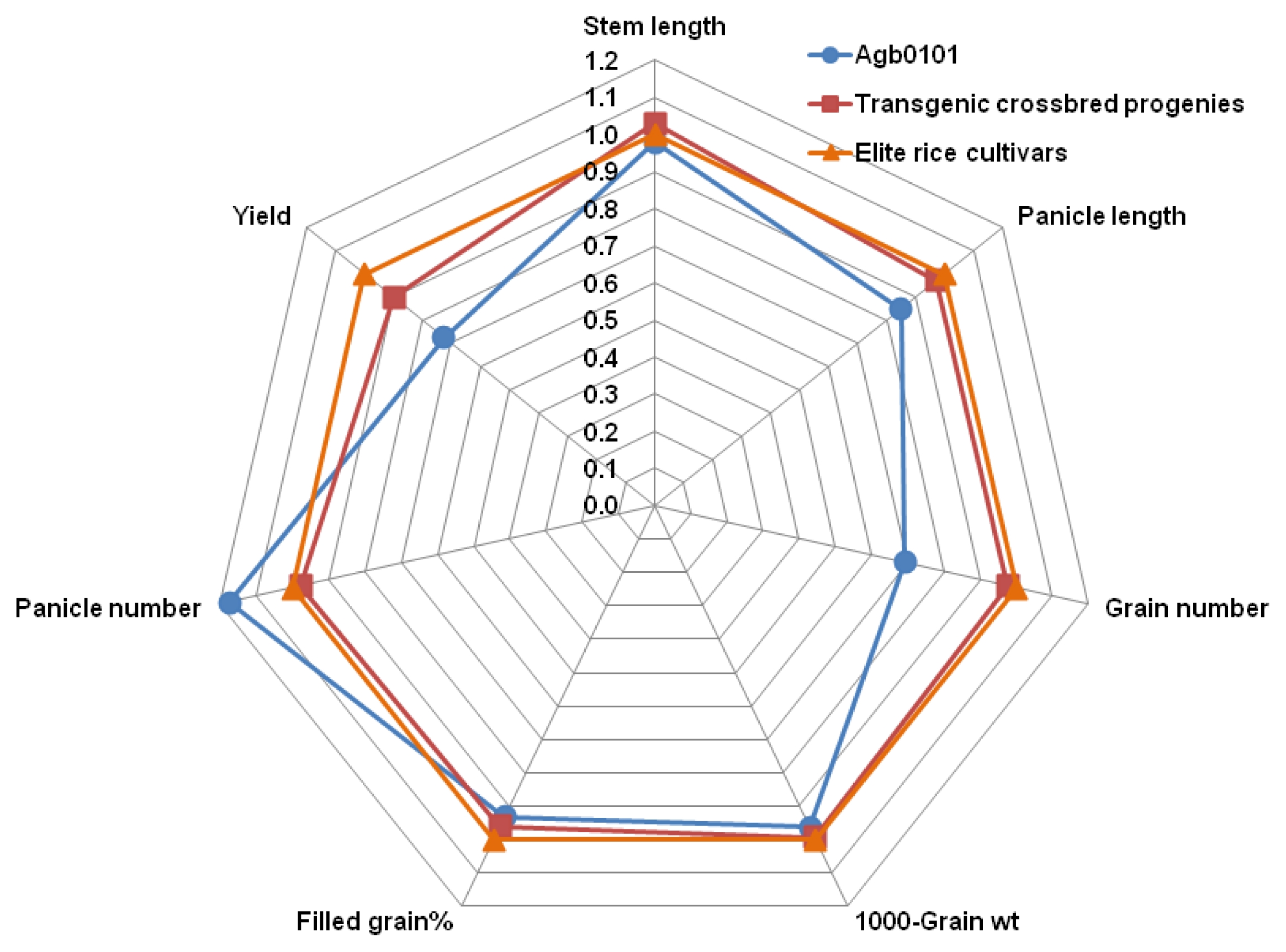
Fig. 2Cluster analysis for crossbred progeny lines between Agb0101 and each of the four elite cultivars in 2010.
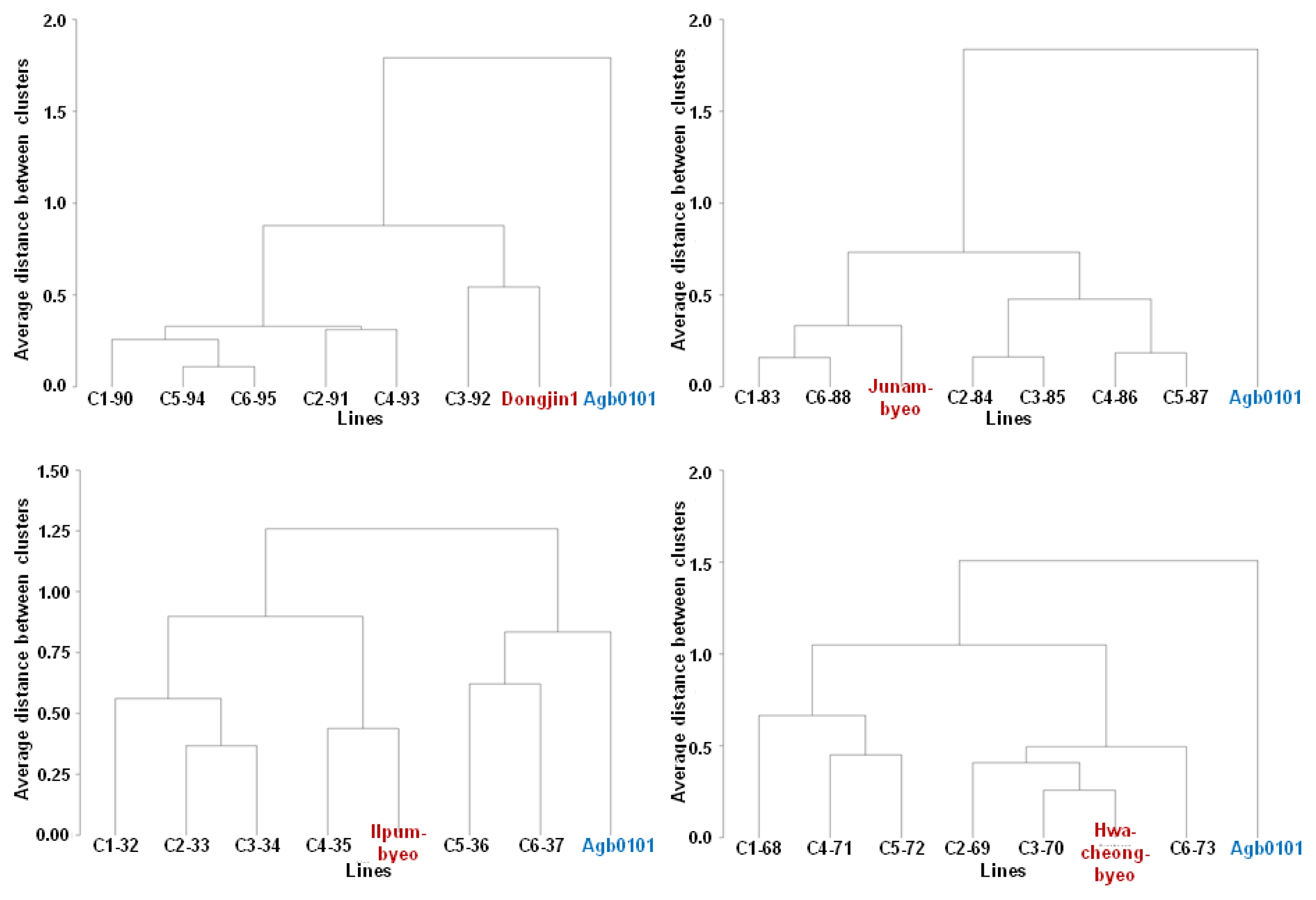
Fig. 3Comparison of grain yield of crossbred progeny lines between Agb0101 and elite cultivars in 2010. Error bar indicates standard error.
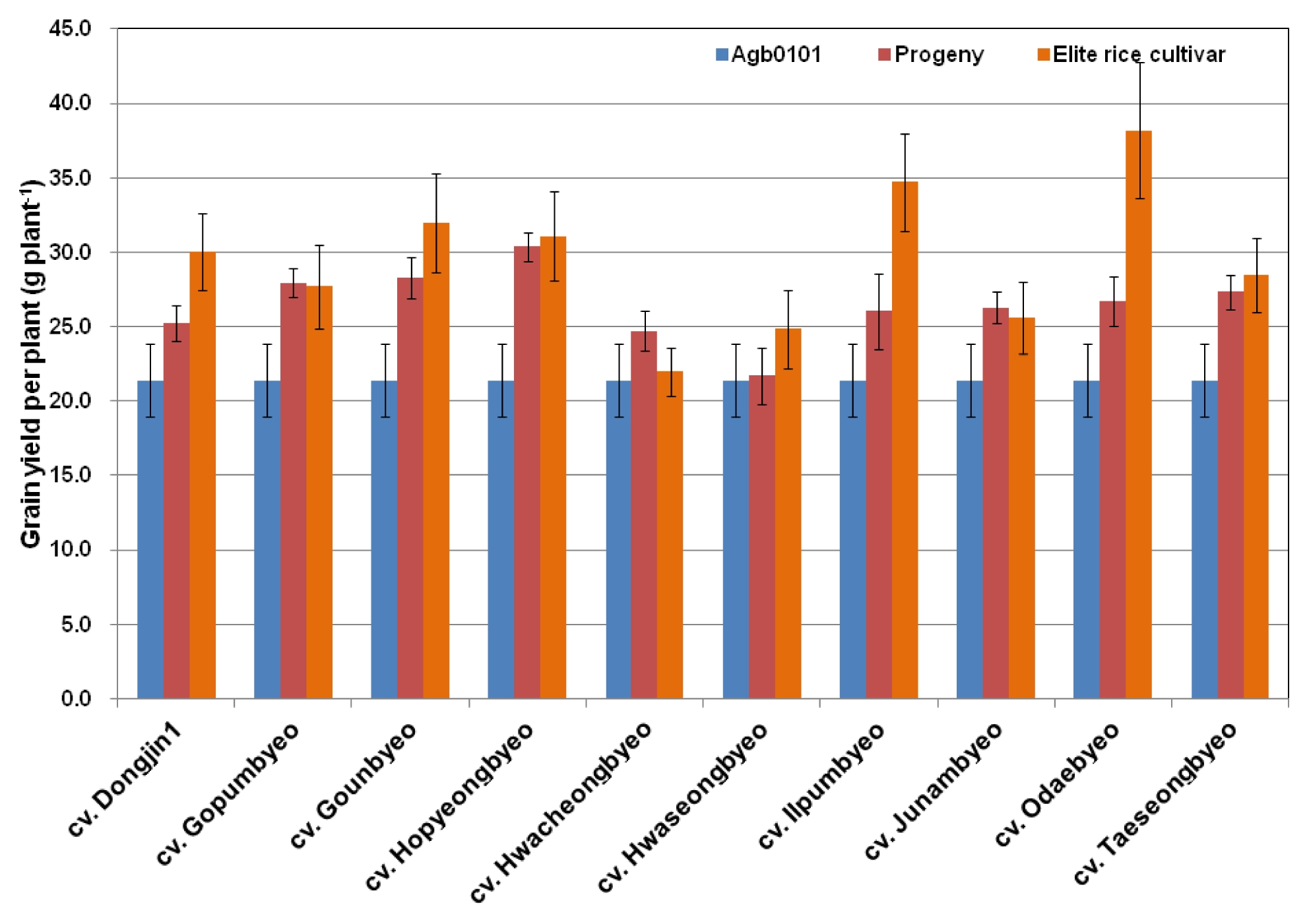
Table 1Growth and yield components of Agb0101 and cv. Nakdongbyeo from 2006 to 2008.
Table 1
|
Traits and grain yield |
Agb0101z)
|
cv. Nakdongbyeoz)
|
|
Stem length (cm) |
74.0 ± 6.0b
|
77.0 ± 5.1a
|
|
Panicle length (cm) |
16.0 ± 1.5b
|
20.0 ± 0.6a
|
|
Panicle number per hill |
13.7 ± 1.5ns
|
13.0 ± 1.2ns
|
|
Filled grain percentage (%) |
91.3 ± 0.6b
|
93.0 ± 0.7a
|
|
Grain number per panicle |
68.3 ± 3.2b
|
85.3 ± 3.5a
|
|
1000 grain weight (g)y)
|
21.9 ± 0.1ns
|
22.0 ± 0.2ns
|
|
Grain yield (kg 10a−1)y)
|
358.3 ± 24.9b
|
437.7 ± 39.1a
|
Table 2Inheritance analysis of cry1Ac/bar transgene in F2 or BC1F1 populations.
Table 2
|
Population |
No. of plants with herbicide response to Basta®z)
|
Expected ratio (R:S)z)
|
χ2 y)
|
P-value |
|
|
Total |
R |
S |
|
F2 populations |
|
cv. Nakdongbyeo/Agb0101 |
1111 |
823 |
288 |
3:1 |
0.504 |
0.478 |
|
|
BC1F1 populations |
|
cv. Odae//Agb0101/cv. Odae |
50 |
23 |
27 |
1:1 |
0.320 |
0.602 |
|
cv. Taeseong//Agb0101/cv. Taeseong |
50 |
24 |
26 |
1:1 |
0.080 |
0.798 |
|
cv. Goun//Agb0101/cv. Goun |
49 |
24 |
25 |
1:1 |
0.020 |
0.932 |
|
cv. Hwaseong//Agb0101/cv. Hwaseong |
46 |
22 |
24 |
1:1 |
0.087 |
0.091 |
|
cv. Ilpum//Agb0101/cv. Ilpum |
48 |
23 |
25 |
1:1 |
0.083 |
0.827 |
|
cv. Hopyeong//Agb0101/cv. Hopyeong |
48 |
23 |
25 |
1:1 |
0.083 |
0.883 |
|
cv. Gopum//Agb0101/cv. Gopum |
30 |
14 |
16 |
1:1 |
0.133 |
0.087 |
Table 3Selection of crossbred Agb0101 progeny lines in 2009 and 2010.
Table 3
|
Crosses of Agb0101 with |
Generationz)
|
No. of lines with all plants resistant to Basta®y)
|
No. of lines selected |
No. of plants selected |
|
2009 |
2010 |
2009 |
2010 |
2009 |
2010 |
2009 |
2010 |
|
cv. Dongjin1 |
F3* |
F4* |
5 (5) |
6 (6) |
2 |
4 |
6 |
12 |
|
|
cv. Gopumbyeo |
F4
|
F5
|
1 (1) |
3 (3) |
1 |
3 |
3 |
9 |
|
BC1F3
|
BC1F4
|
0 (3) |
0 (3) |
1 |
0 |
3 |
0 |
|
|
cv. Gounbyeo |
F4
|
F5
|
1 (1) |
3 (3) |
1 |
3 |
3 |
9 |
|
|
cv. Hopyeongbyeo |
F4
|
F5
|
0 (3) |
2 (3) |
1 |
2 |
3 |
6 |
|
BC1F3
|
BC1F4
|
0 (3) |
0 (3) |
1 |
0 |
3 |
0 |
|
|
cv. Hwacheongbyeo |
F4
|
F5
|
3 (3) |
6 (6) |
2 |
2 |
6 |
6 |
|
|
cv. Hwaseongbyeo |
F4
|
F5
|
0 (1) |
0 (3) |
1 |
0 |
3 |
0 |
|
F3* |
F4* |
1 (1) |
3 (3) |
1 |
1 |
3 |
3 |
|
|
cv. Ilpumbyeo |
F4
|
F5
|
1 (3) |
3 (3) |
1 |
1 |
3 |
3 |
|
BC1F3
|
BC1F4
|
1 (3) |
1 (3) |
1 |
0 |
3 |
0 |
|
F3* |
F4* |
1 (1) |
- |
0 |
- |
0 |
- |
|
|
cv. Junambyeo |
F3* |
F4* |
3 (3) |
6 (6) |
2 |
3 |
6 |
9 |
|
|
cv. Odaebyeo |
F4
|
F5
|
1 (1) |
- |
0 |
- |
0 |
- |
|
BC1F3
|
BC1F4
|
1 (3) |
4 (6) |
2 |
2 |
6 |
6 |
|
|
cv. Taeseongbyeo |
F3* |
F4* |
3 (3) |
- |
0 |
- |
0 |
- |
|
BC1F3
|
BC1F4
|
1 (3) |
5 (6) |
2 |
2 |
6 |
6 |
References
- Chen DH, Ronald PC. 1999. A rapid DNA mini preparation method suitable for AFLP and other PCR applications. Plant Mol Biol Rep. 17: 53-57.
- Fujimoto H, Itoh K, Yamamoto M, et al. 1993. Insect resistant rice generated by introduction of a modified δ-endotoxin gene of Bacillus thuringiensis. Nat Biotechnol. 11: 1151-1155.
- Garris AJ, Tai TH, Coburn J, et al. 2005. Genetic structure and diversity in Oryza sativa L. Genetics. 169: 1631-1638.
- Heong KL, Escalada MM, Mai V. 1994. An analysis of insecticide use in rice: case studies in the Philippines and Vietnam. Intl J Pest Manage. 40: 173-178.
- High SM, Cohen MB, Shu QY, et al. 2004. Achieving successful deployment of Bt rice. Trends Plant Sci. 9: 286-292.
- Huang J, Hu R, Rozelle S, et al. 2005. Insect-resistant GM rice in farmers’ fields: assessing productivity and health effects in China. Science. 308: 688-690.
- Jiang Y, Huang S, Cai M, et al. 2013. Yield changes of Bt-MH63 with cry1C*or cry2A*genes compared with MH63 (Oryza sativa) under different nitrogen levels. Field Crop Res. 151: 101-106.
- Kim EH, Suh SC, Park BS, et al. 2009. Chloroplast-targeted expression of synthetic cry1Ac in transgenic rice as an alternative strategy for increased pest protection. Planta. 230: 397-405.
- Kim S, Kim C, Li W, et al. 2008. Inheritance and field performance of transgenic Korean Bt rice lines resistant to rice yellow stem borer. Euphytica. 164: 829-839.
- Lee GH, Paik CH, Choi MY, et al. 2006. Economic injury levels for the rice leaffolder Cnaphalocrocis medinalis ( Lepidoptera: Pyralidae) in the rice paddy field. Korean J Appl Entomol. 45: 327-331.
- Lee RY, Shin KS, Suh SC, et al. 2009. Molecular characterization of lepidopteran pest-resistant transgenic rice events expressing synthetic cry1Ac. Plant Biotechnol Rep. 3: 317-324.
- Lu BR, Snow AA. 2005. Gene flow from genetically modified rice and its environmental consequences. BioScience. 55: 669-678.
- Nathan SS. 2006. Effects of Melia azedarach on nutritional physiology and enzyme activities of the rice leaffolder, Cnaphalocrocis medinalis. Pestic Biochem Physiol. 84: 98-108.
- Shu Q, Cui H, Ye G, et al. 2002. Agronomic and morphological characterization of Agrobacterium-transformed Bt rice plants. Euphytica. 127: 345-352.
- Shu Q, Ye G, Cui H, et al. 2000. Transgenic rice plants with a synthetic cry1Ab gene from Bacillus thuringiensis were highly resistant to eight lepidopteran rice pest species. Mol Breed. 6: 433-439.
- Tu J, Zhang G, Datta K, et al. 2000. Field performance of transgenic elite commercial hybrid rice expressing Bacillus thuringiensis δ-endotoxin. Nat Biotechnol. 18: 1101-1104.
- Wang F, Jian Z, Nie L, et al. 2012a. Effects of N treatments on the yield advantage of Bt-SY63 over SY63 (Oryza sativa) and the concentration of Bt protein. Field Crops Res. 129: 39-45.
- Wang F, Ye C, Zhu L, et al. 2012b. Yield differences between Bt transgenic rice lines and their non-Bt counterparts, and its possible mechanism. Field Crops Res. 126: 8-15.
- Wang Y, Zhang G, Du J, et al. 2010. Influence of transgenic hybrid rice expressing a fused gene derived from cry1Ab and cry1Ac on primary insect pests and rice yield. Crop Prot. 29: 128-133.
- Wang YQ, Johnston S. 2007. The status of transgenic rice R&D in China. Nat Biotechnol. 25: 717-718.
- Wang Z, Shu Q, Ye G, et al. 2002. Genetic analysis of resistance of Bt rice to stripe stem borer (Chilo suppressalis). Euphytica. 123: 379-386.
- Won YJ, Yi GH, Cho JH, et al. 2004. Establishment of a new breeding scheme for rapid release of variety using bar gene transformed rice. Korean J Plant Biotechnol. 31: 7-11.
- Wu G, Cui H, Ye G, et al. 2002. Inheritance and expression of the cry1Ab gene in Bt (Bacillus thuringiensis) transgenic rice. Theor Appl Genet. 104: 727-734.
- Xia H, Chen L, Wang F, et al. 2010. Yield benefit and underlying cost of insect-resistance transgenic rice: implication in breeding and deployment of transgenic crops. Field Crops Res. 118: 215-220.
- Xia H, Lu BR, Xu K, et al. 2011. Enhanced yield performance of Bt rice under target-insect attacks: implications for field insect management. Transgenic Res. 20: 655-664.
- Yea GY, Shu QY, Yao HW, et al. 2001. Field evaluation of resistance of transgenic rice containing a synthetic cry1Ab gene from Bacillus thuringiensis Berliner to two stem borers. J Econ Entomol. 94: 271-276.
- Yu J, Hu S, Wang J. 2002. A draft sequence of the rice genome. Science. 296: 79-92.