Abstract
Grain size is one of the most important factors determining grain yield in rice breeding. In previous studies, we constructed high-density maps for two quantitative trait loci (QTL) for grain weight, tgw2 and gw9.1, using progeny derived from crosses between the japonica cultivar Hwaseong and Oryza grandiglumis, and Hwaseong and O. rufipogon (IRGC 105491), respectively. The wild alleles contributed an increase in grain weight at these two loci. We developed an F2 population (146 plants) by crossing two near isogenic lines (NILs) harboring tgw2 and gw9.1 to know how they interact in the near isogenic background. Simple sequence repeat markers tightly linked to two QTL were used to check the genotype of the F2 population. Based on the genotype at two loci, 146 F2 plants were classified into 9 groups with a combination of three genotypes at each two loci. Two gene interaction was not significant (P=0.99) in the F2. Homozygous plants with wild alleles at two loci showed significantly higher 1,000 grain weight than plants with a single QTL in the F2 and F3. These results indicate that two QTLs act additively in distinct or complementary pathways in controlling GW. Gene expression analysis was also performed to know the relationship of the gw9.1 QTL with three major grain size genes with Hwaseong and two NILs plants at the transcription level. The results from this study provide insight into grain size regulation in rice and are likely to be useful for marker aided selection for grain size.
-
Key words: Rice, Grain weight, Quantitative trait loci, Interaction, Near isogenic line
INTRODUCTION
Rice (
Oryza sativa L.) is one of the most important cereal food crop. Rice yield is closely related with grain weight. Grain weight shows a positive correlation with grain size which is measured by grain length (GL), grain width (GW), and grain thickness (GT). The identification of major quantitative trait loci (QTL) for grain shape is an important objective of rice genetic research and breeding program (
Kang et al. 2011;
Huang et al. 2013;
Nagata et al. 2015). Extensive studies were carried out to identify QTL for grain size and nearly 200 QTL for GL and GW have been reported in rice (
Nagata et al. 2015). More than ten genes to control grain size have been cloned and molecularly characterized,
GS3 (
Fan et al. 2006),
GW2 (
Song et al. 2007),
qGL3 (
Hu et al. 2012),
GS5 (
Li et al. 2011),
qSW5 (
Shomura et al. 2008),
GW8 (
Wang et al. 2012),
HGW (
Li et al. 2012),
TGW6 (
Ishimaru et al. 2013),
GW6a (
Song et al. 2015), and
GS6 (
Sun et al. 2013).
GW2 encodes an E3 ubiquitin ligase and affects GW by restricting cell division. A significant increase of 49.8% in grain weight was observed in the near isogenic line (NIL)-
GW2 compared with the control, FAZ1-NIL (
Song et al. 2007).
GS5 encodes a putative serine carboxypeptidase and functions as a positive factor for GW (
Li et al. 2011).
SPL16 (
GW8) promotes cell division leading to increase in GW and showed a 13.9% advantage in 1,000 grain weight (TGW;
Wang et al. 2012). These studies improved our understanding of genetic control over grain size traits; however the employment of these major QTL in practical breeding program for enhancing rice yield potential is hampered. This is because the information of additive and epistatic interactions across multiple grain size genes essential for combining favorable alleles resulting in enhanced traits is not sufficient for fine-tuning grain shape in practical breeding programs (
Huang et al. 2013). In this regard, it is necessary to decipher the interactions among grain size genes and combine appropriate genes/QTLs.
QTL-NILs are promising materials to detect interactions among genes and to characterize gene actions in details because the background genetic effects on the analysis can be minimized in QTL-NILs. However, a few studies reported the employment of QTL-NIL for analyzing the interaction among grain size QTL.
Jin et al. (2011) detected an epistatic interaction between two QTL,
gw8.1 and
gw9.1.
Wang et al. (2012) also examined the interaction between
GS3 and
qGW8 in a near-isogenic (HJX74) background.
Lu et al. (2013) compared the grain size among haplotypes of
GW2,
GS3,
qSW5, and
GS5 in the Zhanshan97 background. However, detailed analysis of interaction among grain shape QTLs will provide more insights into the molecular mechanisms underlying seed size development in rice and is likely to be useful for gene pyramiding aimed at improving rice grain yield.
In our previous studies, two QTL-NILs,
tgw2-NIL (
Yoon et al. 2006) and
gw9.1-NIL (
Xie et al. 2008) related with grain size were developed by introgressing chromosomal segments of two wild species,
O. grandiglumis (IRGC 101154) and
O. rufipogon (IRGC 105491) into the Korean elite
japonica cultivar Hwaseong, respectively. This study was carried out to elucidate the interaction of two QTL,
tgw2 and
gw9.1. Also, to know the relationship of the
gw9.1 QTL with three major grain size genes
GW2,
GS3, and
HGW at the transcription level, gene expression analysis was performed with three lines, Hwaseong and two NILs.
MATERIALS AND METHODS
Plant materials
Molecular marker-assisted backcrossing was used to pyramid two QTL,
tgw2 and
gw9.1. 146 F
2 plants were produced from a cross between two QTL-NILs and genotyped using simple sequence repeat (SSR) markers tightly linked to the two QTL. Four F
2 plants with 1 of 4 types of QTL-NILs: Hwaseong (Hwaseong homozygous at both loci),
tgw2-NIL (
O. grandiglumis homozygous at
tgw2 and Hwaseong homozygous at
gw9.1),
gw9.1-NIL (Hwaseong homozygous at
tgw2 and
O. rufipogon homozygous at
gw9.1), and
tgw2+
gw9.1-NIL (
O. grandiglumis and
O. rufipogon homozygous at
tgw2 and
gw9.1) were selected and advanced to F
3 lines (
Fig. 1).
During the summer of 2013, 146 F
2 plants were grown in the experimental farm, Chungnam National University, Daejeon, Korea at a density of 15×30 cm (line×row). Seeds were sown on April 25 and transplanted to the field on May 29. To minimize border effect, seedlings of F
2 plants were planted in the middle, excluding the first five and last five lines in each row. 146 F
2 plants were measured for GL, width, thickness and TGW in brown rice. Fully-filled seeds were re-dried in an oven at 30°C for 24 hours. TGW was evaluated by measuring the weight of 250 randomly-selected brown rice; this method was performed in triplicates and the values were averaged to yield a single mean. Four F
3 lines selected from the F
2 population were grown in the field during the summer of 2014. The planting density was the same as 2013. Four F
3 lines were evaluated for 11 traits (culm length [CL], panicle length [PL], panicle number [PN], spikelets per panicle [SPP], fertility [FER]). Each line was represented by a row of 30-day-old seedlings planted with 15 cm between plant and 30 cm between rows in a completely randomized block design with two replications. CL was measured in centimeters from the surface of soil to panicle neck. PL was measured in centimeters from panicle neck to the tip of panicle. PN was calculated as the number of panicles per plant. For SPP, all seeds on panicle were counted including unfilled seeds. For FER, filled seeds were counted and divided with SPP (%). The middle 10 plants from each line were chosen for the evaluation, and the average of the measurements was used for the phenotype of each line for the TGW. Grains with hulls were allowed to dry naturally after harvesting, seeds were re-dried in an oven at 30°C for 24 hours. The TGW was evaluated by measuring the weight of 250 randomly selected brown rice per plant (10 plants per line). GL, GW, and GT were measured. GL, GW, and GT were measured for 15 grains (brown rice) per plant (10 plants per line) using a 150-mm Vernier caliper (Mitutoyo Corp., Kawasaki, Japan). TGW was corrected for the 15% grain moisture content. Amylose content (%) and grain chalkiness were checked using a category of 0 (no chalky in the center and belly) to 9 (the grain opaque area >75%) (
Juliano 1985). Means for two replications were evaluated for each trait and used in data analysis.
DNA was extracted from the fresh leaf tissue of each of 146 F
2 plants. DNA extraction was performed as described in
Causse et al. (1994). SSR analysis of the F
2 population was performed according to the method described by
Panaud et al. (1996). SSR markers tightly linked to two QTLs, RM12813 for
tgw2 and ex9 for
gw9.1 (
Xie et al. 2008) were used for genotyping 146 F
2 plants. Protocols for marker amplification using the polymerase chain reaction (PCR) and for size separation using polyacrylamide gel electrophoresis and marker detection using the silver staining procedure were as described in
Xie et al. (2008). The silver staining kits were purchased from Bioneer Co., Daejeon, Korea (
www.bioneer.co.kr).
Total RNA was extracted from panicles, less than 1 cm long from plants around 23 days before heading; Hwaseong,
tgw2-NILs, and
gw9.1-NILs using Nucleospin RNA plant kit (MACHEREY-NAGEL, Düren, Germany). Sampling time and procedure followed those described in previous studies (
Song et al. 2007;
Mao et al. 2010;
Li et al. 2012). The first-strand cDNA was synthesized using extracted RNA using a PrimeScript RT-PCR Kit (TaKaRa Biotechnology Co. Ltd, Tokyo, Japan). Gene expression was performed by quantitative Real-Time PCR (CFX Connect Real-Time System; Bio-Rad, Hercules, CA, USA) with IQ
™ SYBR
® Green Supermix (BIO-RAD). Quantitative RT-PCR was done by using 3 steps protocol; 95°C for 1 minute for denaturation, 40 cycles of 95°C for 15 seconds (denaturation), 55°C for 30 seconds (annealing) and 72°C for 30 seconds (extension), followed by final extension at 72°C for 7 minutes, and melting curve, temperature increased from 72°C to 95°C by 0.2°C for 5 seconds. For qRT-PCR primers,
OsUBQ5 (
Oryza sativa Ubiquitin 5) was used as control and three primers for qRT-
GW2 (
Yan et al. 2011;
Lu et al. 2013), qRT-
GS3 (
Yan et al. 2011), and
HGW were designed (
Table 1). Gene expression was performed in three replicates and the average values of three replicates were used in data analysis.
To analyze the interaction effect for grain weight, nine genotypes were identified from 146 F2 plants based on SSR genotypes tightly linked to each target QTL. In the F2 generation, QTL was declared if the phenotype was associated with a marker locus at P<0.05 using one-way ANOVA. Two-way ANOVA was used to analyze the epistatic interaction between two QTL. The proportion of observed phenotypic variation attributable to a particular QTL was estimated by the coefficient of determination (R2). The mean values of four QTL-NILs in the F3 were compared using Duncan’s multiple range test using SAS 9.3 program (SAS Institute, Cary, NC, USA).
RESULTS
QTL Confirmation
The existence of two QTL was confirmed by QTL analysis of the F
2 population (
Table 2,
Fig. 2). The
O. grandiglumis and
O. rufipogon alleles increased grain weight in the Hwaseong background which is consistent with those in earlier reports (
Yoon et al. 2006;
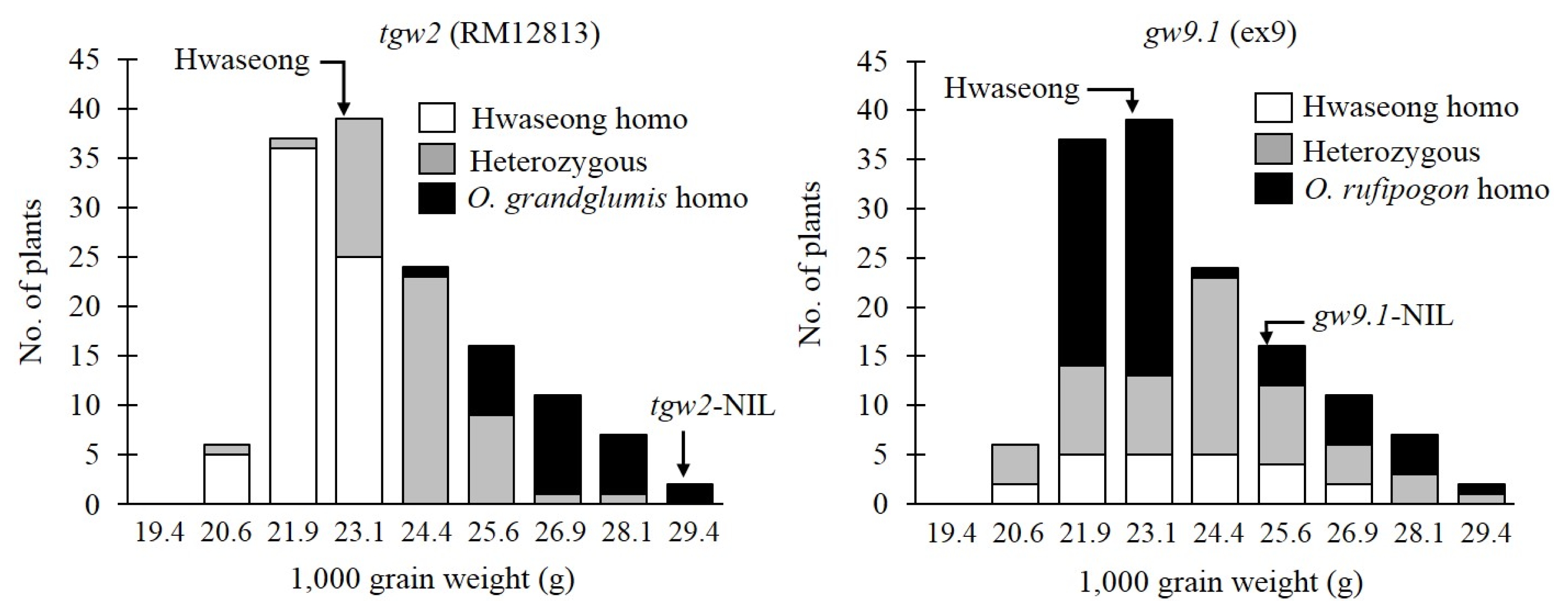
Xie et al. 2008). The genotype-based distribution of the grain weight in the F
2 at the two loci is shown in
Fig. 2. Grain weight of two NILs for
tgw2 and
gw9.1 was 29.6 g and 24.7 g, respectively, and significantly higher than that of Hwaseong-NIL (23.0 g) (
P<0.05). At the
tgw2 locus, the mean TGW of the homozygous
O. grandiglumis class was significantly higher than those of the heterozygous and Hwaseong homozygous class. There was also a significant difference between heterozygote plants and Hwaseong homozygote plants (
Table 2). This result indicated that the
O. grandiglumis allele increased TGW in the Hwaseong background in an additive fashion.
TGW of the homozygous
O. rufipogon class at
gw9.1 was significantly higher than the Hwaseong homozygous class. However, no significant difference between TGW of the homozygous
O. rufipogon class and the heterozygous class at
gw9.1 was detected. The finding that
tgw2 and
gw9.1 explained 73.5% and 38.8% of the phenotypic variance for TGW in the F
2, respectively confirms the existence of the two QTLs (
Table 2).
Based on the genotypes at the two loci, RM12813 for
tgw2 and ex9 for
gw9.1, 146 F
2 plants were classified into nine groups with a combination of three genotypes (homozygous for Hwaseong,
O. rufipogon/
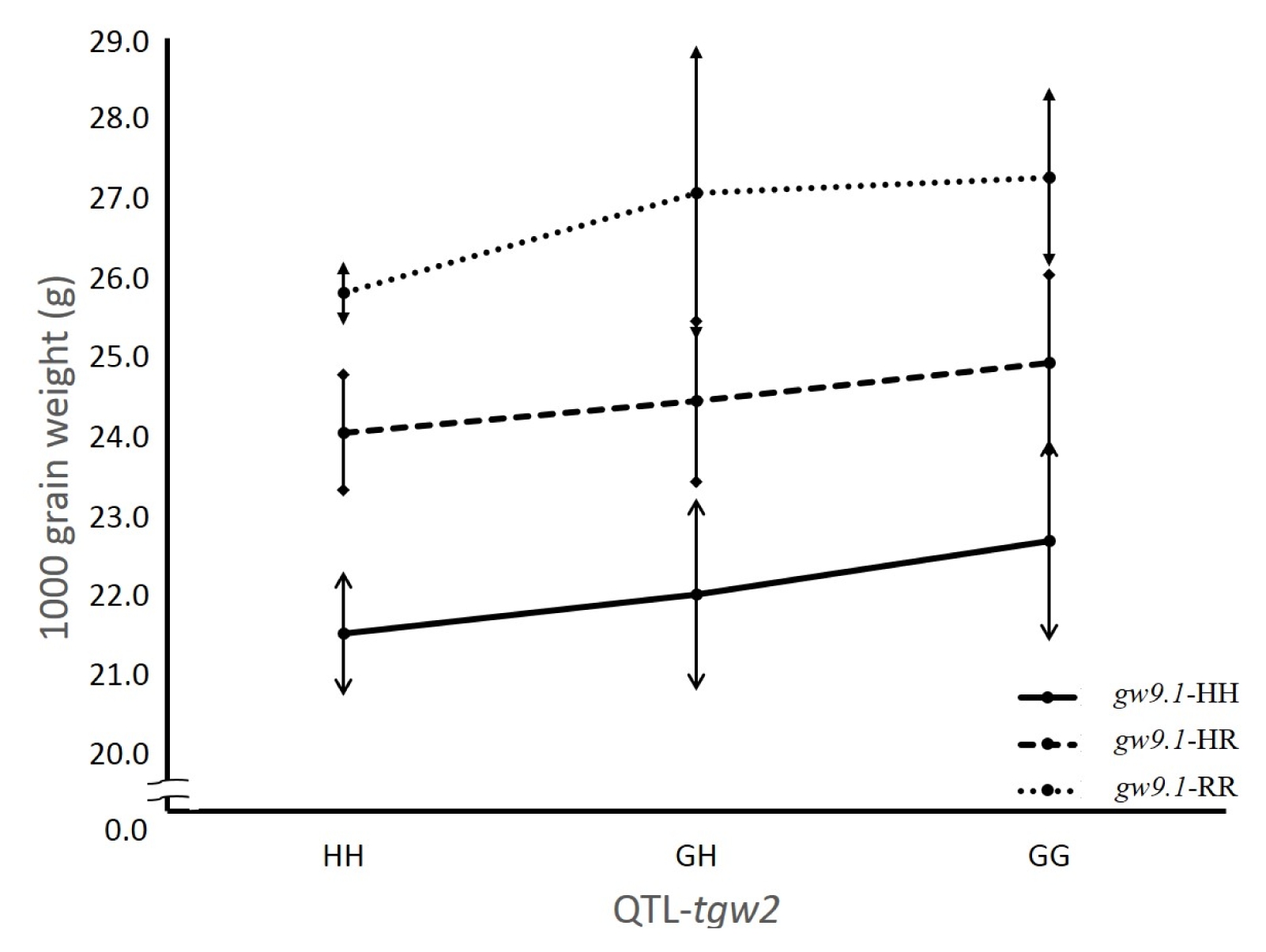
O. grandiglumis or heterozygous) at each locus. To test the pyramiding effect of the two QTL, the average grain weight of nine genotype classes in the F
2 population was compared. Digenic interaction was not detected based on two-way ANOVA (
P=0.99) (
Fig. 3). The TGW increasing effect of the
O. grandiglumis allele was observed in three genotype classes at the
tgw2 locus. Also, the
O. rufipogon allele increased TGW in three genotype classes at the
gw9.1 locus. The highest grain weight was observed in genotype class homozygous for
O. grandiglumis and
O. rufipogon at both loci
tgw2 and
gw9.1 indicating no epistatic interaction between
tgw2 and
gw9.1. To further confirm this finding that no interaction exists between
tgw2 and
gw9.1, grain weight of four QTL-NILs was measured in the F
3 (
Table 3). Four F
3 lines were selected from the F
2 population based on the genotypes at two loci (
Fig. 1). The effect of the
O. rufipogon allele at
gw9.1 on TGW was observed based on comparison of grain weight between Hwaseong and
gw9.1-NIL. The effect of the
O. grandiglumis allele at
tgw2 on TGW was observed based on comparison of grain weight between Hwaseong and
tgw2-NIL plants. Also, the additive effect of the
O. grandiglumis allele at
tgw2 and the
O. rufipogon allele at
gw9.1 on TGW was observed based on comparison of grain weight between
tgw2-NIL and
tgw2+
gw9.1-NIL. TGW difference between
tgw2-NIL and
tgw2+
gw9.1-NIL caused by the genotype of
gw9.1 was observed. The finding indicates that there is no interaction between
tgw2 and
gw9.1. To clarify which grain shape trait was associated with the increase in TGW, NILs were evaluated for GL, GW, and GT (
Table 3,
Fig. 4). GL and width of
tgw2+
gw9.1-NIL plants were significantly higher than those of
tgw2-NIL and
gw9.1-NIL plants. A phenotypic difference between
tgw2-NIL and
tgw2+
gw9.1-NIL caused by the
O. rufipogon allele at
gw9.1 was observed. Similar results were observed for GT: grains of the
tgw2-NIL and
tgw2+
gw9.1-NIL plants were significantly thicker than
gw9.1-NIL grains. These results indicate that the variation in grain weight in
gw9.1-NIL is associated with GL and width whereas three grain shape traits are responsible for the variation in GW in
tgw2-NIL.
To know the effect of the chromosomal segment harboring
tgw2 and
gw9.1 from
O. grandiglumis and
O. rufipogon on agronomic traits, NILs were evaluated for 11 agronomic traits (
Table 3). Results showed that no significant difference was observed for CL and SPP. PL was significantly higher in
gw9.1-NIL than the other NILs and Hwaseong. However,
tgw2-NIL and
tgw2+
gw9.1-NIL plants had significantly lower number of panicles per plant than Hwaseong and this result seems to imply that QTL associated with panicle number is located in the
O. grandiglumis introgression on chromosome 2 and is consistent with the previous study (
Yoon et al. 2006). Also,
tgw2-NIL and
tgw2+
gw9.1-NIL plants displayed higher ratio of grain chalkiness and amylose content than Hwaseong.
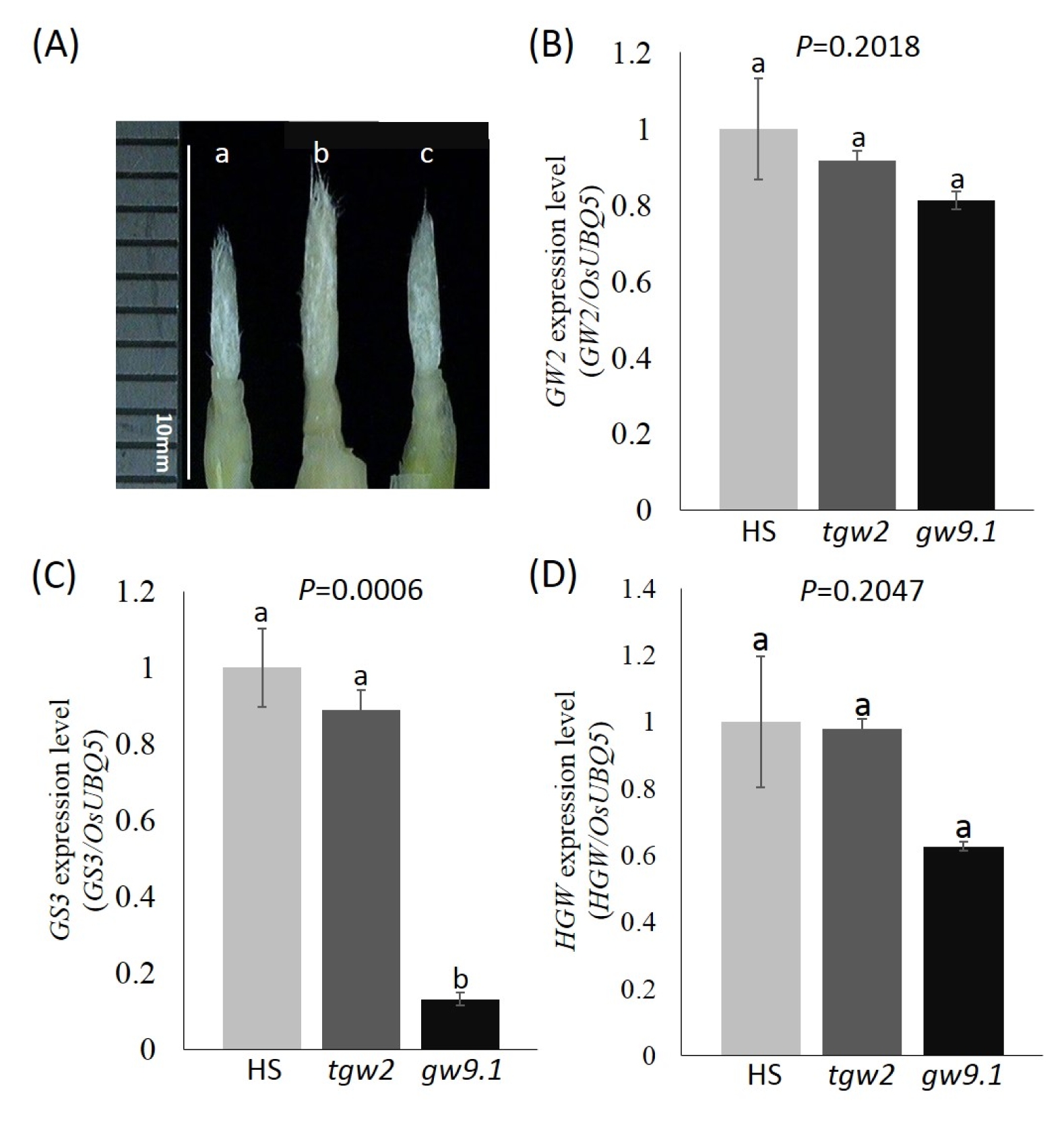
To investigate whether
gw9.1 interacts with other three grain size genes,
GW2,
GS3, and
HGW, we performed the quantitative RT-PCR with three groups: Hwaseong,
tgw2-NIL and
gw9.1-NIL (
Fig. 5). For
GW2 expression (
Fig. 5B), three groups did not show any significant difference supporting the results from phenotypic data that
tgw2 and
gw9.1 act in an additive manner. A significant difference was detected in
GS3 expression level between Hwaseong and
gw9.1-NIL, and
tgw2-NIL and
gw9.1-NIL (
Fig. 5C). The difference between Hwaseong and
tgw2-NIL in the expression level of
GS3 was observed (
P=0.189) although it did not reach the significance level (
P<0.05). The expression of
GS3 in
gw9.1-NIL was remarkably reduced compared to other two groups. This result might imply that the expression of
GS3 may be negatively regulated by
gw9.1. However additional experiments are needed to confirm the effect of
gw9.1 on the expression of
GS3 because the molecular function of
gw9.1 is not known and
gw9.1-NIL harbors several
O. rufipogon genes in the introgressed segment (
Xie et al. 2008). It is speculated that
GS3 expression was affected by other
O. rufipogon genes in the introgression in
gw9.1-NIL. To analyze the interaction between
gw9.1 and
GS3, cloning of the
gw9.1 gene is necessary. No significant difference among three groups was observed in
HGW expression (
Fig. 5D).
DISCUSSION
Cloning and molecular characterization of several major QTL for grain shape traits have advanced our understanding of the grain size regulation in rice. However, the complicated additive and epistatic interactions among major grain size genes and some minor QTL are still unclear, and these impede the process of gene pyramiding program to attain higher grain yield in rice (
Huang et al. 2013). In rice, primary mapping populations have been extensively used in QTL analyses (
Yano and Sasaki 1997). It would be more difficult to determine the true genetic action of the QTL because the segregation of other QTLs often affects the genetic parameters of a given QTL in primary mapping population (
Yano and Sasaki 1997). Only a limited number of interactions between QTLs were detected because primary populations cannot provide much information about the real nature of interactions among QTLs due to background noise. For example, two studies reported different results on the interaction between QTLs for grain size. Major QTL,
GW2, and
qSW5/GW5 for grain size were detected using an F
2 population from a cross between ‘FAZ1’ and ‘Jizi1560’ and no significant interaction was observed between three QTL,
GW2,
GS5, and
qSW5/GW5 (
Ying et al. 2012). However, gene expression analysis with
GW2-RNAi lines and CSSL of
qSW5 at the transcriptional level clearly showed that
qSW5 can be down-regulated by repression of
GW2 transcription implying that
GW2 might work upstream of
qSW5 (
Yan et al. 2011). Also, in an association study using
japonica rice, a significant difference in GL was detected between two
GW2 genotypes in the presence of nonfunctional
GS3-A alleles, and the result indicated that two QTL,
GW2 and
GS3 interact each other (
Lu et al. 2013).
QTL-NIL could provide a solution to this problem because the background genetic effects on the analysis can be minimized by QTL-NIL or populations derived from crosses between QTL-NILs with a uniform genetic background.
Jin et al. (2011) detected an epistatic interaction between
gw8.1 and
gw9.1 using four QTL-NILs with allelic combination of
gw8.1 and
gw9.1 in a near-isogenic (Hwaseong) background.
Wang et al. (2012) also developed four contrasting allelic combinations of
GS3 and
qGW8 in a near-isogenic (HJX74) background and examined the interaction between
GS3 and
qGW8. NIL-
GW8/gs3 plants harboring the
qgs3 allele from Basmati385 had grains heavier and longer than those from NIL-
GW8/GS3 control plants and this result implies that two QTL act in an additive manner. These results clearly indicate that utilization of QTL-NIL is a powerful strategy for analyzing the gene interaction of QTLs.
In this study, we described the pyramided effect of
gw9.1 with
tgw2 on agronomic traits. The results indicated that
tgw2 showed no interaction with another grain weight gene,
gw9.1. Compared with TGW of Hwaseong,
tgw2 from
O. grandiglumis increased GW by up to 28%,
gw9.1 from
O. rufipogon by up to 7.4%, and both by up to about 38.3%. TGW of three QTL-NILs,
tgw2-NIL,
gw9.1-NIL and
tgw2+
gw9.1-NIL were significantly higher than that of Hwaseong. TGW of
tgw2+
gw9.1-NIL plants was significantly higher than those of
gw9.1-NIL and
tgw2-NIL plants. These results indicate that two QTLs act additively in distinct or complementary pathways in controlling grain weight. Analysis of grain shape traits suggests that the increase in GL might be responsible for increased grain weight in
gw9.1-NIL whereas the increase in GL, width and thickness is associated with increased weight in
tgw2-NIL and
tgw2+
gw9.1-NIL. A drastic increase in grain size due to
tgw2 did not increase grain yield proportionally in
tgw2-NIL and
tgw2+
gw9.1-NIL plants owing to decrease in grain filling and panicles per plant (data not shown) and this is a typical example of imbalance between the sink and source potentials (
Takai et al. 2013). Also, grain quality traits (high rate of white center and white belly, higher amylose content) were observed in
tgw2-NIL and
tgw2+
gw9.1-NIL plants compared to Hwaseong. It has to be determined whether this deleterious effect is due to a pleiotropy of the
tgw2 QTL or its linkage with other genes. This result suggests that breeders should practice caution when choosing QTL alleles for grain shape with pleiotropy or tight linkage with genes for deleterious effects.
We performed the quantitative RT-PCR with three groups, Hwaseong,
tgw2-NIL, and
gw9.1-NIL to investigate the interactions among grain size genes,
GW2,
GS3, and
HGW. As for
GW2 expression, no significant difference among three groups was detected supporting the result of the phenotype data. For
GS3 expression, the difference between Hwaseong and
tgw2-NIL was observed (
P=0.189) although it did not reach the significance level (
P<0.05). This result is not consistent with the results from the study by
Yan et al. (2011) reported that
GS3 was positively regulated by
GW2 based on the gene expression level of
GW2 in both RNAi-
GS3 and RNAi-
GW2. Additional experiments are needed to analyze their interactions using QTL-NILs in more details at the phenotypic level. A significant difference between Hwaseong and
gw9.1-NIL, and
tgw2-NIL and
gw9.1-NIL were observed and these results suggest a possibility that
gw9.1 might affect the expression of
GS3. For
HGW, there was no significant difference among three groups.
Li et al. (2012) analyzed the
GW2 expression level in
HGW mutants and found that the
GW2 expression level in
HGW mutants were significantly reduced compared to wild type (Zhounghua 11). The finding from this study that no significant difference between Hwaseong and
tgw2-NIL in
HGW expression might support the finding that
HGW might be a upstream regulator of
GW2 (
Li et al. 2012).
To elucidate the interaction of two QTLs, gw9.1 and tgw2, an F2 population was developed by crossing two NILs harboring tgw2 and gw9.1. SSR markers tightly linked to two QTLs were used to check the genotype of the F2 population and to select four F3 QTL-NILs with a combination of two QTLs. Two-way ANOVA revealed no interaction between 2 QTLs in the F2 population. TGW of homozygous plants with wild alleles at two loci was significantly higher than that of plants with a single QTL in the F2 and F3. These results indicate that two QTLs act additively in distinct or complementary pathways in controlling grain weight. Although the double-QTLs line gave further advantage in increasing TGW in the Hwaseong background, an increase in grain weight did not result in an increase in grain yield. However, it should be cautioned that grain filling and grain quality traits of the double-QTLs plants were decreased compared to Hwaseong indicating that tgw2 may have some effect on plant morphology. It has to be determined whether this deleterious effect is due to a pleiotropy of the tgw2 QTL or linkage of tgw2 with other genes.
ACKNOWLEDGEMENTS
This work was carried out with the support of “Golden Seed Project” (Project No. PJ00993301), Ministry of Agriculture, Food and Rural Affairs (MAFRA), Ministry of Oceans and Fisheries (MOF), Rural Development Administration (RDA) and Korea Forest Service (KFS), Republic of Korea.
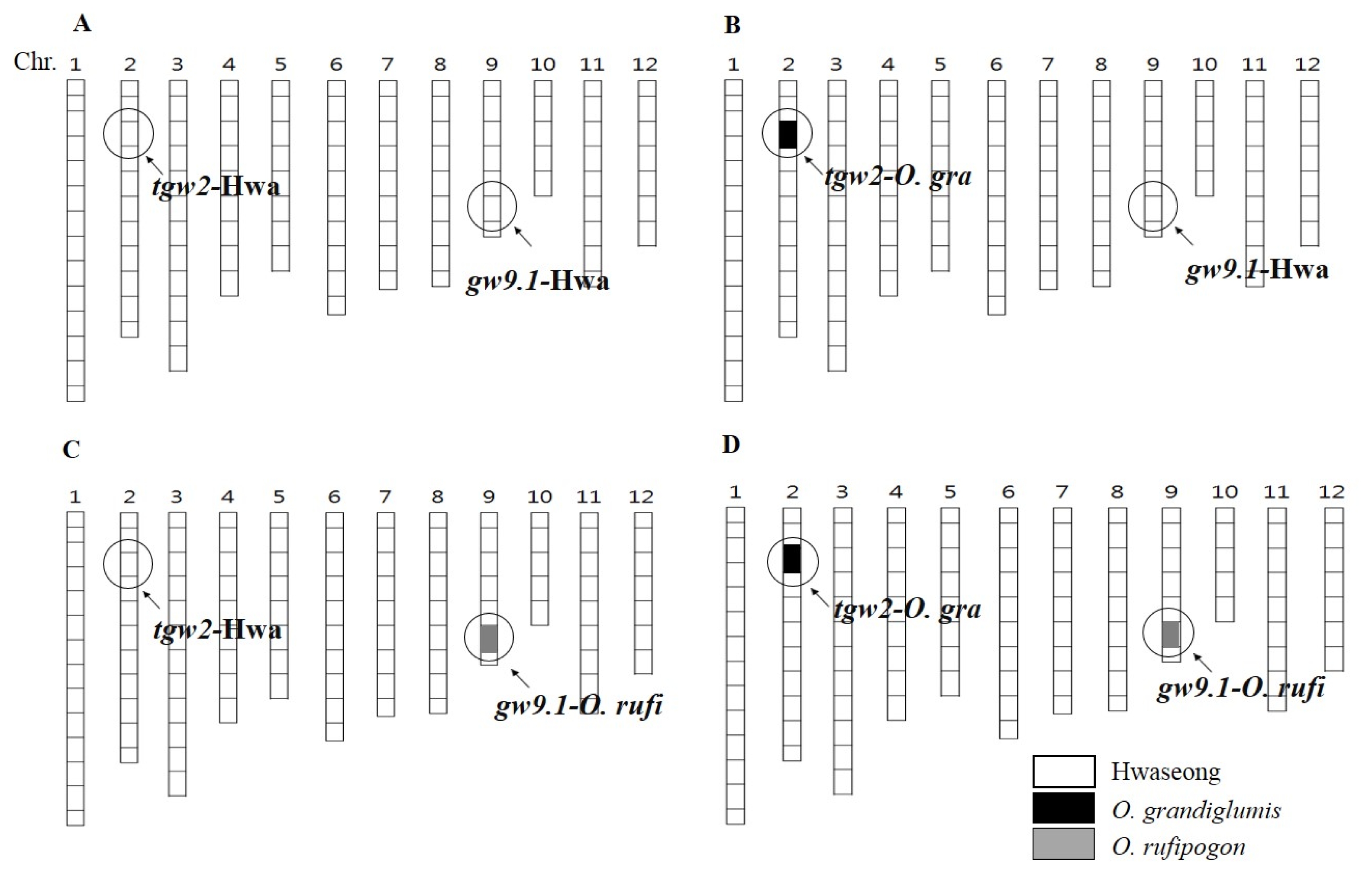
Fig. 1
Graphical genotypes of 4 representative groups. The two circles on the chromosomes represent regions for 2 quantitative trait loci. Open bars and solid bars in black show segments of the chromosomes derived from Hwaseong and Oryza grandiglumis (O. gra) or O. rufipogon (O. rufi). Each number on top of the bars are chromosome number. (A) Hwaseong (Hwa). (B) tgw2-NIL. (C) gw9.1-NIL. (D) tgw2+gw9.1-NIL.
Chr.: chromosome, NIL: near isogenic line.
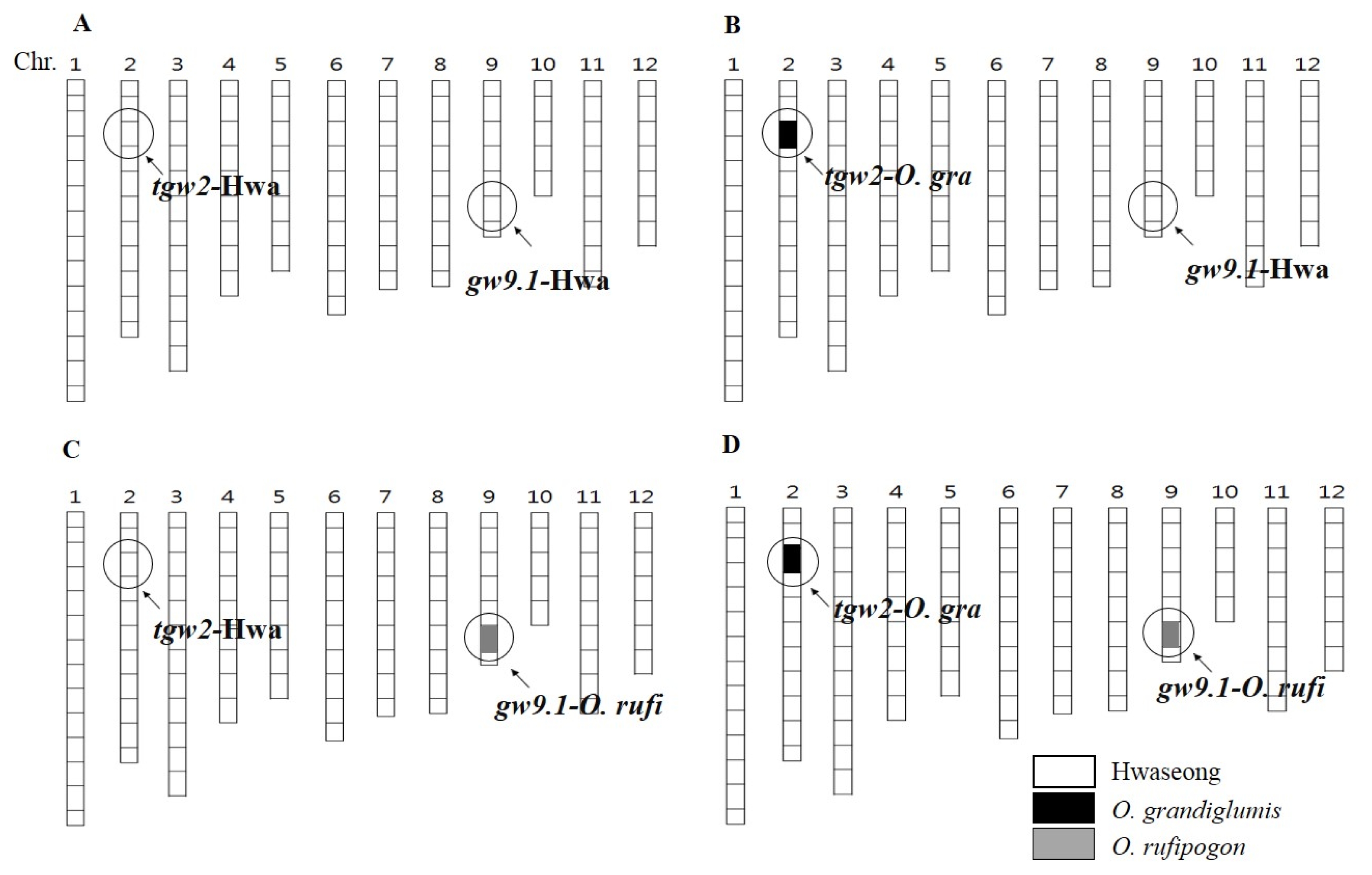
Fig. 2Frequency distribution of 1,000 grain weight in 3 genotype classes in an F2 population. NIL: near isogenic line, O.: Oryza.
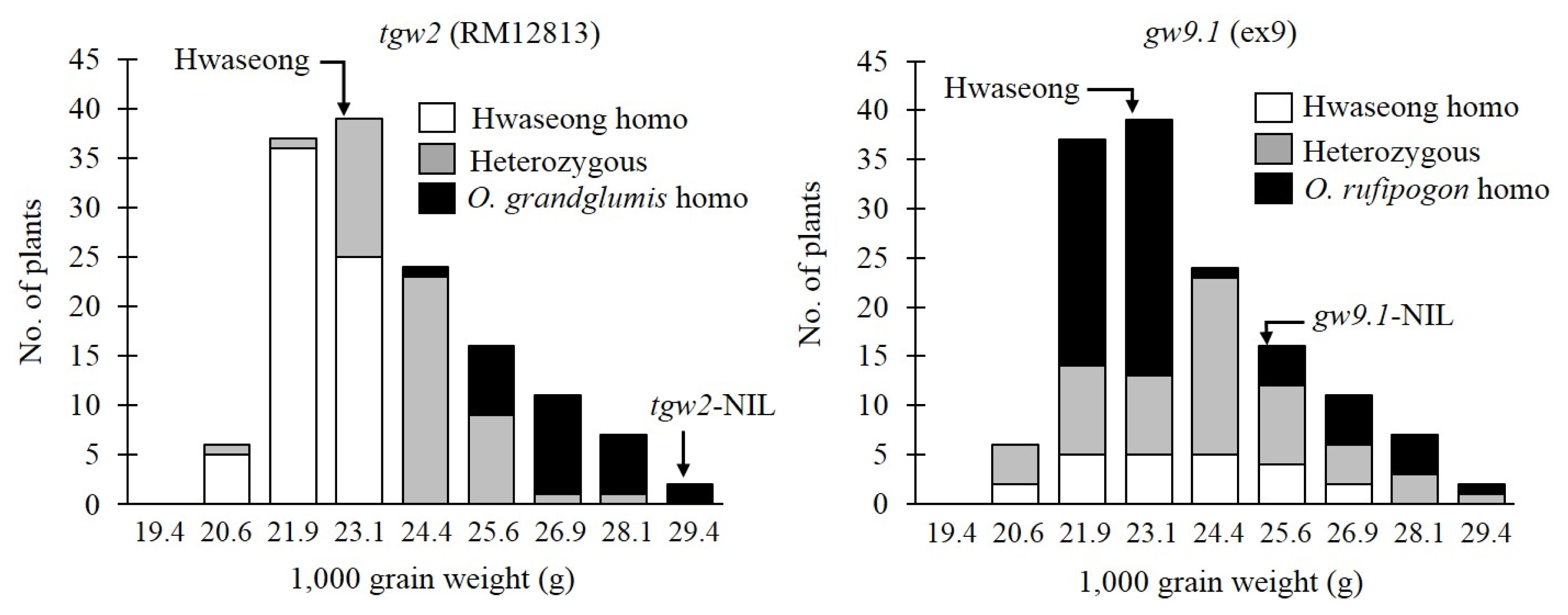
Fig. 3Differences in 1,000 grain weight for different genotype classes between tgw2 and gw9.1 in the F2 population from the cross between two quantitative trait loci (QTL)-near isogenic lines (NILs) tgw2-NIL and gw9.1-NIL. HH: homozygous for Hwaseong, GH/HR: heterozygous for Oryza grandiglumis or O. rufipogon, GG/RR: homozygous for O. grandiglumis or O. rufipogon.
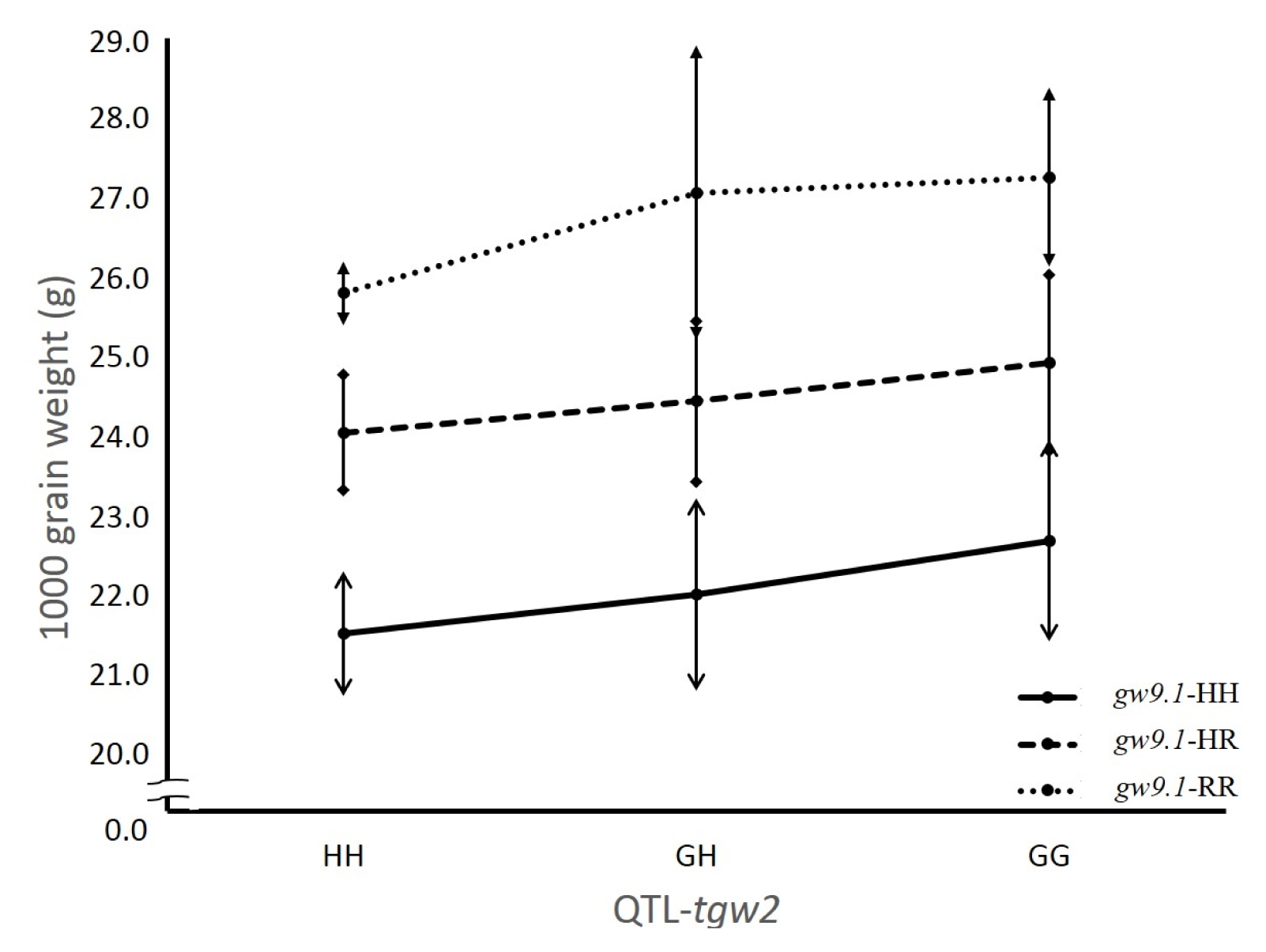
Fig. 4Comparison of the grain shape traits among 4 groups. (A) Grain length. (B) Grain width. (C) Grain thickness. a: Hwaseong, b: tgw2-near isogenic line (NIL), c: gw9.1-NIL, d: tgw2+gw9.1-NIL. White bar on picture indicates 10 mm.

Fig. 5Gene expression level of 3 different grain shape genes in Hwaseong (HS), tgw2-near isogenic line (NIL), and gw9.1-NIL. (A) Panicle samples for RNA extraction. White bar indicates 10 mm. Samples sizes are less than 1 cm. (B) GW2, (C) GS3, and (D) HGW. OsUBQ5 was used as normalization control for the RNA level. Error bars indicate standard deviation of three replicates. Each expression was repeated three times. One way ANOVA and Duncan’s multiple range test were done with each stage in GW2, GS3, and HGW at the significant level of P=0.05. Numbers followed by the same letter above the error bar are not significantly different at the P=0.05 based on Duncan’s multiple range test. a: HS, b: tgw2-NIL, c: gw9.1-NIL.
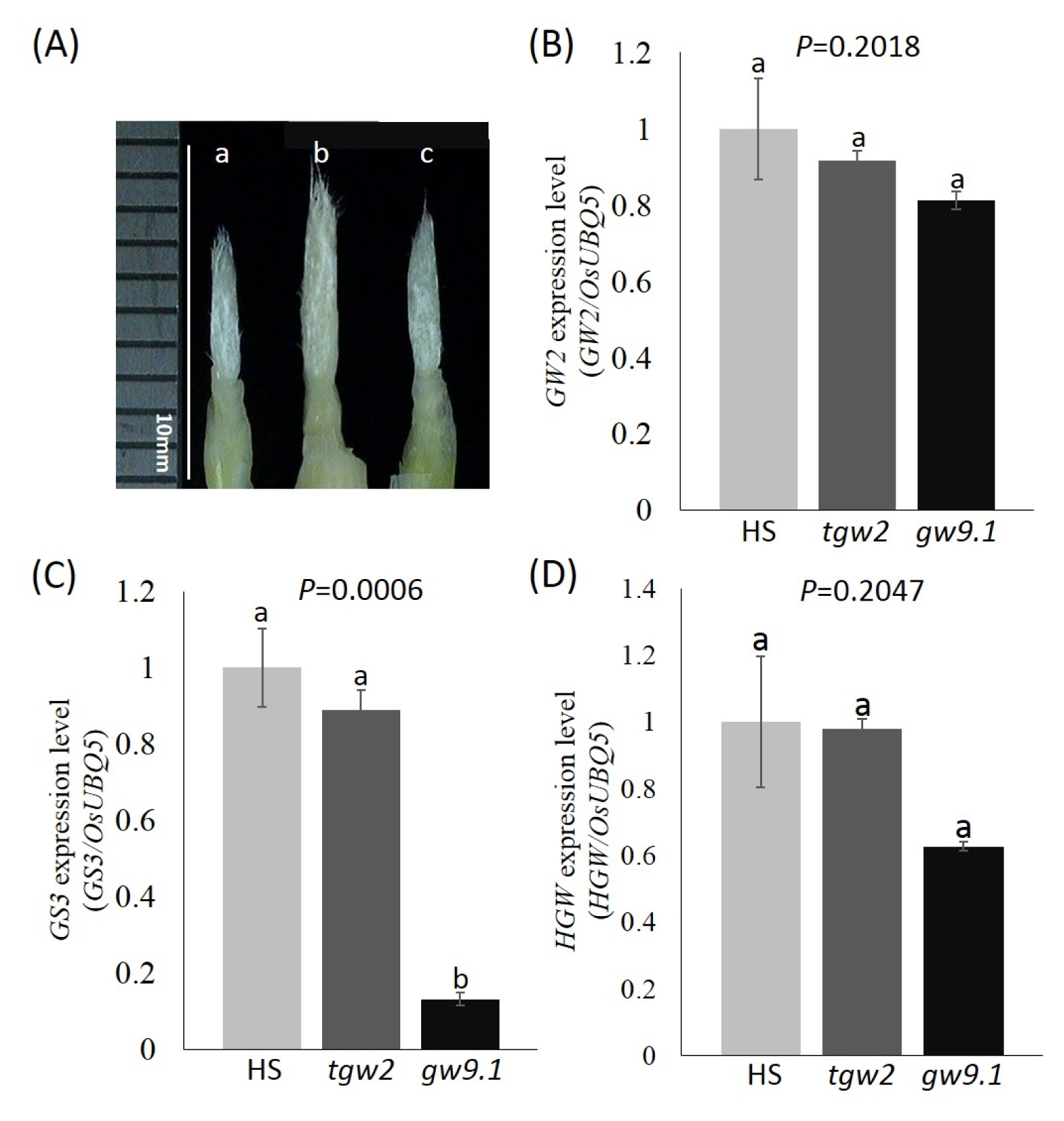
Table 1Primers for quantitative RT-polymerase chain reaction.
Table 1
|
Primerz)
|
Sequences (5′→3′)y)
|
Predicted size (bp) |
|
OsUBQ5
|
F |
GAAGTAAGGAAGGAGGAGGA |
100 |
|
R |
AAGGTGTTCAGTTCCAAGG |
|
qRT_GW2
|
F |
CTGCAGCAGGGAAGTGGT |
96 |
|
R |
ATGTCTGAGCCCGCGTAG |
|
qRT_GS3
|
F |
CATCGGAGAAGCGAAGTCA |
109 |
|
R |
CAGCAGCAGATCCAGGAGA |
|
HGW
|
F |
AGAACCCCAACTGGAGCAAG |
118 |
|
R |
ACTGGATCCCCTGGAGTAGC |
Table 2QTLs detected for grain weight based on one-way ANOVA in the F2 population.
Table 2
|
Cross |
QTLz)
|
Chr. |
Marker |
P-value |
R2
|
Mean |
|
|
HH |
GH/HR |
GG/RR |
|
tgw2/gw9.1
|
tgw2
|
2 |
RM12813 |
<0.0001 |
0.73 |
22.3by)
|
24.3b |
26.8a |
|
gw9.1
|
9 |
ex9 |
0.01 |
0.39 |
22.3b |
22.8ab |
23.9a |
Table 3Comparison of 11 traits among 4 different groups at F3 generation.
Table 3
|
Line |
Trait |
|
|
CLz)
|
PL |
PN |
SPP |
FER |
GL |
GW |
GT |
TGW |
CH |
AC |
|
Hwaseong |
92a |
19.3a |
13a |
102a |
92.0a |
5.29c |
2.87c |
2.17b |
23.0d |
0 |
20.1b |
|
tgw2-NIL |
90a |
17.8b |
11b |
100a |
84.3b |
5.37b |
3.18b |
2.35a |
29.6b |
4 |
24.1a |
|
gw9.1-NIL |
90a |
20.2a |
13a |
105a |
92.9a |
5.36b |
3.10b |
2.20b |
24.7c |
1 |
20.9b |
|
tgw2+gw9.1-NIL |
91a |
19.3a |
11b |
98a |
85.4b |
5.42a |
3.31a |
2.40a |
31.8a |
4 |
24.2a |
References
- Causse MA, Fulton TM, Cho YG, Ahn SN, Chunwongse J, Wu K, et al. 1994. Saturated molecular map of the rice genome based on an interspecific backcross population. Genetics. 138: 1251-1274.
- Fan C, Xing Y, Mao H, Lu T, Han B, Xu C, et al. 2006. GS3, a major QTL for grain length and weight and minor QTL for grain width and thickness in rice, encodes a putative transmembrane protein. Theor Appl Genet. 112: 1164-1171.
- Hu Z, He H, Zhang S, Sun F, Xin X, Wang W, et al. 2012. . motif-containing serine/threonine protein phosphatase determines the large grain QTL trait in rice. J Integr Plant Biol. 54: 979-990.
- Huang R, Jiang L, Zheng J, Wang T, Wang H, Huang Y, et al. 2013. Genetic bases of rice grain shape: so many genes, so little known. Trends Plant Sci. 18: 218-226.
- Ishimaru K, Hirotsu N, Madoka Y, Murakami N, Hara N, Onodera H, et al. 2013. Loss of function of the IAA-glucose hydrolase gene TGW6 enhances rice grain weight and increases yield. Nat Genet. 45: 707-711.
- Jin FX, Ji SD, Xie XB, Kang JW, Ju HG, Ahn SN. 2011. Detection of epistatic interaction of two QTLs, gw8.1 and gw9.1, underlying grain weight using nearly isogenic lines in rice. Jpn J Breed. 61: 69-75.
- Juliano BO. 1985. Rice : chemistry and technology. 2nd ed. St. Paul, Minn.: American association of cereal chemists.
- Kang JW, Yang P, Yun YT, Ahn SN. 2011. Mapping grain weight QTL Using near isogenic lines from an interspecific cross. Korean J Breed Sci. 43: 225-231.
- Li J, Chu H, Zhang Y, Mou T, Wu C, Zhang Q, et al. 2012. The rice HGW gene encodes a ubiquitin-associated (UBA) domain protein that regulates heading date and grain weight. PLoS One. 7: e34231
- Li Y, Fan C, Xing Y, Jiang Y, Luo L, Sun L, et al. 2011. Natural variation in GS5 plays an important role in regulating grain size and yield in rice. Nat Genet. 43: 1266-1269.
- Lu L, Shao D, Qiu X, Sun L, Yan W, Zhou X, et al. 2013. Natural variation and artificial selection in four genes determine grain shape in rice. New Phytol. 200: 1269-1280.
- Mao H, Sun S, Yao J, Wang C, Yu S, Xu C, et al. 2010. Linking differential domain functions of the GS3 protein to natural variation of grain size in rice. Proc Natl Acad Sci USA. 107: 19579-19584.
- Nagata K, Ando T, Nonoue Y, Mizubayashi T, Kitazawa N, Shomura A, et al. 2015. Advanced backcross QTL analysis reveals complicated genetic control of rice grain shape in a japonica x indica cross. Breed Sci. 65: 308-318.
- Panaud O, Chen X, McCouch SR. 1996. Development of microsatellite markers and characterization of simple sequence length polymorphism (SSLP) in rice (Oryza sativa L.). Mol Gen Genet. 252: 597-607.
- Shomura A, Izawa T, Ebana K, Ebitani T, Kanegae H, Konishi S, et al. 2008. Deletion in a gene associated with grain size increased yields during rice domestication. Nat Genet. 40: 1023-1028.
- Song XJ, Huang W, Shi M, Zhu MZ, Lin HX. 2007. . for rice grain width and weight encodes a previously unknown RING-type E3 ubiquitin ligase. Nat Genet. 39: 623-630.
- Song XJ, Kuroha T, Ayano M, Furuta T, Nagai K, Komeda N, et al. 2015. Rare allele of a previously unidentified histone H4 acetyltransferase enhances grain weight, yield, and plant biomass in rice. Proc Natl Acad Sci USA. 112: 76-81.
- Sun L, Li X, Fu Y, Zhu Z, Tan L, Liu F, et al. 2013. GS6, a member of the GRAS gene family, negatively regulates grain size in rice. J Integr Plant Biol. 55: 938-949.
- Takai T, Adachi S, Taguchi-Shiobara F, Sanoh-Arai Y, Iwasawa N, Yoshinaga S, et al. 2013. A natural variant of NAL1, selected in high-yield rice breeding programs, pleiotropically increases photosynthesis rate. Sci Rep. 3: 2149
- Wang S, Wu K, Yuan Q, Liu X, Liu Z, Lin X, et al. 2012. Control of grain size, shape and quality by OsSPL16 in rice. Nat Genet. 44: 950-954.
- Xie X, Jin F, Song MH, Suh JP, Hwang HG, Kim YG, et al. 2008. Fine mapping of a yield-enhancing QTL cluster associated with transgressive variation in an Oryza sativa x O. rufipogon cross. Theor Appl Genet. 116: 613-622.
- Yan S, Zou G, Li S, Wang H, Liu H, Zhai G, et al. 2011. Seed size is determined by the combinations of the genes controlling different seed characteristics in rice. Theor Appl Genet. 123: 1173-1181.
- Yano M, Sasaki T. 1997. Genetic and molecular dissection of quantitative traits in rice. Plant Mol Biol. 35: 145-153.
- Ying JZ, Gao JP, Shan JX, Zhu MZ, Shi M, Lin HX. 2012. Dissecting the genetic basis of extremely large grain shape in rice cultivar ‘JZ1560’. J Genet Genom. 39: 325-333.
- Yoon DB, Kang KH, Kim HJ, Ju HG, Kwon SJ, Suh JP, et al. 2006. Mapping quantitative trait loci for yield components and morphological traits in an advanced backcross population between Oryza grandiglumis and the O. sativa japonica cultivar Hwaseongbyeo. Theor Appl Genet. 112: 1052-1062.