Abstract
To introduce downy mildew resistance from a yellow-colored resistant cultivar, ‘Santero’, into a yellow breeding line, OT803, the F1 hybrid was produced by crossing Santero and OT803. The bulb color of the F1 hybrids became light pink, suggesting involvement of complementation between the DFR-A and ANS genes in the onion anthocyanin biosynthesis pathway. Since Santero contained active DFR-A and inactive ANS alleles, OT803 was assumed to harbor active ANS and inactive DFR-A alleles. However, some yellow-colored individuals of OT803 were shown to contain the homozygous genotype of the active DFR-AR4-like allele. Sequencing of 4,830-bp full-length sequences of this DFR-AR4-like allele revealed that the nucleotide sequences of the DFR-AR4 and DFR-AR4-like alleles were identical except for a single nucleotide deletion in the last exon. This single base-pair deletion resulted in creation of a premature stop codon at 2-bp downstream of the deletion mutation. This new DFR-A mutant allele was designated DFR-APS2. The RT-PCR results showed that transcripts of the DFR-APS2 allele were significantly reduced, suggesting involvement of nonsense-mediated mRNA decay (NMD) mechanism. The systematic process consisting of PCR amplification and sequencing of the PCR products was modified to identify the DFR-APS2 allele among 16 different DFR-A alleles. No additional accession was found to contain the DFR-APS2 allele from 155 diverse onion germplasm, indicating very limited distribution of this new DFR-APS2 allele.
-
Key words: Onion, Allium cepa, Bulb color, Dihydroflavonol 4-reductase (DFR), Molecular marker
INTRODUCTION
Anthocyanin, one of major flavonoid compounds, is responsible for the red bulb color, and quercetin derivatives are predominant in yellow onions (
Fossen et al. 1996;
Rhodes and Price 1996;
Slimestad et al. 2007). Inheritance studies (
Reiman 1931;
Clarke et al. 1944;
El-Shafie and Davis 1967) reported that five major loci were involved in determination of onion bulb colors. The
I locus, known as a color inhibiting factor, determines the white bulb color when its genotype is homozygous dominant. Meanwhile, the
C locus, known as a basic color factor, also determines the white bulb color only when its genotype is homozygous recessive. When the genotype of the
G locus is homozygous recessive, the chartreuse bulb color appears. The
R and
L loci are complementarily involved in determination of color difference between yellow and red bulb colors.
The
R and
L loci were revealed to encode dihydro-flavonol 4-reductase (DFR) and anthocyanidin synthase (ANS) enzymes, respectively, in the onion anthocyanin biosynthesis pathway (
Kim et al. 2004a,
2005a). One leaky and three inactive
ANS alleles were identified from diverse onion germplasm (
Kim et al. 2004b,
2005a,
2016). Recently, integration of a non-autonomous DNA trans-poson in the promoter region was revealed to cause significant reduction of the
ANS gene transcription (
Kim et al. 2015). Three homologous genes coding for the DFR enzyme were isolated in the previous study (
Kim et al. 2005b), and they were designated
DFR-A,
DFR-B, and
DFR-C. Among them, only the
DFR-A gene was active and determined the bulb color phenotypes (
Kim et al. 2005b). Since the first inactive
DFR-A allele was identified (
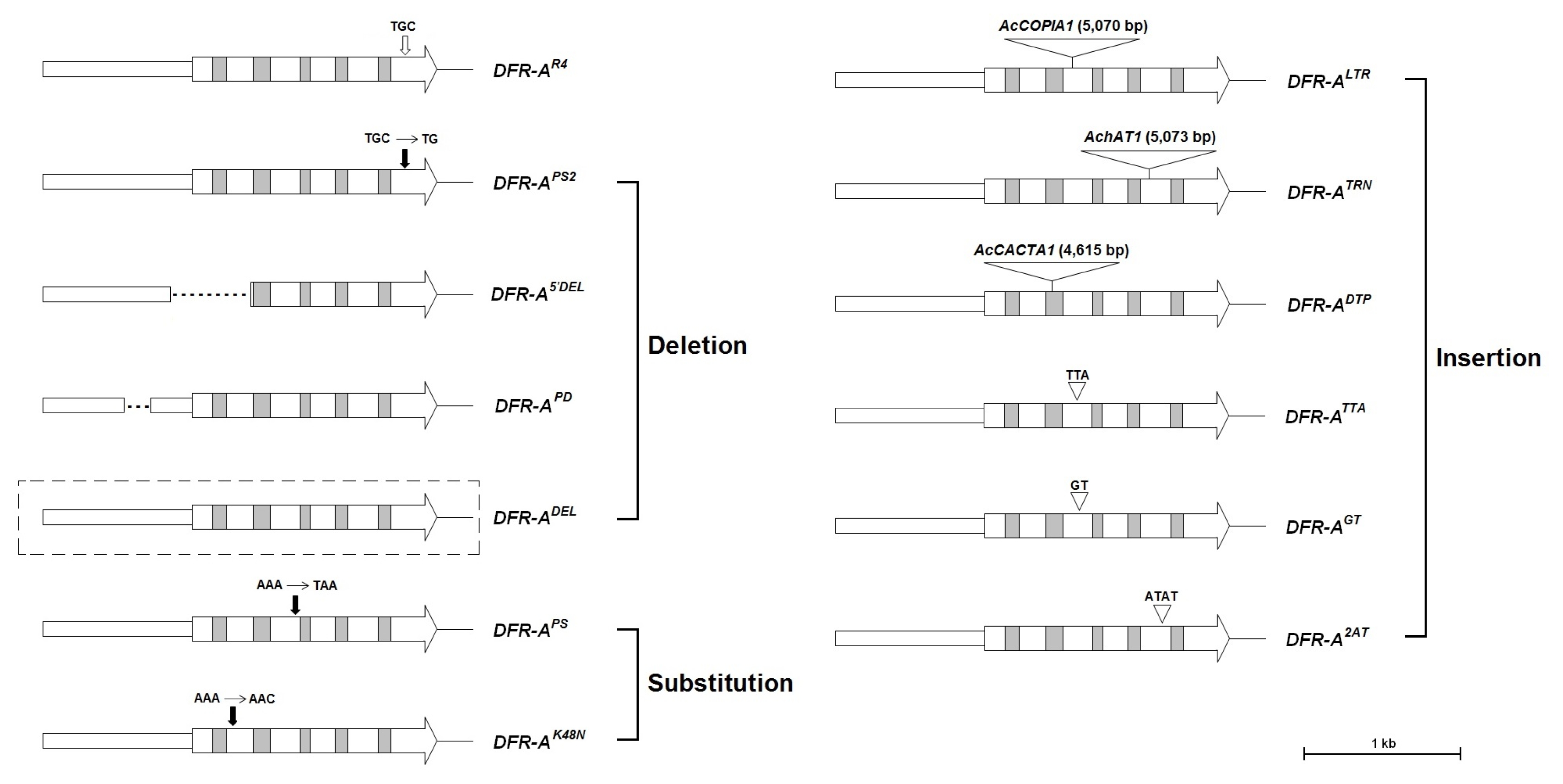
Kim et al. 2005b), eleven mutant alleles containing diverse insertion, deletion, or substitution mutations have been identified (
Kim et al. 2009,
2015;
Song et al. 2014). Three of them harbored transposable elements including a LTR retrotransposon and two different DNA transposons (
Kim et al. 2015).
In this study, a novel DFR-A allele containing a premature stop codon, which was created by one base-pair deletion, was identified from one breeding line. The inactivation mechanism of this mutant allele was proposed, and a systematic process for identification of 16 different DFR-A alleles was updated to include the newly identified DFR-A allele.
MATERIALS AND METHODS
Plant materials
A F
1 population originating from the cross between a downy mildew resistant cultivar, ‘Santero’, and a breeding line (OT803) was used to identify a new
DFR-A allele. To assess the frequency of the new
DFR-A allele, total genomic DNAs of 18 accessions containing the
DFR-AR4 allele were analyzed (
Supplementary Table 1). These 18 accessions were selected from 155 accessions collected from diverse countries in the previous study (
Song et al. 2014). Individual plants of OT803 which contained the homozygous genotype of the new mutant
DFR-A allele and a red-colored breeding line, H6, were used for RT-PCR amplification of the
DFR-A gene.
Total genomic DNAs were extracted from leaves of seedling or sprouted bulbs using a cetyl trimethy-lammonium bromide (CTAB) method (
Doyle and Doyle 1987). PCR was performed in 25-μL reaction mixtures containing 0.05 μg template, 2.5 μL 1× PCR buffer, 0.2 μL forward primer (10 μM), 0.2 μL reverse primer (10 μM), 0.2 μL dNTPs (10 mM each), and 0.25 μL polymerase mix (Advantage 2 Polymerase Mix; Clontech, Palo Alto, CA, U.S.A.). The primer sequences used in this study are listed in
Table 1.
PCR amplification was performed with an initial denaturation step at 94°C for 5 minutes, followed by 40 cycles of 94°C for 30 seconds, 65°C for 30 seconds, 72°C for 3 minutes, and a final 10 minutes extension at 72°C. The PCR products were visualized on a 1.5% agarose gel after ethidium bromide staining. For sequencing of PCR products, they were first purified using a QIAquick PCR Purification kit (QIAGEN, Valencia, CA, U.S.A.), and sequencing reactions were performed by a specialized company (Macrogen, Seoul, Republic of Korea). The full-length sequences of the new
DFR-A allele were obtained using the method described in the previous study (
Song et al. 2014).
Total RNAs were extracted from fresh leaf sheaths of four-leaf stage seedlings using a RNA extraction kit (RNeasy Plant Mini Kit, QIAGEN) following the manufacturer’s instructions. cDNAs were synthesized using a commercial cDNA synthesis kit (SuperScript™ III first-strand synthesis system for RT-PCR; Invitrogen, Carlsbad, CA, U.S.A.). RT-PCR amplification was performed with an initial denaturation step at 94°C for 3 minutes followed by 30 cycles of 94°C for 30 seconds, 65°C for 30 seconds, and 72°C for 3 minutes, and a final 10-minute extension at 72°C. The primer pair used in RT-PCR is listed in
Table 1. The onion tubulin sequence obtained from EST sequences (TC125) from the DFCI
Allium cepa Gene Index (
Antonescu et al. 2010) was used as a control.
RESULTS
Identification of a novel DFR-A mutant allele determining the yellow bulb color in onion
In order to introduce downy mildew resistance from a resistant cultivar, Santero, into a breeding line (OT803), the F
1 hybrids were produced by the cross between Santero and OT803. Although the bulb color of both parental lines were yellow, the bulb color of the F
1 hybrids became light pink (
Fig. 1). The light pink bulb color was assumed to appear by complementation between the
DFR-A and
ANS genes involved in the anthocyanin biosynthesis pathway as shown in the previous study (
Kim et al. 2005a). Since Santero contained an active
DFR-AR3 allele and inactive
ANSPS and
ANSS188L alleles (
Kim et al. 2016), OT803 was assumed to harbor an inactive
DFR-A allele and an active
ANS allele. As expected, ten individuals of OT803 contained all active
ANSh1 allele (
Table 2). Regarding the
DFR-A gene, the inactive
DFR-ATTA and the active
DFR-AR4-like alleles were identified from ten individuals, although the leaf sheath color of ten individuals were apparently yellow.
The 4,830-bp full-length sequences including putative promoter regions of the
DFR-AR4-like allele were obtained by sequencing of the PCR products. The nucleotide sequences of the
DFR-AR4-like allele were identical to those of the
DFR-AR4 allele except for a single base-pair change positioned at the 121th nucleotide of the last exon (
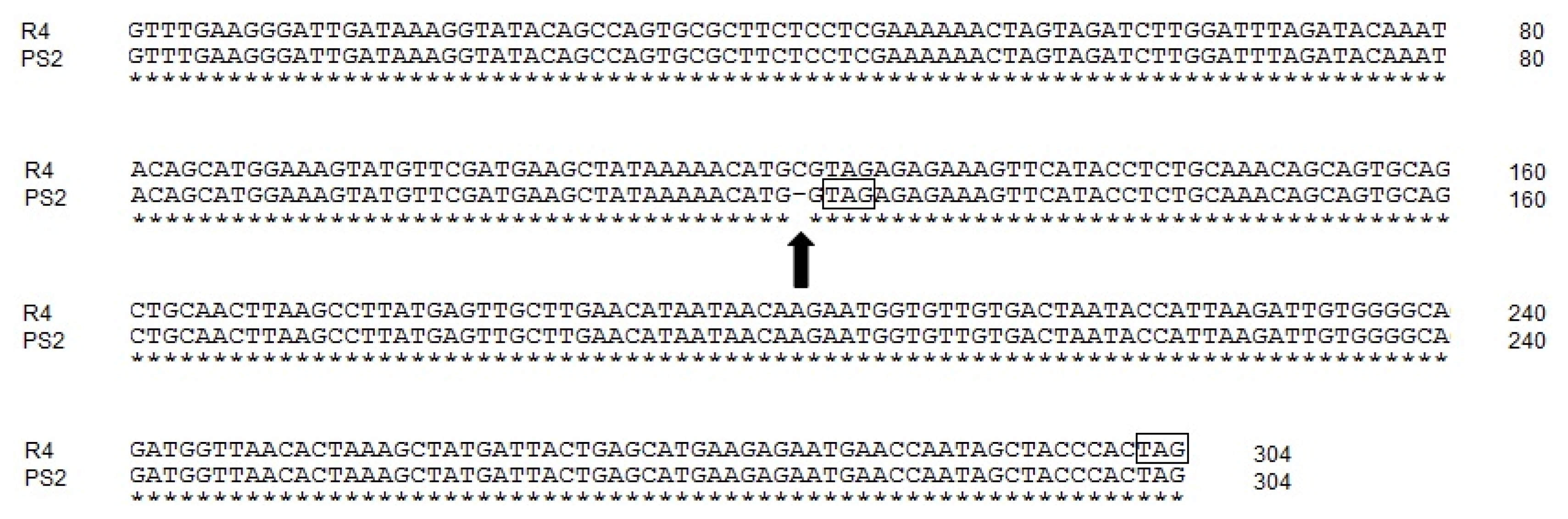
Fig. 2). The 121th cytosine nucleotide in the last exon was deleted in the
DFR-AR4-like allele. This single base-pair deletion created a premature stop codon at 2-bp downstream of the deletion mutation (
Fig. 3).
In the previous study (
Kim et al. 2009), transcripts of the
DFR-APS allele containing a premature stop codon in the exon3 (
Fig. 2) were shown to be significantly reduced. The nonsense-mediated mRNA decay (NMD) mechanism was proposed to cause degradation of the mRNA transcripts. To verify whether transcripts of the
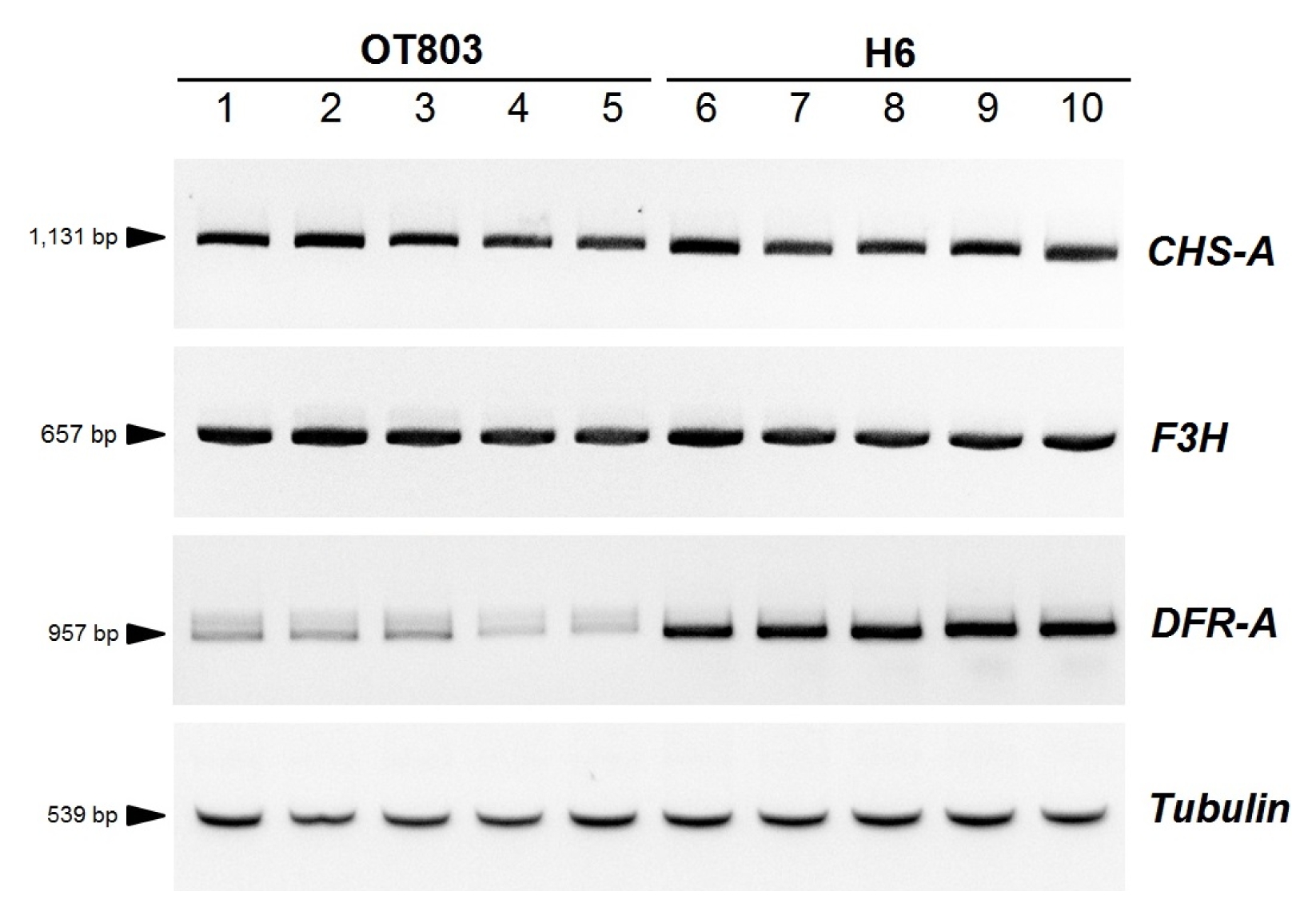
DFR-AR4-like allele were reduced by the same NMD mechanism, RT-PCR amplification was performed using five OT803 individuals harboring the homozygous
DFR-AR4-like allele. Results showed that the
DFR-A transcripts of all OT803 individuals were significantly reduced, and bands of the PCR products were smear, compared with the red breeding line ‘H6’ which contained the active
DFR-AR1 allele (
Fig. 4). However, transcription levels of other structural genes (
CHS-A and
F3H) involved in the onion anthocyanin biosynthesis pathway were not significantly different between OT803 and H6 individuals (
Fig. 4). Hereafter, this novel mutant allele is designated
DFR-APS2.
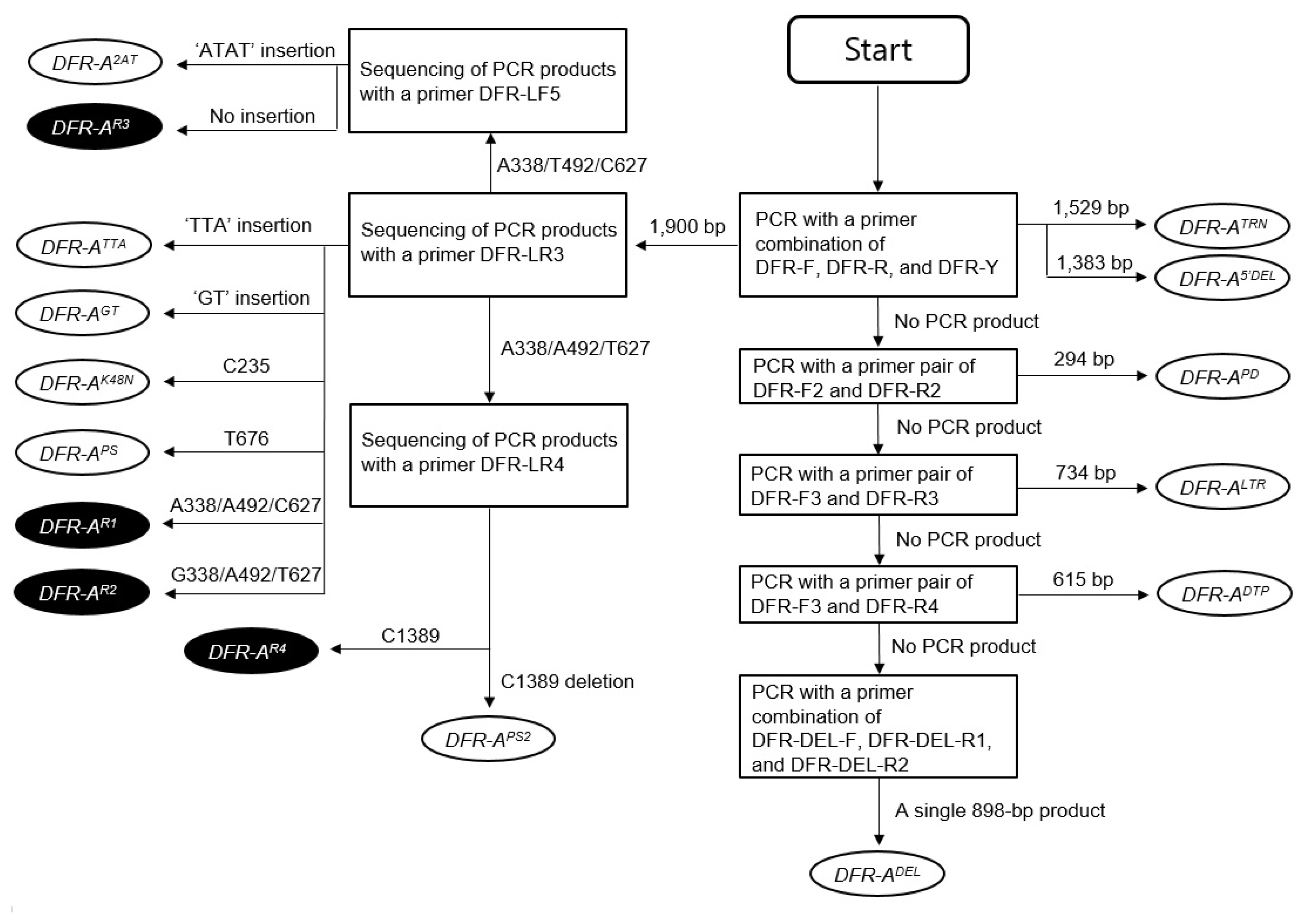
For identification of specific
DFR-A alleles from diverse onion germplasm, a systematic process consisting of PCR amplification and sequencing of the PCR products was devised for identification of 15 different
DFR-A alleles in the previous studies (
Song et al. 2014;
Kim et al. 2015). For identification of the
DFR-APS2 allele, one additional step of sequencing of the PCR product using the DFR-LR4 primer was added in the process to distinguish the
DFR-AR4 and
DFR-APS2 alleles (
Fig. 5).
In the previous study (
Song et al. 2014), the
DFR-A genotypes of 155 onion accessions originating from diverse countries were identified by using the process for identification of 15
DFR-A alleles. However, at that time, the
DFR-APS2 allele could not be distinguished from the
DFR-AR4 allele by such process since the position of the 1-bp deletion in the
DFR-APS2 allele could not be sequenced by the DFR-LR3 primer (
Fig. 5). Therefore, PCR products of 18 accessions containing the
DFR-AR4 allele were further sequenced by the DFR-LR4 primer following the updated process (
Fig. 5). However, sequencing results showed that no accession contained the
DFR-APS2 allele, indicating very limited distribution of the
DFR-APS2 allele in onion germplasm.
DISCUSSION
A novel inactive
DFR-A allele containing a premature stop codon in the last exon was identified in this study. This is the 12
th inactive
DFR-A allele identified from onion germplasm. In the previous studies (
Kim et al. 2005b,
2009,
2015;
Song et al. 2014), eleven inactive
DFR-A alleles containing diverse insertion, deletion, or substitution mutations were identified (
Fig. 2). Unlike other structural and regulatory genes involved in the onion anthocyanin biosynthesis pathway, the
DFR-A gene unusually has a large number of mutant alleles. No inactive mutant alleles of other structural and regulatory genes affecting color phenotypes have been identified in onions except for three
ANS (
Kim et al. 2005a,
2016) and one
CHI (
Kim et al. 2004c) mutant alleles. Although presence of multiple homologous
DFR genes in the onion genome is likely to contribute to creation of a multitude of
DFR-A mutants, the exact mechanism responsible for frequent mutation of the
DFR-A gene is not resolved yet.
Khar et al. (2008) showed that numerous copies of
DFR homologs existed in the onion genome by Southern blot analysis.
The
DFR-APS2 allele is probably derived from the
DFR-AR4 allele recently, since there is only a single nucleotide deletion between 4,830-bp promoter and coding regions of the
DFR-AR4 and
DFR-APS2 alleles. The single nucleotide deletion changes the reading frame and create a premature stop codon. However, loss of the last 60 residues, which is caused by the premature stop codon, may not have a significant effect on the function of the DFR enzyme, since the premature stop codon is positioned at the last exon. Indeed, none of the catalytic triad and 28 residues involved in binding of substrate and NADPH is positioned in the last exon (
Petit et al. 2007).
However, the RT-PCR result showed that the transcripts of the
DFR-APS2 allele were significantly reduced, and the PCR products appeared as smear bands compared with the normal
DFR-AR1 allele. Since there is no polymorphism in the 2.5-kb upstream region containing putative promoter regions between the
DFR-AR4 and
DFR-APS2 alleles, transcription of the
DFR-APS2 allele is assumed to be normal. Therefore, the NMD mechanism is most likely to be responsible for degradation of the
DFR-APS2 mRNA transcripts. The NMD mechanism, which is well conserved in all eukaryote, plays an important role in preventing deleterious effects of truncated proteins by degradation of defective mRNAs harboring premature stop codons (
Hentze and Kulozik 1999;
Neu-Yilik et al. 2004;
Chang et al. 2007;
Shaul 2015).
Although the causal genes responsible for the bulb color difference between red and yellow onions have been revealed as the genes coding for DFR and ANS enzymes (
Kim et al. 2004a,
2005a), designing of universal functional markers based on mutations is not a straightforward process due to a high level of allelic variation of the
DFR-A gene. In addition, complementation between the
DFR-A and
ANS genes makes it more complicated to identify specific
DFR-A alleles of yellow onion germ-plasm. Therefore, systematic processes are first required for identification of specific alleles of the
DFR-A and
ANS genes. Once specific
DFR-A or
ANS alleles of parental lines in segregating populations are determined, functional markers can be designed on the basis of polymorphism between two alleles. We updated the process for identifying 16
DFR-A alleles in this study. Combined with the process for
ANS allele identification (
Kim et al. 2016), the updated process for
DFR-A allele identification becomes an essential tool in onion breeding programs.
Supplementary Information
ACKNOWLEDGEMENTS
This research was supported by the Agriculture Research Center program, Golden Seed Project (Center for Horticultural Seed Development, No 213007-05-1-SBB10) and a grant from the Next-Generation BioGreen 21 Program (Plant Molecular Breeding Center No. PJ011034). The authors thank Ji-wha Hur, Jeong-Ahn Yoo, and Su-jung Kim for their dedicated technical assistance.
Fig. 1The onion bulb of the F1 hybrid between a yellow cultivar, Santero, and a yellow breeding line, OT803.

Fig. 2Organization of the inactive mutant DFR-A alleles isolated from onions. Arrow-shaped boxes indicate coding regions and the 5′-to-3′ direction. The empty and gray boxes in the coding regions indicate exons and introns, respectively. The vertical filled arrows on the exons indicate the positions of sequence changes. The nucleotide sequences on the vertical empty arrow indicate the normal sequences of the functional alleles. The dotted lines in the promoter region indicate the deleted sequences. The dotted line outside the DFR-ADEL allele indicate entire deletion of the genic region. Nucleotide sequences and names on the inverted triangles indicate inserted sequences and transposable elements, respectively. Lengths of transposable elements are shown in parenthesis.
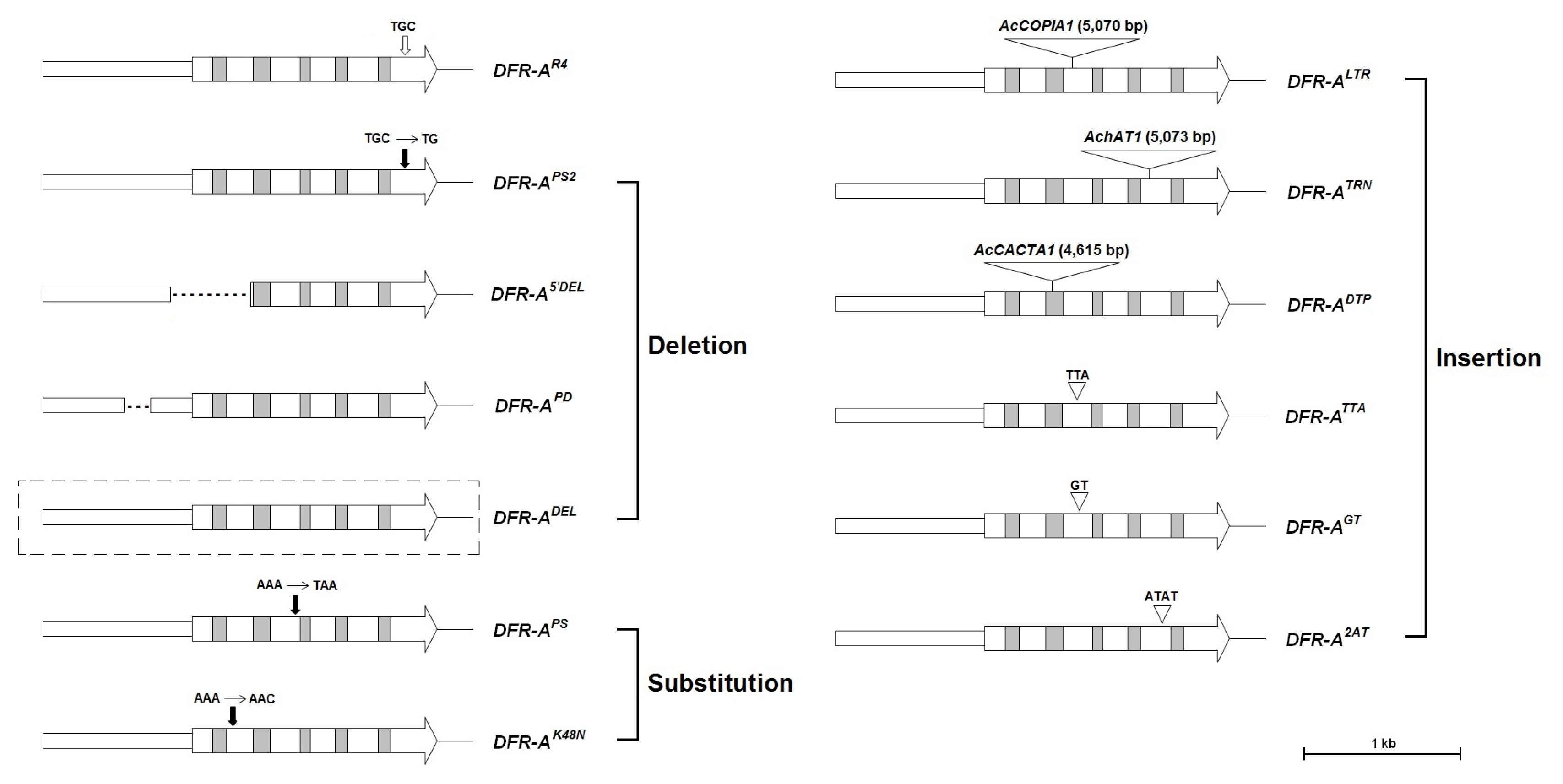
Fig. 3Alignment of partial nucleotide sequences of the DFR-AR4 and DFR-APS2 alleles. The vertical filled arrow indicates the positon of single base-pair deletion. The rectangular box indicates the position of the premature stop codon.
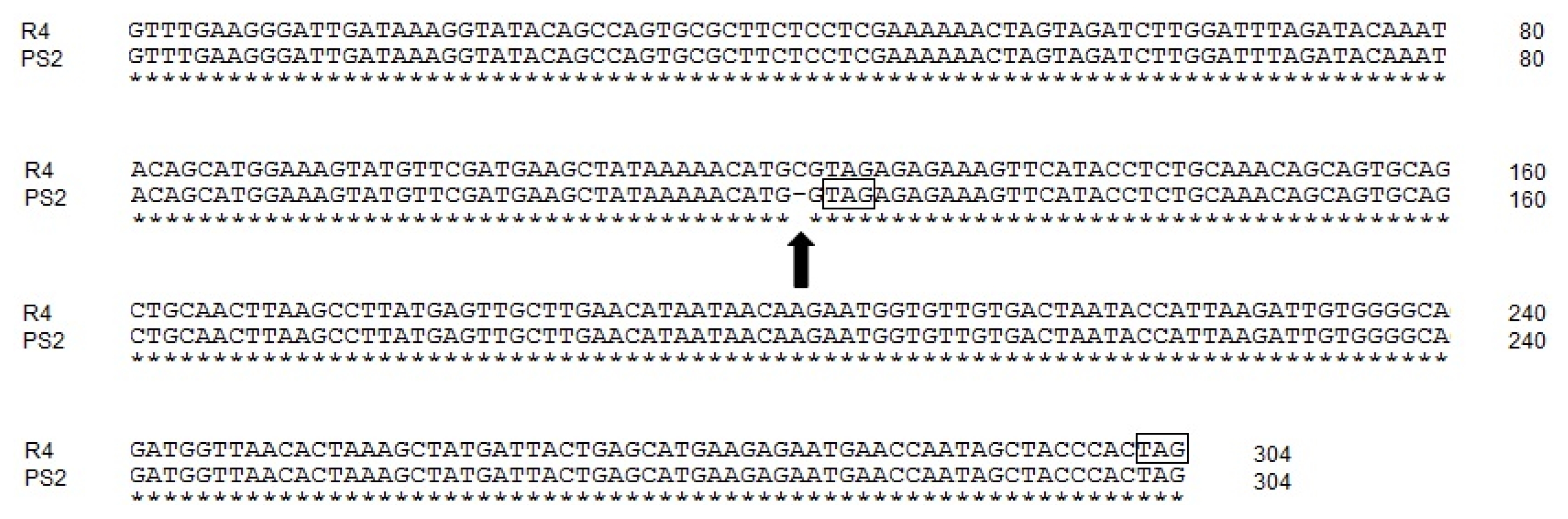
Fig. 4RT-PCR amplification of the DFR-A gene. 1–5: individuals of OT803 containing the homozygous DFR-APS2 allele. 6–10: individuals of H6 containing the homozygous DFR-AR1 allele. CHS-A: chalcone synthase-A, F3H: flavanone 3-hydroxylase
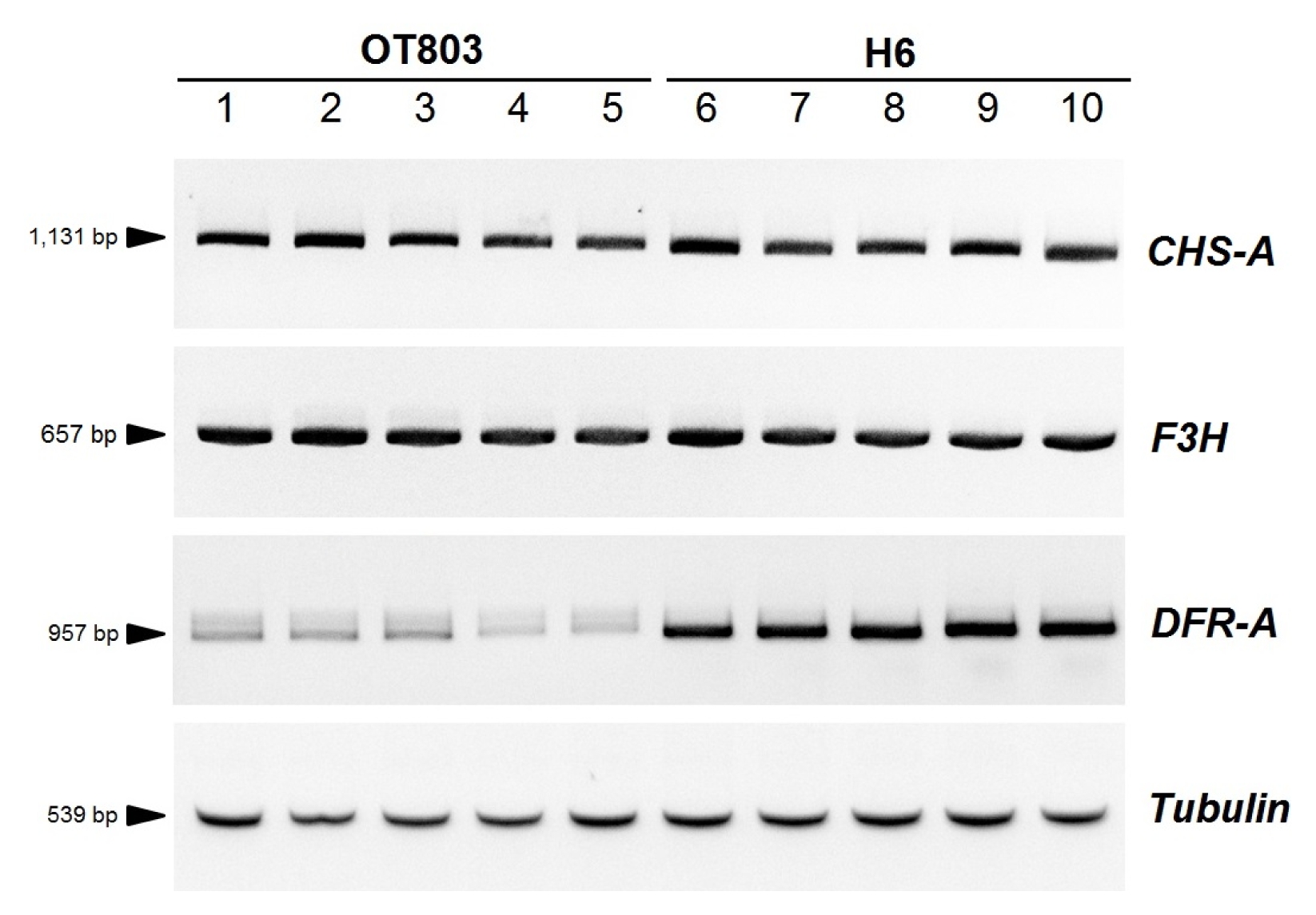
Fig. 5Flowchart showing the process for identification of 16 DFR-A alleles. The SNPs are shown with numbers indicating the distances from the start codon of the DFR-A gene. Four active and 12 inactive DFR-A alleles are shown in filled and empty oval-shaped circles, respectively.
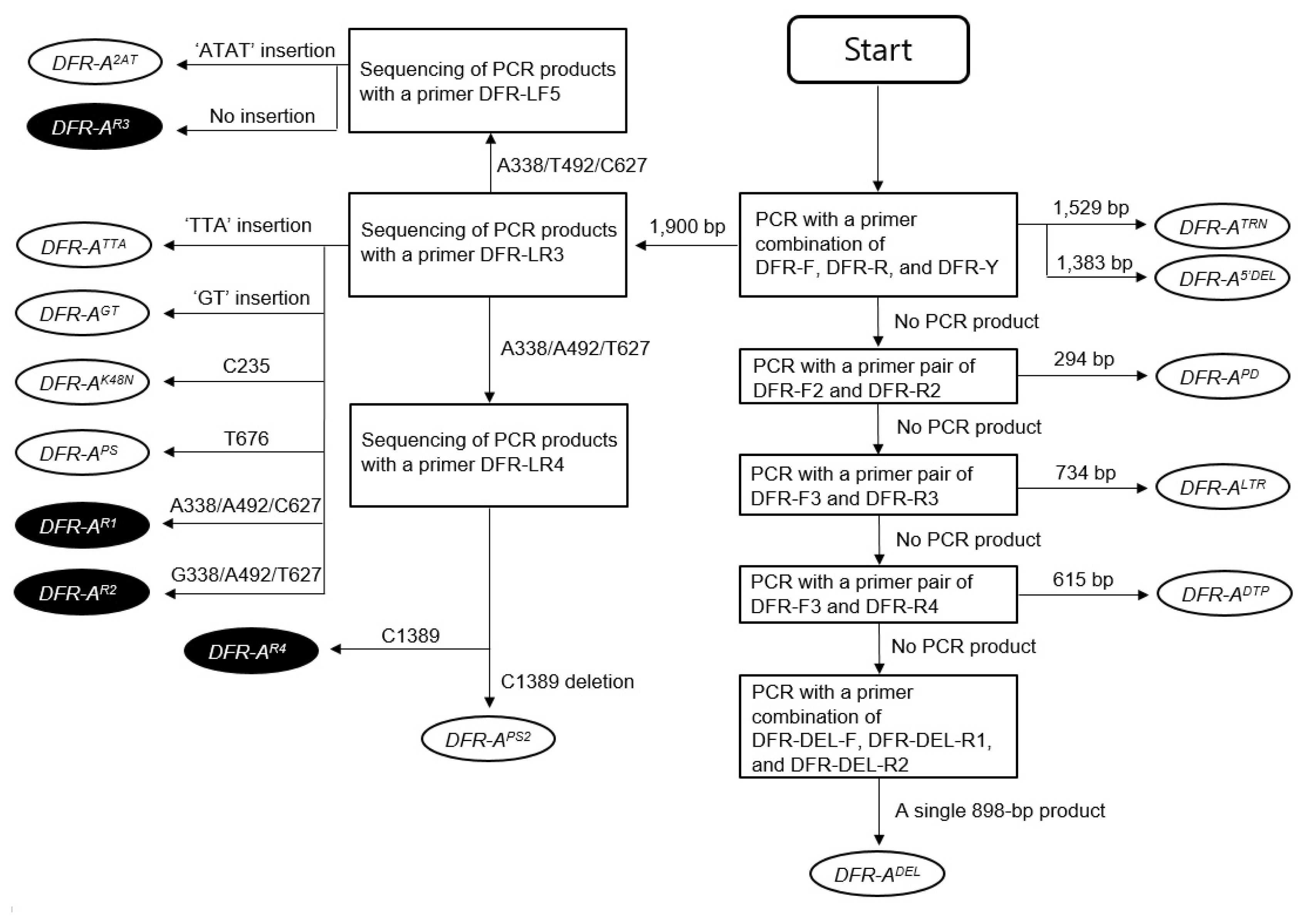
Table 1Primer sequences used in this study.
Table 1
|
Name |
Sequence (5′ to 3′) |
Application |
Reference |
|
DFR-F |
ATGCCAGTGGAGTGCATGTTGAATGGT |
DFR-A allele identification |
, Kim et al. (2005b)
|
|
DFR-R |
TGGGTAGCGATTGGTTCATTCTCTTCA |
DFR-A allele identification and RT-PCR |
, Kim et al. (2005b)
|
|
DFR-LR3 |
TTGCAAACTCCCATGCAGCTTTCTCTG |
DFR-A allele identification |
, Song et al. (2014)
|
|
DFR-LR4 |
TTAACCATCTGCCCCACAAT |
DFR-A allele identification |
This study |
|
DFR-LF5 |
GAGGAGCACCAAAAGCCCGAATACGAT |
RT-PCR |
, Song et al. (2014)
|
|
Tubulin-F |
CTGGGAGCTTTACTGCCTTG |
RT-PCR |
, Song et al. (2014)
|
|
Tubulin-R |
CAAGGGACCTCCTGCAAATA |
RT-PCR |
, Song et al. (2014)
|
|
CHS-F |
GAGGGTCCAGCCACGGTGTTAGCTG |
RT-PCR |
, Kim et al. (2004b)
|
|
CHS-R |
ATCAATGGCCACACTCCTAAGCACC |
RT-PCR |
, Kim et al. (2004b)
|
|
F3H-F |
GAAGACGAGCGTCCCAAGATTGCAT |
RT-PCR |
, Kim et al. (2004b)
|
|
F3H-R |
TCCACCATCTTTAGTCGCTTGCAGACC |
RT-PCR |
, Kim et al. (2004b)
|
Table 2Composition of DFR-A and ANS alleles of parental lines (Santero and OT803) and the F1 hybrid. The active DFR-A and ANS alleles are shown in boldfaces. The DFR-AR4-like allele is designated as DFR-APS2 allele.
Table 2
|
Population |
Genotype |
|
|
DFR-A
|
ANS
|
|
Santero |
DFR-AR3/DFR-AR3
|
ANSPS/ANSS188L
|
|
OT803 |
Mixed with homozygous and heterozygous DFR-APS2 and DFR-ATTA
|
ANSh1/ANSh1
|
|
F1 hybrid |
DFR-APS2/DFR-AR3
|
ANSh1/ANSS188L
|
References
- Antonescu C, Antonescu V, Sultana R, Quackenbush J. 2010. Using the DFCI gene index databases for biological discovery. Curr Protoc Bioinformatics. Unit1.6.1-Unit1.636.
- Brewster JL. 1994. Onions and other vegetable Alliums. CAB International. Wallingford, UK: pp. 1-26.
- Chang Y, Imam JS, Wilkinson MF. 2007. The nonsense-mediated decay RNA surveillance pathway. Annu Rev Biochem. 76: 51-74.
- Clarke AE, Jones HA, Little TM. 1944. Inheritance of bulb color in the onion. Genetics. 29: 569-575.
- Clere N, Faure S, Martinez MC, Andriantsitohaina R. 2011. Anticancer properties of flavonoids: roles in various stages of carcinogenesis. Cardiovasc Hematol Agents Med Chem. 9: 62-77.
- Cook NC, Samman S. 1996. Flavonoids-Chemistry, metabolism, cardioprotective effects, and dietary sources. Nutr Biochem. 7: 66-76.
- Cushnie TP, Lamb AJ. 2011. Recent advances in understanding the antibacterial properties of flavonoids. Int J Antimicrob Agents. 38: 99-107.
- Czemmel S, Heppel SC, Bogs J. 2012. R2R3 MYB transcription factors: key regulators of the flavonoid biosynthetic pathway in grapevine. Protoplasma. 249: S109-S118.
- Doyle JJ, Doyle JL. 1987. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem Bull. 19: 11-15.
- El-Shafie MW, Davis GN. 1967. Inheritance of bulb color in the onion (Allium cepa L.). Hilgardia. 38: 607-622.
- Fini A, Brunetti C, Di Ferdinando M, Ferrini F, Tattini M. 2011. Stress-induced flavonoid biosynthesis and the antioxidant machinery of plants. Plant Signal Behav. 6: 709-711.
- Fossen T, Andersen OM, Ovstedal DO, Pedersen AT, Raknes A. 1996. Characteristic anthocyanin pattern from onions and other Allium spp. J Food Sci. 61: 703-706.
- Goodrich J, Carpenter R, Coen ES. 1992. A common gene regulates pigmentation pattern in diverse plant species. Cell. 68: 955-964.
- Hentze MW, Kulozik AE. 1999. A perfect message: RNA surveillance and nonsense-mediated decay. Cell. 96: 307-310.
- Holton TA, Cornish EC. 1995. Genetics and biochemistry of anthocyanin biosynthesis. Plant Cell. 7: 1070-1083.
- Khar A, Jakse J, Havey MJ. 2008. Segregations for onion bulb colors reveal that red is controlled by at least three loci. J Amer Soc Hort Sci. 133: 42-47.
- Kim E, Kim C, Kim S. 2016. Identification of two novel mutant ANS alleles responsible for inactivation of anthocyanidin synthase and failure of anthocyanin production in onion (Allium cepa L.). Euphytica. 212: 427-437.
- Kim S, Baek D, Cho DY, Yoon M. 2009. Identification of two novel inactive DFR-A alleles responsible for failure to produce anthocyanin and development of a simple PCR-based molecular marker for bulb color selection in onion (Allium cepa L.) Theor. Appl Genet. 118: 1391-1399.
- Kim S, Binzel M, Yoo K, Park S, Pike LM. 2004a. Inactivation of DFR (Dihydroflavonol 4-reductase) gene transcription results in blockage of anthocyanin production in yellow onions (Allium cepa). Mol Breed. 14: 253-263.
- Kim S, Binzel ML, Yoo K, Park S, Pike LM. 2004b. Pink (P), a new locus responsible for a pink trait in onions (Allium cepa) resulting from natural mutations of anthocyanidin synthase. Mol Genet Genomics. 272: 18-27.
- Kim S, Jones R, Yoo K, Pike LM. 2004c. Gold color in onions (Allium cepa): a natural mutation of the chalcone isomerase gene resulting in a pre-mature termination codon. Mol Genet Genomics. 272: 411-419.
- Kim S, Jones R, Yoo K, Pike LM. 2005a. The L locus, one of complementary genes required for anthocyanin production in onions (Allium cepa), encodes anthocyanidin synthase. Theor Appl Genet. 111: 120-127.
- Kim S, Park JY, Yang T. 2015. Characterization of three active transposable elements recently inserted in three independent DFR-A alleles and one high-copy DNA transposon isolated from the Pink allele of the ANS gene in onion (Allium cepa L.). Mol Genet Genomics. 290: 1027-1037.
- Kim S, Yoo K, Pike LM. 2005b. Development of a PCR-based marker utilizing a deletion mutation in the DFR (dihydroflavonol 4-reductase) gene responsible for the lack of anthocyanin production in yellow onions (Allium cepa). Theor Appl Genet. 110: 588-595.
- Lotito SB, Frei B. 2006. Comsumption of flavonoids-rich foods and increased plasma antioxidant capacity in humans: cause, consequence, or epiphenomenon? Free Radic Biol Med. 41: 1727-1746.
- Neu-Yilik G, Gehring NH, Hentze MW, Kulozik AE. 2004. Nonsense-mediated mRNA decay: from vacuum cleaner to Swiss army knife. Genome Biol. 5: 218
- Nishiumi S, Miyamoto S, Kawabata K, Ohnishi K, Mukai R, Murakami A, et al. 2011. Dietary flavonoids as cancer-preventive and therapeutic biofactors. Front Biosci. 3: 1332-1362.
- Petit P, Granier T, d’Estaintot BL, Manigand C, Bathany K, Schmitter J, et al. 2007. Crystal structure of grape dihydroflavonol 4-reductase, a key enzyme in flavonoid biosynthesis. J Mol Biol. 368: 1345-1357.
- Petroni K, Tonelli C. 2011. Recent advances on the regulation of anthocyanin synthesis in reproductive organs. Plant Sci. 181: 219-229.
- Quattrocchio F, Wing JF, Leppen HTC, Mol JN, Koes RE. 1993. Regulatory genes controlling anthocyanin pigmentation are functionally conserved among plant species and have distinct sets of target genes. Plant Cell. 5: 1497-1512.
- Ramsay NA, Glover BJ. 2005. MYB-bHLH-WD40 protein complex and the evolution of cellular diversity. Trends Plant Sci. 10: 63-70.
- Reiman GH. 1931. Genetic factors for pigmentation in the onion and their relation to disease resistance. J Agr Res. 42: 251-278.
- Rhodes MJC, Price KR. 1996. Analytical problems in the study of flavonoid compounds in onions. Food Chem. 57: 113-117.
- Russo M, Spagnuolo C, Tedesco I, Bilotto S, Russo GL. 2012. The flavonoid quercetin in disease prevention and therapy: Facts and fancies. Biochem Pharmacol. 83: 6-15.
- Shaul O. 2015. Unique aspects of plant nonsense-mediated mRNA decay. Trends Plant Sci. 20: 767-779.
- Shirley BW. 1996. Flavonoid biosynthesis: ‘new’ functions for an ‘old’ pathway. Trends Plant Sci. 1: 377-382.
- Slimestad R, Fossen T, Vågen IM. 2007. Onions: a source of unique dietary flavonoids. J Agric Food Chem. 55: 10067-10080.
- Song S, Kim C, Moon JS, Kim S. 2014. At least nine independent natural mutations of the DFR-A gene are responsible for appearance of yellow onions (Allium cepa L.) from red progenitors. Mol Breed. 33: 173-186.
- Spelt C, Quattrocchio F, Mol JN, Koes RE. 2000. antho-cyanin1 of Petunia encodes a basic helix-loop-helix protein that directly activates transcription of structural anthocyanin genes. Plant Cell. 12: 1619-1631.
- Veitch NC, Grayer RJ. 2011. Flavonoids and their glycosides, including anthocyanins. Nat Prod Rep. 28: 1626-1695.
- Yamazaki M, Makita Y, Springob K, Saito K. 2003. Regulatory mechanisms for anthocyanin biosynthesis in chemotypes of Perilla frutescens var. crispa. Biochem Eng J. 14: 191-197.