Abstract
Pyropia tenera is an intertidal red alga of commercial significance owing to its popularity as a health-promoting seafood product. This alga grows in marine environments and is frequently exposed to high salinity and osmotic stress, which impact its growth. Therefore, the enhancement of stress tolerance in P. tenera is critical. In the present work, we aimed to elucidate the mechanisms underlying abiotic stress tolerance in this species; specifically, we identified the P. tenera S-adenosylmethionine synthetase-encoding gene (PtSAMS) and characterized its biological function. This gene, which is known to play a role in stress tolerance in other plants, was cloned and overexpressed in Escherichia coli under high-salinity conditions. The PtSAMS gene was found to encode a 385-amino-acid protein with a molecular weight of 41.8 kDa. In silico sequence alignment and phylogenetic analysis of the PtSAMS amino acid sequence showed that the encoded protein comprises three conserved domains and two motifs that are highly conserved in other plants. Growth assay results indicated that PtSAMS-overexpressing E. coli cells exhibit enhanced tolerance to salt stress. The results suggest that PtSAMS expression is induced by a combination of ion toxicity and osmotic stress resulting from exposure to high salinity in marine environments, and that this gene is expressed at housekeeping levels owing to growth in such conditions. The findings suggest that PtSAMS could be used as a potentially valuable bioresource with utility in the genetic engineering of salt stress-tolerant crop plants.
-
Key words: S-adenosylmethionine synthetase (SAMS), Pyropia tenera, Abiotic stress, Salinity, Recombinant protein, Tolerance
INTRODUCTION
Algae have been consumed as edible seaweed in Asia for thousands of years.
Pyropia (Rhodophyta) have been cultivated since the 17
th century in Korea, Japan, and China and represent commercially valuable seafood.
Pyropia, which is highly popular owing to its delicious taste, contains high levels of protein and is rich in essential amino acids and vitamins (
Blouin et al. 2011). The market for
Pyropia is worth about 7 trillion, and its exports have experienced recent growth. It expected that the
Pyropia market size would continue to increase as a result of its recognition as a health-beneficial food product.
The environment in which seaweed is cultivated is exposed to harsh marine conditions. Aquaculture productivity is affected by temperature, rainfall, laver disease, and other factors. Therefore, the identification of rapidly growing or disease-resistant seaweed species is critical.
Salinity is one of the major abiotic factors that limit crop growth and productivity. Salt stress results in excessive ion toxicity, oxidative stress, and physiological drought, consequently causing disruption of cell organelles and their metabolism, eliciting nutrient imbalance in the plant, and reducing its osmotic potential (
Tester and Bacic 2005). These effects lead to arrest of growth and development of the plant, thereby limits its survival.
S-Adenosylmethionine synthetase (SAMS) catalyzes the conversion of adenosine 5′-triphosphate (ATP) and methionine into S-adenosylmethionine (SAM) (
Pajares and Markham 2011). SAM plays a central role in diverse biological processes, such as by serving as a methyl donor during transmethylation in plants, and acting as a common precursor in the biosynthesis of polyamines and ethylene (
Kumar et al. 1997;
Roeder et al. 2009). The SAMS-encoding gene has been cloned from various species such as bacteria, yeasts, humans, animals, and plants. In plants, SAMS has been reported to play a role in developmental process and the stress response (
Pulla et al. 2009). In
Arabidopsis thaliana, the
SAMS gene plays a role in seed germination, as shown in a previous study of MTO3 mutants that showed delayed germination and high level concentration of methionine phenotype Knockdown of the
SAMS gene resulted in delayed flowering time and dwarf phenotype in rice (
Boerjan et al. 1994;
Li et al. 2011). Expression of
SAMS is induced under low temperature in
Arabidopsis, rice, and maize (
Cui et al. 2005;
Amme et al. 2006;
Yang et al. 2006;
Uvackova et al. 2012).
Pisum sativum SAMS1 is expressed during pea development (
Gómez-Gómez and Carrasco 1998). Overexpression of the stress-inducible gene
GsSAMS2 enhances salt tolerance in transgenic
Medicago sativa (
Hua et al. 2012).
Suaeda salsa inhabits saline or alkaline soil such as coastal salt-flats. Recently, overexpression of
S. salsa SAMS was shown to increase salt tolerance in transgenic tobacco (
Qi et al. 2010). Although
Porphyra yezoensis SAMS (PySAMS) has been cloned (
Yi et al. 2009), its function is still unknown.
Here, we reported the cloning and characterization of the SAMS (PtSAMS) gene in Pyropia tenera. In order to elucidate the function of PtSAMS, we additionally examined the expression of PtSAMS in E. coli under stress conditions. Functional analysis in E. coli suggested that PtSAMS plays a role in the tolerance to stress.
MATERIALS AND METHODS
Plant materials
The P. tenera strain used in this study was received from the Seaweed Research Center, National Research & Development Institute. P. tenera was cultured at 10°C in a growth chamber with bubbling under 50 μmol·m−2s−1 and a photoperiod of 12 hour light and 12 hour dark. The culture media used was Provasoli’s enrichment solution (PES); the media were changed every week (Kakinuma et al. 2016).
RNA extraction, reverse transcription, and isolation of PtSAMS gene
The
PtSAMS sequence was obtained by performing a search of the NCBI (National Center of Biotechnology Information) database, based on the sequence of
Porphyra yezoensis S-adenosylmethionine synthetase (
SAMS) gene. Total RNA was extracted from
P. tenera with Hybrid-R (GeneAll, Seoul, Korea) according to the manufacturer’s guide. The first-strand cDNA was synthesized using amfiRivert cDNA Synthesis Platinum Master Mix (GenDEPOT, Texas, US). The full-length
PtSAMS gene was isolated through RT-PCR using the
PtSAMS-F (5′-CACCATGGCAGCCATGAAG-3′) and
PtSAMS-R (5′-ACGACGCTCTAGAGCTCAC-3′) primers based on the
P. yezoenesis sequence (GenBank accession: FJ404748) (
Table 1).
PtSAMS was amplified by PCR using the
PtSAMS-NdeI-F (5′-CCCCATATGGCAGCCATGAAGA-3′) and
PtSAMS-
XhoI-R (5′-TCACTCGAGGAGCTCAAGC-3′) primers with flanking restriction sites of
NdeI and
XhoI, respectively (
Table 1). The PCR product was digested with
NdeI and
XhoI restriction endonucleases and cloned into pET28(b). The pET28(b)-
PtSAMS plasmid was transformed into the
E. coli strain BL21(DE3). The pET28(b) empty vector was used as a control in spot assay and liquid growth assay of stress treatment (
Yadav et al. 2012).
Sequence analysis
PtSAMS and protein sequences of other SAMS of different species were searched in the NCBI database. Amino acid alignments were conducted using the ClustalW algorithm implemented in BioEdit (
Thompson et al. 1994). The phylogenetic tree of the SAMS protein was constructed using the Geneious software 9.0.4. using a neighbor-joining (NJ) method with bootstrap set at 10,000 replicates.
Genomic DNA was isolated from
P. tenera using the DNeasy Plant Maxi-prep Kit (Qiagen, Hilden, Germany). Genomic DNA (5 μg) was digested with
BamHI and
KpnI, separated on 0.7% agarose gel, and transferred to Hybond-N+ membrane. The membrane was treated by UV crosslinking to fix the genomic DNA. Probe DNA was prepared by PCR for a specific region of
PtSAMS. The gene-specific primers used were
PtSAMS-303-F (5′-CCAGTCCCCTGAGATTGCTG-3′) and
PtSAMS-955-R (5′-CTGAAATCGGCTCGGCAATG-3′) (
Table 1). Probe labeling with dioxigenin (DIG), hybridization, and detection were carried out using the DIG High Prime DNA Labeling and Detection Starter Kit I (Roche, Basel, Switzerland) according to the manufacturer’s instructions.
cDNA from
P. tenera was used as a template for qRT-PCR using
PtSAMS gene-specific primers
PtSAMS-303-F and
PtSAMS-955-R. Real time qPCR was performed using the Takara PCR system with a SYBER GREEN KIT (Takara, Shiga, Japan). PCR conditions were as follows: 94°C for 2 minutes, 40 cycles at 94°C for 30 seconds, 62°C for 30 seconds, 72°C for 30 seconds, and a final extension at 72°C for 10 minutes. The experiments were repeated three times independently. The
PtGAPDH gene was used as an internal control under the same conditions. Primer sequences were as follows:
PyGAPDH-2771-F (5′-CGCC GAGTACATTGTCGAGT-3′) and
PyGAPDH-3002-R (5′-GTACTTCTCGTGCAGCACCT-3′) (
Lee et al. 2015) (
Table 1).
pET28(b)-
PtSAMS/BL21(DE3) cells were grown in LB medium to OD
600 = 0.6, and 0.1 mM IPTG was added (
Yadav et al. 2012). The inducted cells were grown for 5 hours at 37°C. The cultured cells were dil0uted to an OD
600 of 0.6, and then to 10
−3, 10
−4, and 10
−5. Ten micro liters from each dilution was spotted on to an LB-only plate, or an LB plate supplemented with 400 mM NaCl and 400 mM KCl. The experiments were repeated three times independently.
Growth rate analysis was tested in LB liquid medium or in LB medium supplemented with NaCl and KCl. Then, 400 μL of pET28(b)-PtSMAS recombinant or vector alone in E. coli BL21 cells were inoculated into 10 mL of LB liquid medium for overnight culture. The next day, the cultured cells were diluted to an OD600 of 0.6 and 0.1 mM IPTG was added for induction. Cells were grown for 5 hours at 37°C and then were diluted to OD600 of 0.6. Next, 500 μL of cells were inoculated into 50 mL of LB medium containing 400 mM NaCl, and 400 mM KCl, or LB medium only, and cultured at 37°C. Cells were harvested every 1 hour, until 12 hours, and OD600 values were measured.
RESULTS
Isolation and characterization of PtSAMS
To isolate the
PtSAMS, we designed primer sets based on the
P. yezoensis sequence (Accession number: FJ404748) in the NCBI GenBank sequence database (
Table 1). The
PtSAMS gene was cloned from the cDNA library by PCR and sequenced (data not shown). Sequence analysis showed that the complete open reading frame of
PtSAMS is 1,155 bp in length, and is composed of one exon. The predicted PtSAMS protein comprises 385 amino acid residues with a calculated molecular weight of 41.8 kDa and isoelectric point (PI) of 5.60.
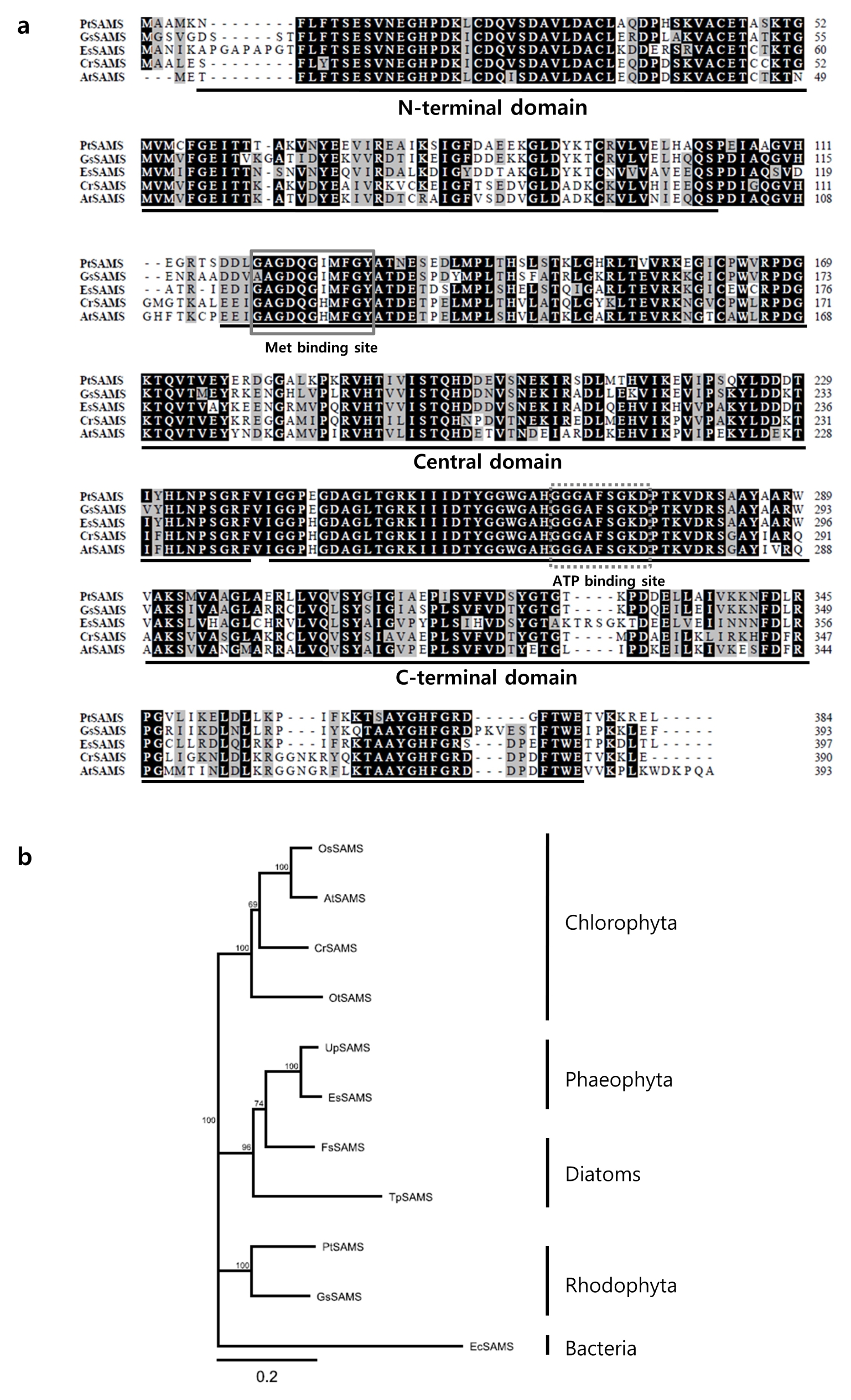
To further characterize the PtSAMS protein, we performed a conserved domain (CD) analysis through the GenomeNet Database Resources (
http://www.genome.jp/tools/motif/). The predicted PtSAMS protein consisted of three S-adenosylmethionine synthetase N-terminal, central, and C-terminal domains. Furthermore, the predicted PtSAMS sequence contained two substrate-binding sites, as well as a site for ATP binding in the central domain and for Met binding in the C-terminal domain (
Fig. 1a). The BioEdit software was used for comparison of the amino acid sequence of PtSAMS with SAMS from other representative Rhodophyta, Phaeophyta, and Chlorophyta (
Fig. 1a). The results showed that PtSAMS shared higher homology with SAMS from plant species such as
Galdieria sulphuraria (76%),
Ectocarpus siliculosus (70%),
Chlamydomonas reinhardtii (69%), and
A. thaliana (66%). To investigate the evolutionary relationship of PtSAMS protein among SAMS proteins of other species, a phylogenetic tree was constructed using Geneious 9.0.4 software on the basis of the multiple amino acid sequences (
Fig. 1b). GenBank accession numbers of the protein sequences are shown in
Table 2. Phylogenetic tree analysis revealed that the PtSAMS of Bangiophyceae was grouped with red algae, showing the highest degree of clustering with Rhodophyta,
Galdieria sulphuraria, and then with SAMS of diatoms and Phaeophyta, followed by Chlorophyta.
Table 3 shows pairwise comparison of the amino acid sequences between PtSMAS and other species. The SAMS sequence was found to be highly conserved among all the different species.
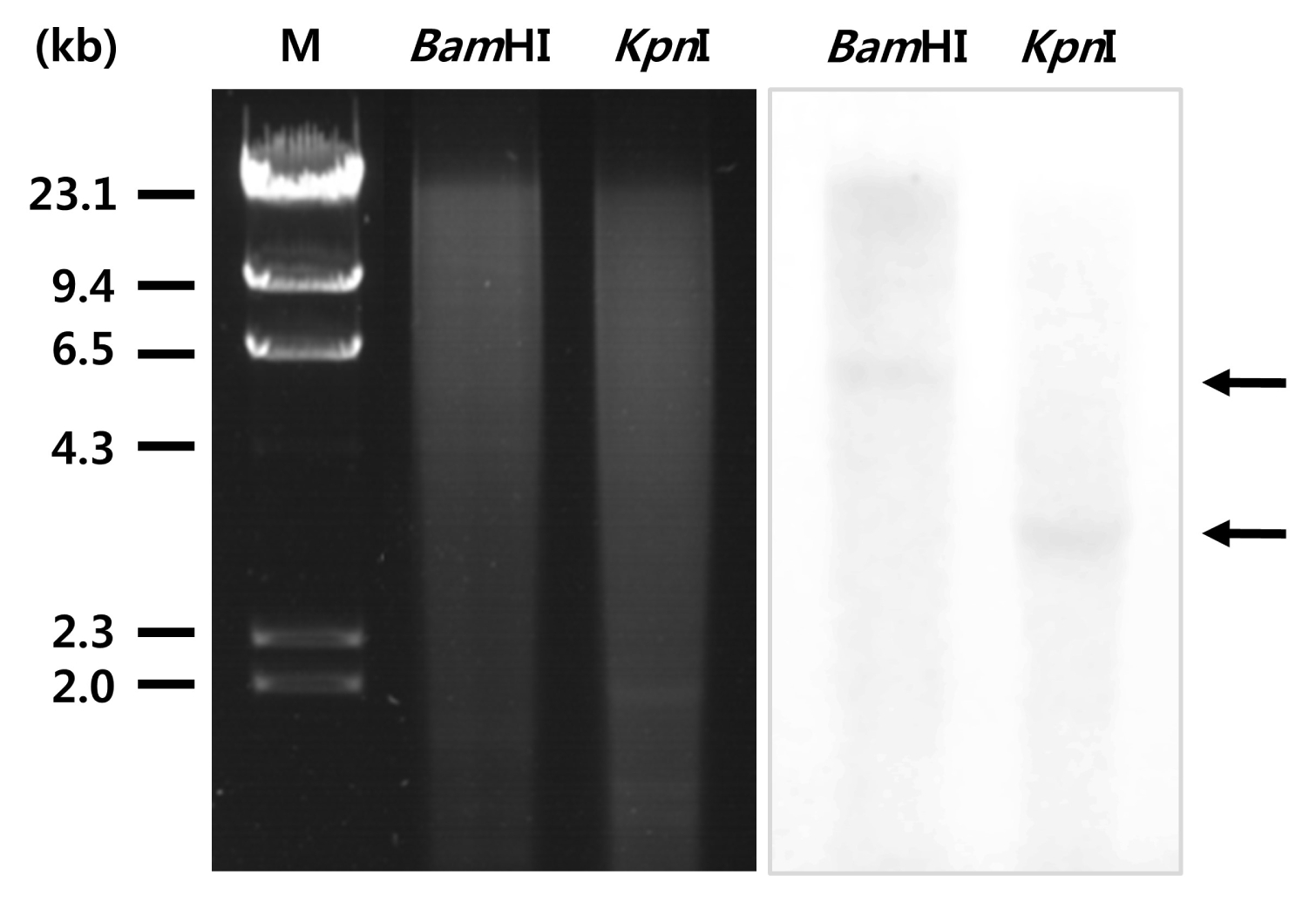
To determine the copy number of
SAMS in the
P. tenera genome, genomic DNA of
P. tenera was digested with restriction endonucleases
BamHI and
KpnI; then, Southern blot analysis was performed. After transferring to a membrane, the separated genomic DNA was hybridized to DIG-labeled
PtSAMS gene-specific probe. Southern blot analysis result clearly showed that the
PtSAMS gene was present as a single copy in
P. tenera (
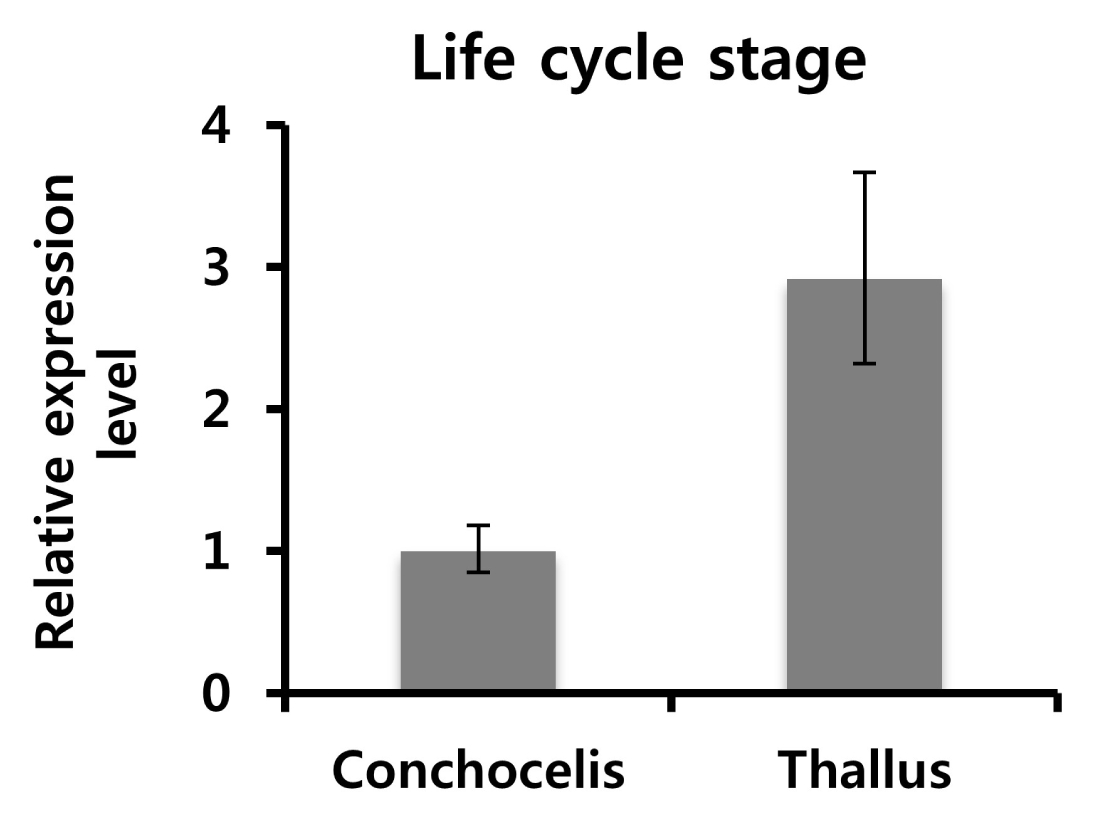
Fig. 2). To examine the expression levels of
PtSAMS transcripts at different life cycles, qPCR analysis was performed (
Fig. 3). Total RNA was isolated from the conchocelis and thallus of
P. tenera. The relative mRNA expression level in the thallus was higher than that in conchocelis by approximately three folds.
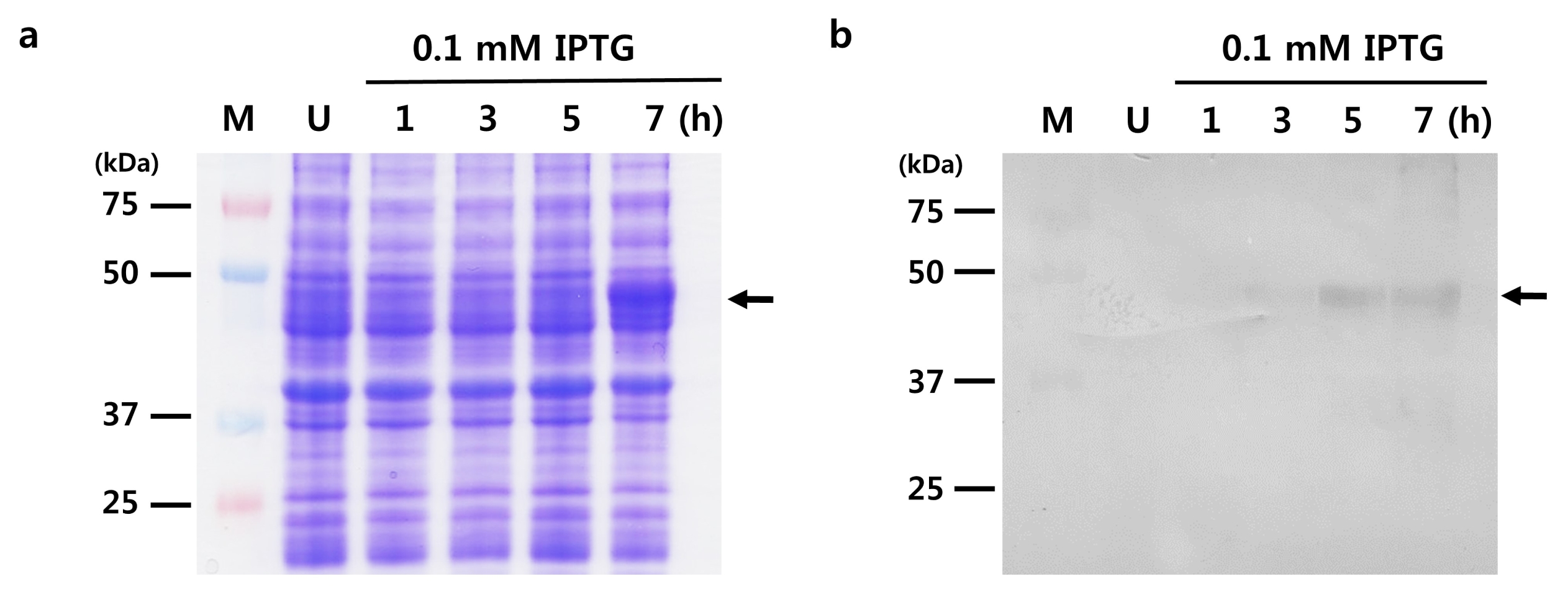
The complete ORF of the
PtSAMS gene was cloned into a pET28(b) vector and expressed in the
E. coli strain BL21(DE3). The empty pET28(b) vector was used as a control in this experiment. Expression of the recombinant PtSAMS protein was induced by adding 0.1 mM IPTG after 1 hour; this reached a maximum at 7 hours (
Fig. 4a). To confirm the production of His tag-fused PtSAMS, western blot analysis was carried out using anti-His (
Fig. 4b). The theoretical molecular weight of the recombinant PtSAMS was about 44.0 kDa.
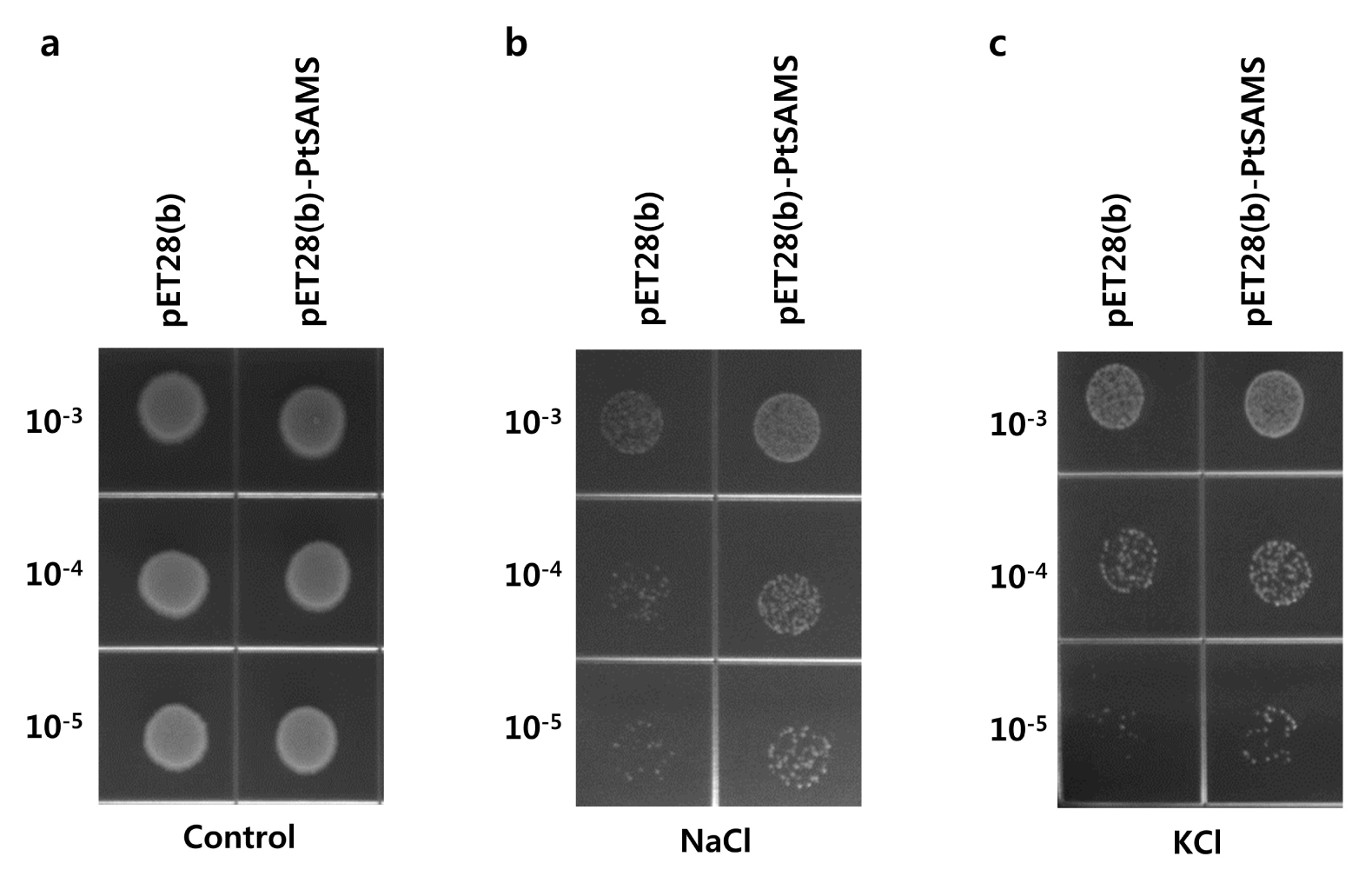
To examine the effect of the overexpression of the
PtSAMS protein in
E. coli under various abiotic stresses, the recombinant cells were spotted on LB medium supplemented with NaCl and KCl (
Fig. 5). The pET28(b)-
PtSAMS and pET28(b) cells showed similar growth on LB medium in overnight grown culture (
Fig. 5a). The pET28(b)-
PtSAMS recombinant cells showed increased number of colonies in NaCl and KCl treatment compared with that in control cells (
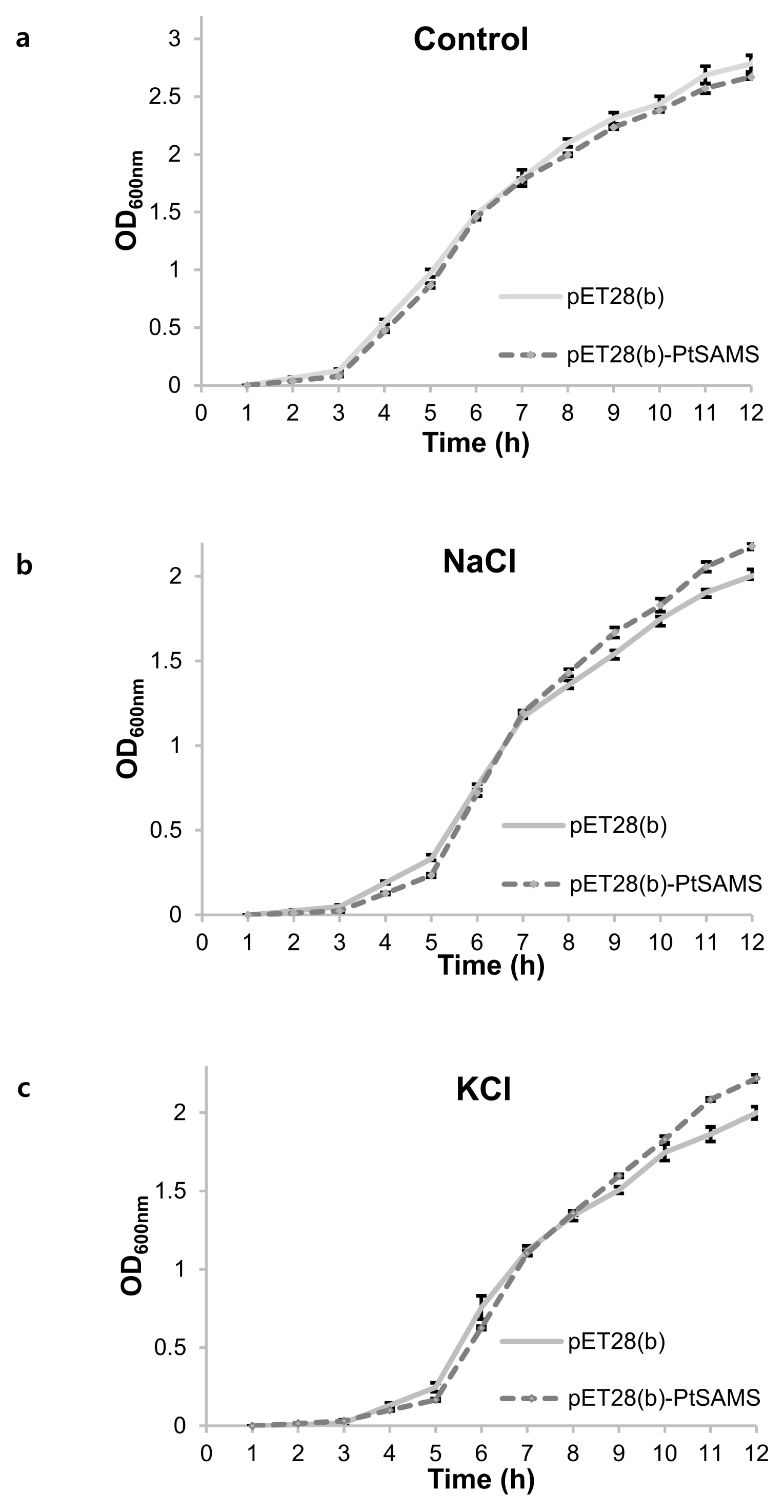
Fig. 5b and 5c). In addition, the growth rate of recombinant cells was analyzed in LB liquid medium. pET28(b)-
PtSAMS recombinant and pET28(b) empty vector control BL21(DE3) cells were inoculated into fresh LB liquid medium and medium supplemented with NaCl and KCl. In LB liquid medium, pET28(b)-
PtSAMS recombinant cell and cells harboring the pET28(b) vector alone showed similar growth at various times. In the NaCl- and KCl-treatment treatment, pET28(b)-
PtSAMS recombinant cells showed better growth compared with cells harboring the vector alone 6 hours after inoculation (
Fig. 6). Therefore, the spot and liquid culture assay indicated a similar pattern between the groups in terms of the abiotic stress response.
DISCUSSION
Pyropia grows in high intertidal zones where this plant experiences environmental conditions such as desiccation, salinity fluctuations, intense radiation, and high temperatures; additionally,
Pyropia is regularly submerged in seawater and exposed to air during high and low tides (
Xu et al. 2017). This plant has developed mechanisms to resist salt stress and survive under severe conditions. However, the molecular mechanisms underlying the salt stress tolerance in Rhodophyta are not fully understood. In this study, we attempted to isolate the salt stress gene in seaweed by cloning the
S-Adenosyl-l-methionine synthetase (SAMS) gene from
P. tenera; this gene was referred to as
PtSAMS. S-Adenosyl-L-methionine synthase synthesizes S-adenosyl-L-methionine (AdoMet) from L-methionine and ATP in both prokaryotes and eukaryotes (
Boerjan et al. 1994). In silico sequence alignment and phylogenetic analysis of amino acid sequence showed that PtSAMS comprises three conserved domains: the N-terminal domain, central domain, and C-terminal domain (
Fig. 1a). Furthermore, PtSAMS contains two motifs, Met-binding and ATP-binding sites, that are highly conserved in other plants. In the phylogenetic tree based on amino acid sequences of different SAMS proteins, PtSAMS was clustered with the SAMS from
G. sulphuraria, Rhodophyta (
Fig. 1b). Taken together, these results suggest that PtSAMS from
P. tenera is a member of the red algae SAMS family. Further, we identified the presence of a single copy of the
SAMS gene in the
Pyropia genome by DNA gel blot analysis (
Fig. 2). To examine
PtSAMS expression at various stages of the life cycle, RT-qPCR amplification of DNA from the conchocelis and thallus of
P. tenera was performed (
Fig. 3). The relative mRNA expression level of
PtSAMS in the thallus was higher than in the conchocelis. However, the expression level did not differ under exposure to abiotic stresses in the thallus (data not shown). As
Pyropia inhabits sea water, this organism is exposed to environmental stresses such as high salt levels and drought, and is therefore adapted for growth and survival in the marine environment (
Xu et al. 2017). It is therefore possible that the expression of the
PtSAMS gene is maintained at the level of a housekeeping gene owing to the constant exposure to such growing conditions.
Salinity stress is an important environmental factor that limits plant growth and development. SAM acts as a universal methyl group donor in biological processes during numerous specific trimethylations of protein, lipids, polysaccharides, and nucleic acids (
Li et al. 2011). SAM additionally serves as precursor for polyamine biosynthesis (
Evans and Malmberg 1989). The plant hormone, ethylene, is synthesized from SAM and participates in various physiological processes such as the stress response (
Wang et al. 2002;
Wang et al. 2013).
SAMS is expressed in various tissues in other plant species (
Li et al. 2011): in rice, the
SAMS gene regulates ethylene-mediated inhibition of root development and alteration of cell wall structures (
Fukuda et al. 2007).
SAMS expression is associated with the mechanism underlying tolerance of plants to abiotic stresses; it has been shown that transgenic
SAMS-expressing plants show enhanced resistance to abiotic stresses. In tomato,
SAMS were differentially expressed after application with salt stress (
Espartero et al. 1994). Overexpression of
SAMS in
M. sativa resulted in enhanced tolerance to cold stress by accelerating polyamine oxidation (
Guo et al. 2014). In addition, the levels of three
SAMS transcripts were increased transiently following the application of various stresses in
Catharanthus roseus; in particular, a high degree of accumulation of
SAM2 transcripts was observed (
Schroder et al. 1997). Taken together with the results of previous studies, the present findings suggest that the expression of the
SAMS gene maybe induced by a combination of ion toxicity and osmotic stress during exposure to salt stress.
E. coli growth assay results showed that pET8(b)-
PtSAMS recombinant
E. coli cells showed better tolerance to salt stress than cells harboring the control vector (
Fig. 5). Similar to our results, previous studies have reported that the expression of stress-induced genes enhances tolerance to various stresses in other plants. Overexpression of
Lycoris radiata SAMS recombinant protein was induced effectively under high salinity (NaCl and KCl) compared with that in control samples (
Li et al. 2013;
Cui et al. 2005).
Ma et al. (2003) found that the expression of the
SAMS gene from
Suaeda salsa is induced under NaCl stress (
Ma et al. 2003). Recently, it has been reported that the
AvSAMS gene confers aluminum stress tolerance and facilitates epigenetic gene regulation (
Ezaki et al. 2016).
SAMS is therefore known to play roles in various plant defense systems. The present results indicate that
PtSAMS maybe plays an important role in salt tolerance. This gene therefore suggests a potential bioresource for the genetic engineering of abiotic stress tolerance in plants.
ACKNOWLEDGEMENTS
This work was supported by a grant from the National Marine Biodiversity Institute Research Program (2017M 01300).
Fig. 1Amino acid sequence alignment of PtSAMS homologs and phylogenetic tree of SAMS from various species. (a) Comparison of amino acid sequences of Pyropia tenera SAMS with the SAMS of other species: Galdieria sulphuraria (XP_005703068.1), Ectocarpus siliculosus (CBJ30117.1), Chlamydomonas reinhardtii (XP_001696661.1), and Arabidopsis thaliana (NP_171751.1); identical residues are shaded black, and similar residues are shaded gray. The domains are indicated by underlining. Met-binding motifs are indicated by boxes. Dotted boxes indicate ATP-binding sites. (b) Phylogenetic tree analysis using Geneious software using Jukes Cantor, Neighbor Joining tree building method with a 10,000 bootstrap repeat value.
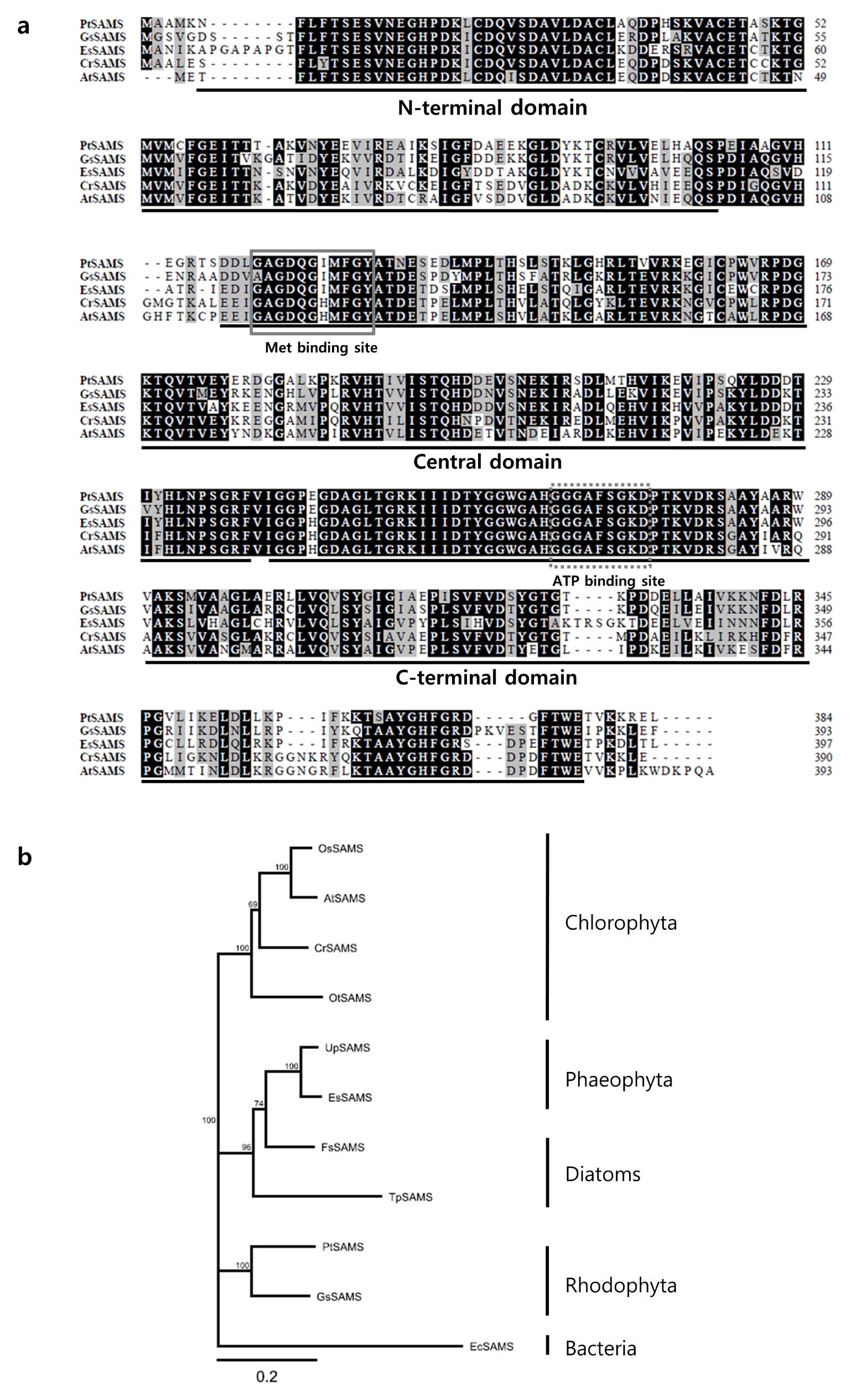
Fig. 2Southern blot analysis of P. tenera genomic DNA: 5 μg of genomic DNA per lane was digested with restriction enzymes and separated by electrophoresis in a 0.7% agarose gel. The DNA was transferred to a nylon membrane and probed with a DIG-labeled PtSAMS cDNA fragment. Molecular size markers are indicated on the left.
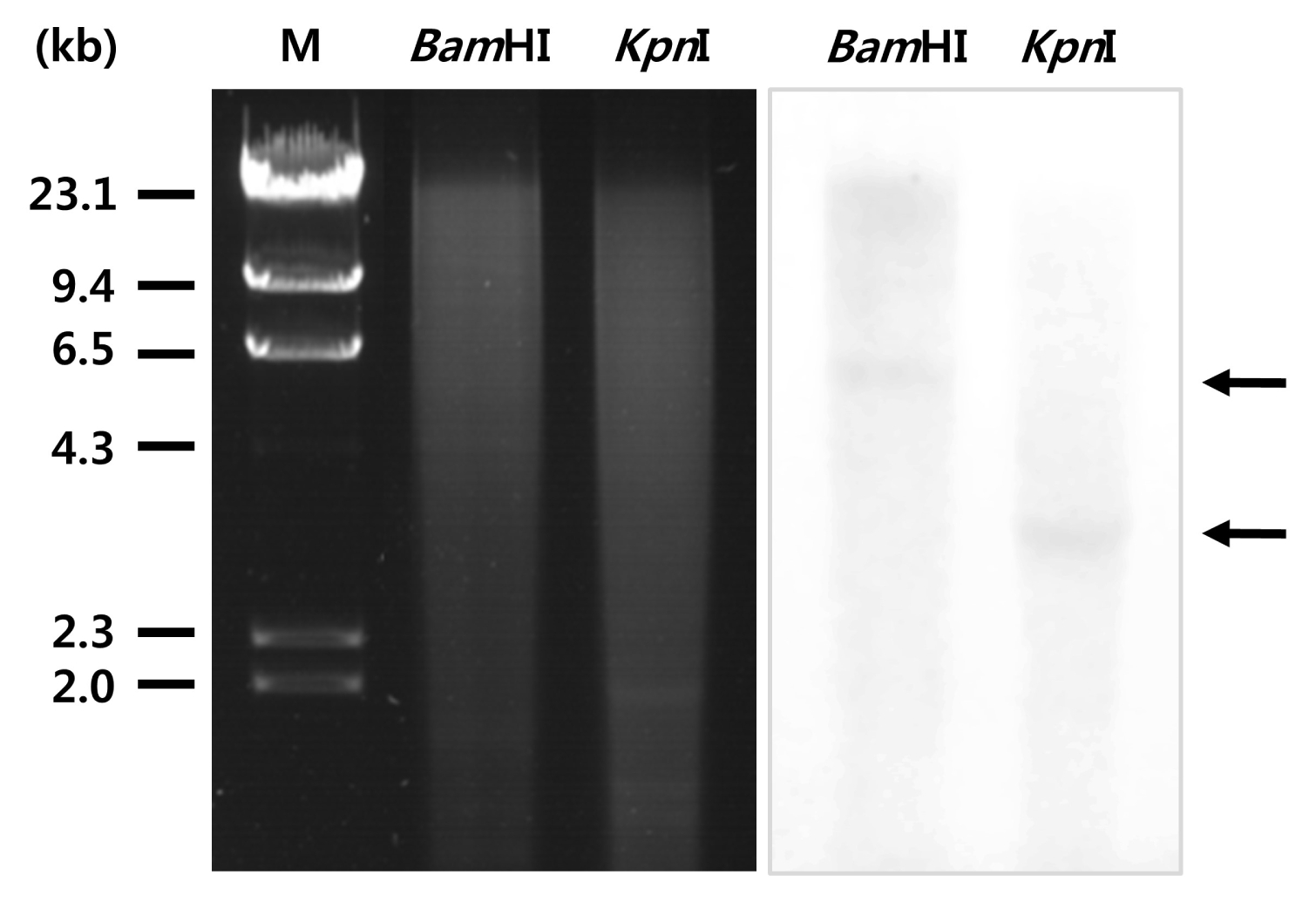
Fig. 3Expression pattern analysis of PtSAMS at different life cycle stages in P. tenera: Total RNA was isolated from conchocelis and thallus of P. tenera at various life stages. The mRNA expression level of the conchocelis stage was set to 1. PtGAPDH was used as normalization control. The results are presented as average values with SD using each three times.
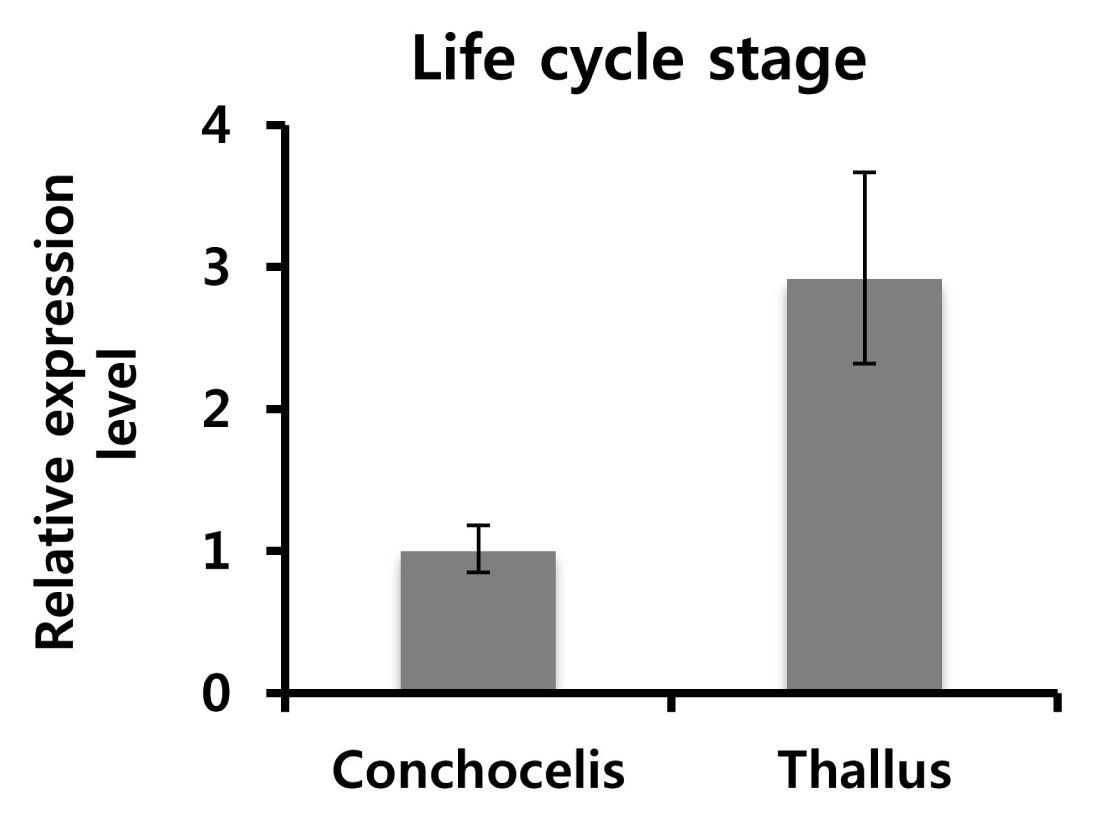
Fig. 4SDS-PAGE and western blot analysis of PtSAMS recombinant protein in E. coli BL21(DE3). (a) SDS-PAGE analysis. M: marker, Lane 1: uninduced protein, Lane 2–4: induced protein by 0.1 mM IPTG for 1, 3, 5 (hour), respectively. (b) Analysis of expression of recombinant PtSAMS protein in E. coli BL21(DE3) cells; western blotting analysis of the PtSAMS recombinant proteins with anti-His tag antibodies. Arrows indicate the His-tagged PtSAMS recombinant protein.
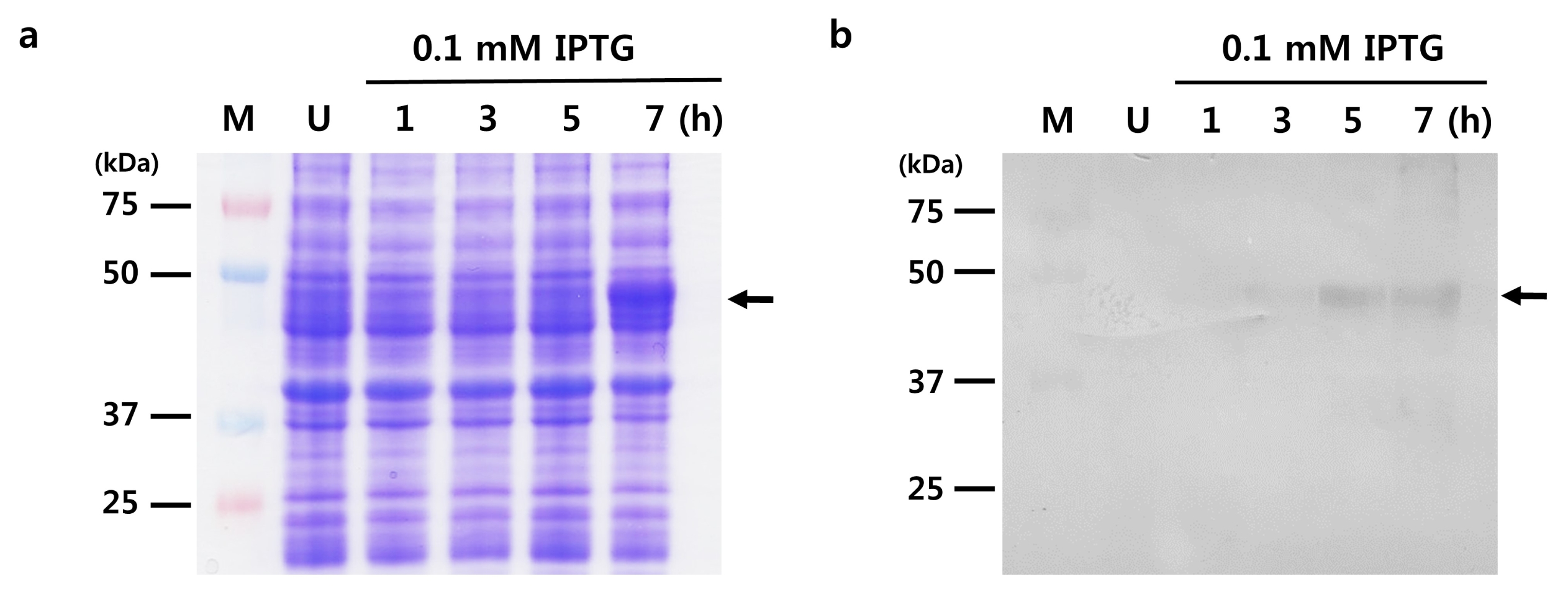
Fig. 5Spot assay of pET28(b) and pET28(b)-PtSAMS recombinant protein in E. coli cells with NaCl and KCl treatment; 10 μL from 10−3 to 10−5 dilution series were spotted on (a) control (LB-only) plate, or (b) LB plate supplemented with 400 mM NaCl, and (c) 400 mM KCl.
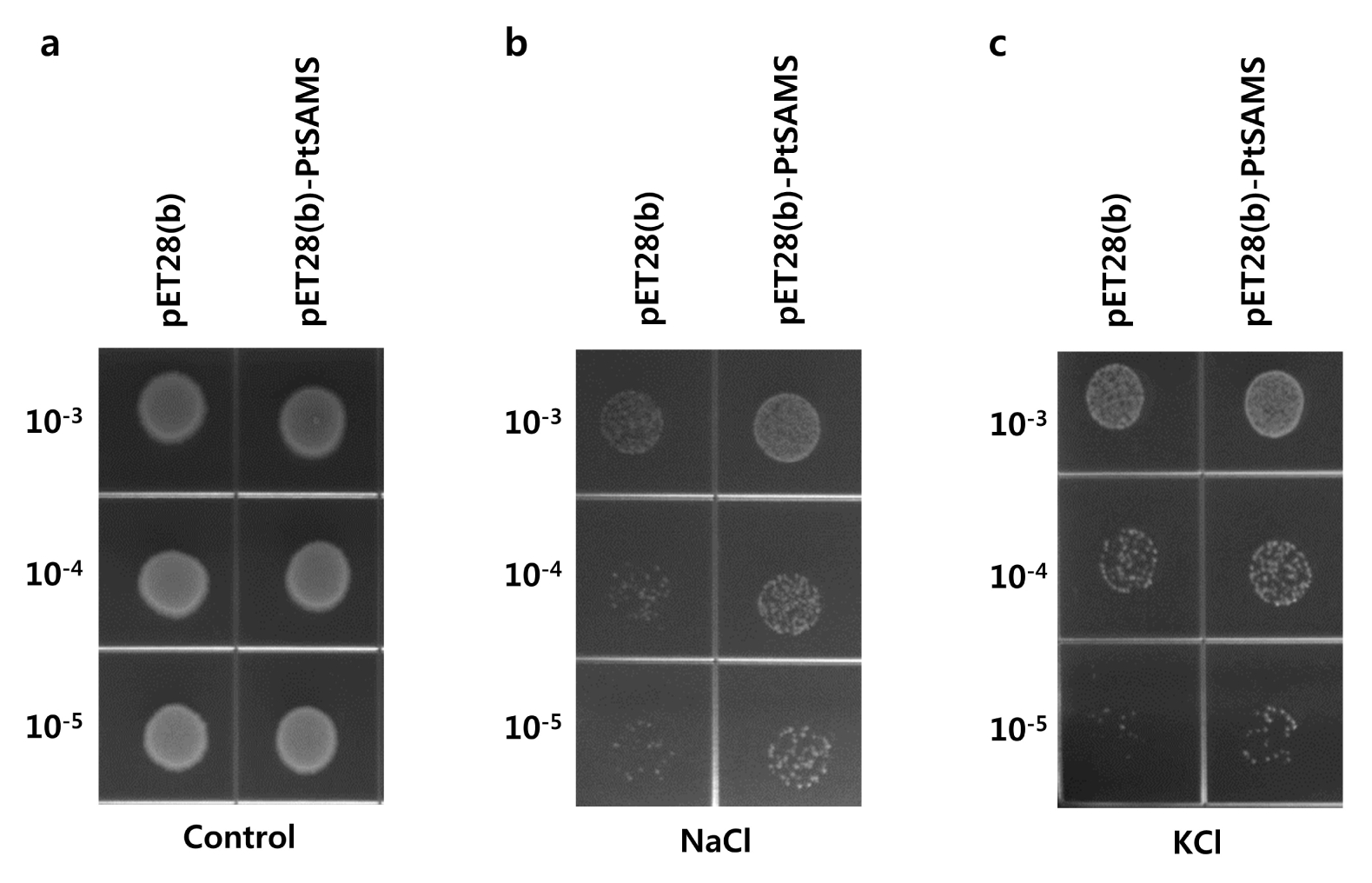
Fig. 6Growth analysis of pET28(b) and pET28(b)-PtSAMS recombinant protein in E. coli cells on liquid medium with NaCl and KCl treatment; (a) LB medium, (b) 400 mM NaCl, and (c) 400 mM KCl. OD600 was recored at 1 hour interval up to 12 hours and mean values are represented in graph.
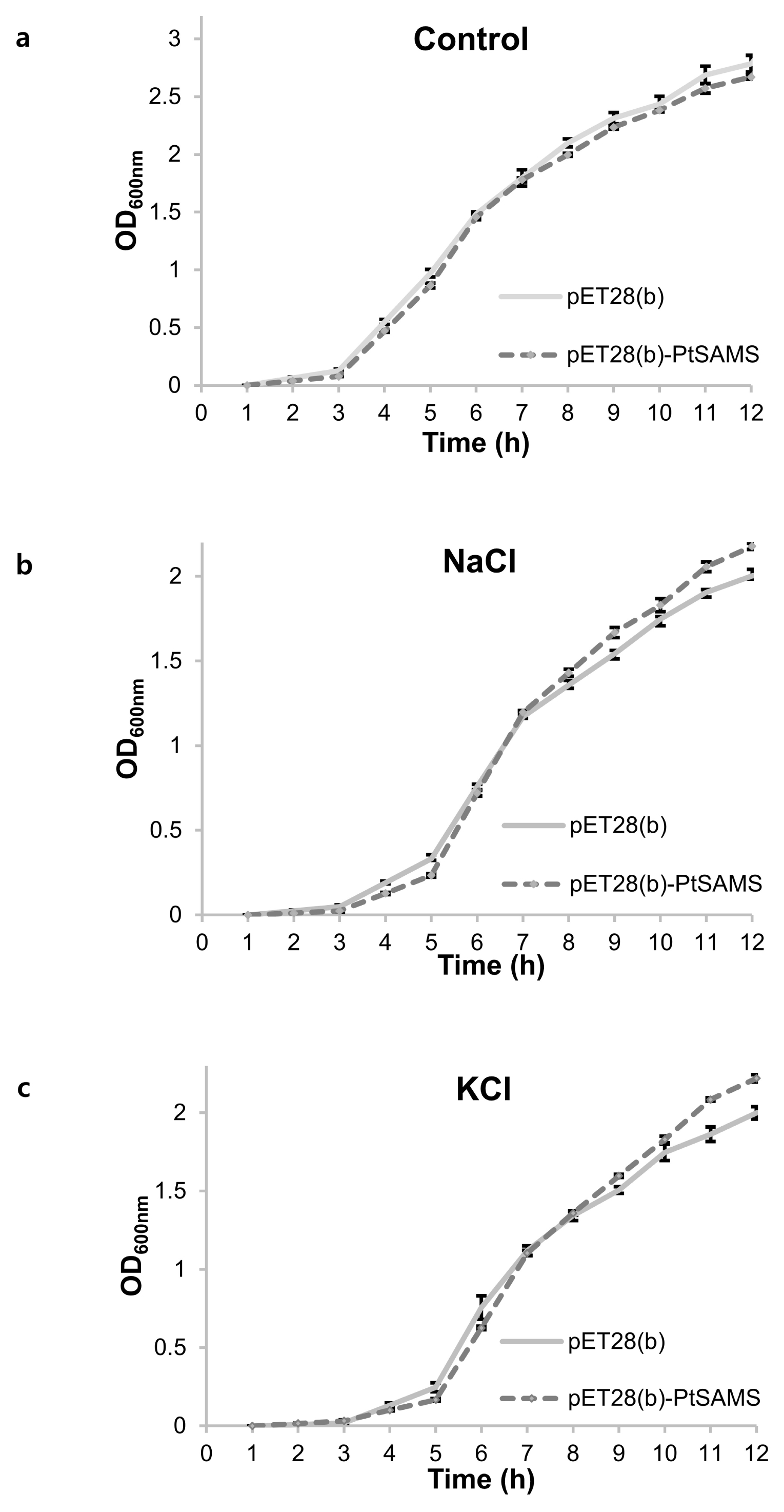
Table 1Gene specific primers used in the study.
Table 1
|
Name |
Sequence (5′-3′) |
Purpose |
|
PtSAMS-F |
CACCATGGCAGCCATGAAG |
For full length of PtSAMS gene |
|
PtSAMS-R |
ACGACGCTCTAGAGCTCAC |
For full length of PtSAMS gene |
|
PtSAMS-NdeI-F |
CCCCATATGGCAGCCATGAAGAz)
|
For protein expression |
|
PtSAMS-XhoI-R |
TCACTCGAGGAGCTCAAGCz)
|
For protein expression |
|
PtSAMS-303-F |
CCAGTCCCCTGAGATTGCTG |
Probe for southern blot analysis |
|
PtSAMS-955-R |
CTGAAATCGGCTCGGCAATG |
Probe for southern blot analysis |
|
PtSAMS-406-F |
GAGGACCTGATGCCCTTGAC |
RT-qPCR |
|
PtSAMS-591-R |
CTGCGTGCTGATGACAATCG |
RT-qPCR |
|
PtGAPDH-2771-F |
CGCCGAGTACATTGTCGAGT |
RT-qPCR |
|
PtGAPDH-3002-R |
GTACTTCTCGTGCAGCACCT |
RT-qPCR |
Table 2GenBank and NCBI reference sequence accession numbers of SAMS sequences.
Table 2
|
Species |
Amino acid sequence length |
NCBI reference sequence |
|
Rhodophyta |
Pyropia yezoensis
|
384 |
ACJ98094.1 |
|
Galdieria sulphuraria
|
393 |
XP_005703068.1 |
|
Phaeophyta |
Ectocarpus siliculosus
|
397 |
CBJ30117.1 |
|
Undaria pinnatifida
|
397 |
AEK80411.1 |
|
Chlorophyta |
Ostreococcus tauri
|
386 |
XP_003083240.1 |
|
Chlamydomonas reinhardtii
|
390 |
XP_001696661.1 |
|
Arabidopsis thaliana
|
393 |
NP_171751.1 |
|
Oryza sativa
|
396 |
AAT94053.1 |
|
Diatoms |
Thalassiosira pseudonana
|
450 |
XP_002288884.1 |
|
Fistulifera solaris
|
384 |
GAX17078.1 |
|
Bacterial |
Escherichia coli
|
384 |
AAA24164.1 |
Table 3Amino acid sequence identity matrix comparison of PtSMAS with SAMS of other species.
Table 3
|
PtSAMS |
GsSAMS |
CrSAMS |
EsSAMS |
AtSAMS |
|
PtSAMS |
100% |
76.84% |
70% |
69.6% |
66.75% |
|
GsSAMS |
|
100% |
69.77% |
68.91% |
67.59% |
|
CrSAMS |
|
|
100% |
65.92% |
81.14% |
|
EsSAMS |
|
|
|
100% |
64.25% |
|
AtSAMS |
|
|
|
|
100% |
References
- Amme S, Matros A, Schlesier B, Mock HP. 2006. Proteome analysis of cold stress response in Arabidopsis thaliana using DIGE-technology. J Exp Bot. 57: 1537-1546.
- Blouin NA, Brodie JA, Grossman AC, Xu P, Brawley SH. 2011. Porphyra: a marine crop shaped by stress. Trends Plant Sci. 16: 29-37.
- Boerjan W, Bauw G, Van Montagu M, Inze D. 1994. Distinct phenotypes generated by overexpression and suppression of S-adenosyl-L-methionine synthetase reveal developmental patterns of gene silencing in tobacco. Plant Cell. 6: 1401-1414.
- Cui S, Huang F, Wang J, Ma X, Cheng Y, Liu J. 2005. A proteomic analysis of cold stress responses in rice seedlings. Proteomics. 5: 3162-3172.
- Espartero J, Pintor-Toro JA, Pardo JM. 1994. Differential accumulation of S-adenosylmethionine synthetase transcripts in response to salt stress. Plant Mol Biol. 25: 217-227.
- Evans JM, Malmberg RL. 1989. Do polyamines have roles in plant develoiment? Annu Rev Plant Physiol Plant Mol Biol. 40: 235-269.
- Ezaki B, Higashi A, Nanba N, Nishiuchi T. 2016. An S-adenosyl methionine synthetase (SAMS) gene from Andropogon virginicus L. confers aluminum stress tolerance and facilitates epigenetic gene regulation in Arabidopsis thaliana. Front Plant Sci. 7: 1627
- Fukuda T, Saito A, Wasaki J, Shinano T, Osaki M. 2007. Metabolic alterations proposed by proteome in rice roots grown under low P and high Al concentration under low pH. Plant Sci. 172: 1157-1165.
- Gómez-Gómez L, Carrasco P. 1998. Differential expression of the S-adenosyl-L-methionine synthase genes during pea development. Plant Physiol. 117: 397-405.
- Guo Z, Tan J, Zhuo C, Wang C, Xiang B, Wang Z. 2014. Abscisic acid, H2O2 and nitric oxide interactions mediated cold-induced S-adenosylmethionine synthetase in Medicago sativa subsp. falcata that confers cold tolerance through up-regulating polyamine oxidation. Plant Biotechnol J. 12: 601-612.
- Hua Y, Zhang BX, Cai H, Li Y, Bai X, Ji W, Wang ZY, Zhu YM. 2012. Stress-inducible expression of GsSAMS2 enhances salt tolerance in transgenic Medicago sativa. African Journal of Biotechnology. 11: 4030-4038.
- Kakinuma M, Nakamoto C, Kishi K, Coury DA, Amano H. 2017. Isolation and functional characterization of an ammonium transporter gene, PyAMT1, related to nitrogen assimilation in the marine macroalga Pyropia yezoensis (Rhodophyta). Mar Environ Res. 128: 76-87.
- Kumar A, Taylor M, Altabella T, Tiburcio AF. 1997. Recent advances in polyamine research. Trends Plant Sci. 2: 124-130.
- Lee MK, Kim IH, Choi YH, Choi JW, Kim YM, Nam TJ. 2015. The proliferative effects of Pyropia yezoensis peptide on IEC-6 cells are mediated through the epidermal growth factor receptor signaling pathway. Int J Mol Med. 35: 909-914.
- Li W, Han Y, Tao F, Chong K. 2011. Knockdown of SAMS genes encoding S-adenosyl-l-methionine synthetases causes methylation alterations of DNAs and histones and leads to late flowering in rice. J Plant Physiol. 168: 1837-1843.
- Li XD, Xia B, Wang R, Xu S, Jiang YM, Yu FB, Peng F. 2013. Molecular cloning and characterization of S-adenosylmethionine synthetase gene from Lycoris radiata. Mol Biol Rep. 40: 1255-1263.
- Ma X, Wang Z, Qi Y, Zhao Y, Zhang H. 2003. Isolation of s-adenosylmethionine synthetase gene from Suaeda salsa and its differential expression under NaCl stress. Acta Botanica Sinica. 45: 1359-1365.
- Pajares MA, Markham GD. 2011. Methionine adenosyl-transferase (s-adenosylmethionine synthetase). Adv Enzymol Relat Areas Mol Biol. 78: 449-521.
- Pulla RK, Kim YJ, Parvin S, Shim JS, Lee JH, Kim YJ, In JG, Senthil KS, Yang DC. 2009. Isolation of S-adenosyl-L-methionine synthetase gene from Panax ginseng C.A. meyer and analysis of its response to abiotic stresses. Physiol. Mol. Biol. Plants. 15: 267-275.
- Qi YC, Wang FF, Zhang H, Liu WQ. 2010. Overexpression of suadea salsa S-adenosylmethionine synthetase gene promotes salt tolerance in transgenic tobacco. Acta Physiologiae Plantarum. 32: 263-269.
- Roeder S, Dreschler K, Wirtz M, Cristescu SM, van Harren FJ, Hell R, Piechulla B. 2009. SAM levels, gene expression of SAM synthetase, methionine synthase and ACC oxidase, and ethylene emission from N. suaveolens flowers. Plant Mol Biol. 70: 535-546.
- Schroder G, Eichel J, Breinig S, Schroder J. 1997. Three differentially expressed S-adenosylmethionine synthetases from Catharanthus roseus: molecular and functional characterization. Plant Mol Biol. 33: 211-222.
- Tester M, Bacic A. 2005. Abiotic stress tolerance in grasses. From model plants to crop plants. Plant Physiol. 137: 791-793.
- Thompson JD, Higgins DG, Gibson TJ. 1994. CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 22: 4673-4680.
- Uvackova L, Takac T, Boehm N, Obert B, Samaj J. 2012. Proteomic and biochemical analysis of maize anthers after cold pretreatment and induction of androgenesis reveals an important role of anti-oxidative enzymes. J Proteomics. 75: 1886-1894.
- Wang F, Cui X, Sun Y, Dong CH. 2013. Ethylene signaling and regulation in plant growth and stress responses. Plant Cell Rep. 32: 1099-1109.
- Wang KL, Li H, Ecker JR. 2002. Ethylene biosynthesis and signaling networks. Plant Cell. 14: Suppl. S131-151.
- Xu K, Xu Y, Ji D, Chen T, Chen C, Xie C. 2017. Cells tile a flat plane by controlling geometries during morphogenesis of Pyropia thalli. PeerJ. 5: e3314
- Yadav NS, Rashmi D, Singh D, Agarwal PK, Jha B. 2012. A novel salt-inducible gene SbSI-1 from Salicornia brachiata confers salt and desiccation tolerance in E coli. Mol Biol Rep. 39: 1943-1948.
- Yang PF, Li XJ, Liang Y, Jing YX, Shen SH, Kuang TY. 2006. Proteomic analysis of the response of Liangyoupeijiu (super high-yield hybrid rice) seedlings to cold stress. J Integr Plant Biol. 48: 945-951.
- Yi LF, Wang P, Zhou X, Liu C. 2009. cDNA cloning and bioinformatic analysis of SAMS gene from Porphyra yezoensis. China Biotechnology. 29: 43-49.