Abstract
Black rot of Brassica crops is the most devastating disease which causes substantial yield reduction of cabbage throughout the world. The use of resistant cabbage cultivars could be inexpensive and effective measure to combat this destructive disease. We screened cabbage inbred lines for black rot disease resistance through bioassay and identified some novel lines that showed race-specific resistance to Xanthomonas campestris pv. campestris (Xcc) races. The pathogenicity test revealed that out of 27 cabbage lines, one (SCNU-C-4074), six (SCNU-C-3631, SCNU-C-3637, SCNU-C-3639, SCNU-C-4072, SCNU-C-4073 and SCNU-C-3273), two (SCNU-C-3273 and SCNU-C-4118), two (SCNU-C-3270 and SCNU-C-4118), two (SCNU-C-3470 and SCNU-C-41148) and four (SCNU-C-107, SCNU-C-3270, SCNU-C-3470 and SCNU-C-4059) were shown to be resistant to Xcc races 1, 2, 3, 5, 6 and 7, respectively while none of these showed resistance against race 4. Furthermore, these resistant and susceptible lines were evaluated by previously reported molecular markers for black rot resistance. The molecular screening results were also revealed the existence of race-specific resistance in these cabbage lines. This result will help Brassica breeder to develop race-specific black rot resistant cabbage cultivars.
-
Key words: Black rot resistance, Xanthomonas campestris pv. campestris, Cabbage, Bioassay, Molecular markers
INTRODUCTION
Cabbage (
Brassica oleracea L.) is the most important vegetable among
Brassica crops in the world due to its nutritional and health benefits (
Lee et al. 2015). Black rot disease, caused by
Xanthomonas campestris pv.
campestris (
Xcc) (Pammel) Dowson, is the most destructive diseases of the crops belonging to the Brassicaceae family (
Williams 1980;
Lema et al. 2012;
Vicente and Holub 2013;
Lee et al. 2015). The major host of
Xcc is
B. oleracea and its subspecies (
Vicente et al. 2001).
Xcc is mainly a seed born pathogen, but can alive in crop residues and cruciferous weeds and ornamentals (
Cook et al. 1952;
Roberts et al. 1999;
Vicente et al. 2001;
Lema et al. 2011). This pathogen can enter into the leaf via insects, wounded tissues and hydathodes (leaf margin) and spreads through vascular tissues (
Tonu et al. 2013).
Xcc can spread via rain, wind, insects, agricultural equipments and irrigation water (
Sharma et al. 2017). The characteristic disease symptoms are V-shaped chlorotic lesions at the leaf margins, necrotic and darkened veins which reduce the quality and production (
Williams 1980;
Kifuji et al. 2013;
Tonu et al. 2013;
Vicente and Holub 2013;
Lee et al. 2015). In favorable condition, black rot disease can reduce crop yield more than 50% (
Williams 1980). However, up to 100% yield loss has been reported in cabbage by farmers in Tanzania (
Massomo et al. 2003). Black rot also decreases the market value of cabbage. Like other plant pathogenic bacteria,
Xcc has been separated into different physiological races based on the response of differential cultivars with resistance genes and pathogens. Up to now, 11 physiological races of
Xcc have been reported (
Kamoun et al. 1992;
Vicente et al. 2001;
Fargier and Manceau 2007;
Cruz et al. 2017). Initially,
Kamoun et al. (1992) reported five
Xcc races (0–4). Later,
Vicente et al. (2001) reclassified
Xcc into six races (1–6). Furthermore,
Jensen et al. (2010) and
Fargier and Manceau (2007) were added race 7 and races 8–9 to Vicente’s classification, respectively. Newly, race 10 and race 11 have reported in Portugal (
Cruz et al. 2017).
The sources of resistance for black rot in
B. oleracea are very less (
Ignatov et al. 1998). However, the existence of black rot resistance was reported in Japanese cabbage (Early Fuji, P143660) as well as in Penca kales (
Bain 1952;
Dickson and Hunter 1987;
Ferreira et al. 1992;
Ignatov et al. 1999).
Ignatov et al. (1998) reported race-specific resistance to
Xcc races 1 and 5 in Japanese cabbage lines and Penca kale landraces. The race-specific resistance to
Xcc in Asian cabbages was said to be inherited from Penca kales (a Portuguese black rot resistant kale landrace) (
Ignatov et al. 1998). There were many research reports describing
Xcc resistant genes in
B. oleracea and related species.
Williams et al. (1972) have been stated that black rot is controlled by one major gene designated as
f and the heterozygous condition is influenced by one dominant and one modifier gene. They screened 300 cultivars and inbred lines of cabbage for
Xcc resistance and this
f gene found only in Early Fuji, a Japanese black rot resistant cabbage cultivar.
Dickson and Hunter (1987) reported that black rot resistance is governed by a single recessive gene in cabbage (PI436606, a cabbage line from China). There are some reported quantitative trait loci (QTLs) and associated markers for black rot resistance in different
Brassica species. For instance,
Camargo et al. (1995) described QTLs for black rot resistance based on restriction fragment length polymorphism (RFLP) loci in cabbage. They found that two QTLs on linkage group (LG) 1 and 9 showed resistance against black rot for both young and old plants while two additional QTLs on LG 2 only for young plant resistance.
Kifuji et al. (2013) reported a QTL for black rot resistance using 161 EST-SNP markers and found that QTL-1 located on LG C02 is the major QTL in cabbage. A major black rot resistance locus called Xca1bo has been mapped on Chromosome 03 in Indian cauliflower (
Saha et al. 2014).
Lee et al. (2015) reported a genetic linkage map where they identified four QTLs such as BRQTL-C1_1 and BRQTL-C1_2 on LG C01, BRQTL-C3 on LG C03 and BRQTL-C6 on LG 06. Among these QTLs, BRQTL-C1_2 designates as the most important QTL for black rot in cabbage while remaining three as minor QTLs.
Sharma et al. (2016) reported black rot resistance locus Xca1bc on LG B-7 in Indian mustard (
Brassica carinata). They also reported that black rot resistance in
B. carinata was controlled by a single dominant gene. Now-a-days, molecular marker based genotyping are extensively used to screen disease resistance. However, the resistance sources in
B. oleracea (C genome) are limited while major resistance sources have been described in A and B genomes (
Taylor et al. 2002;
Sharma et al. 2016).
Xcc can be retained in seeds,
Brassica crop residues, and cruciferous weed and ornamentals which act as the sources of inoculum (
Schaad and Dianese 1981). Thus, it is very difficult to control black rot disease through disease management practices including agrochemicals. For this reason, exploration of
Xcc resistant sources is one of the best ways to abate crop loss from black rot disease. However, new pathogenic races can be evolved due to the intensive cultivation of resistant cultivars that is the main hinder of black rot resistance (
Song et al. 2014). Therefore, in this study, we aimed to screen out novel cabbage lines which provide race-specific resistance against
Xcc races.
MATERIALS AND METHODS
Plant materials
Twenty seven different cabbage (
Brassica oleracea L.) inbred lines were tested in this study (
Supplementary Table S1). Seeds were obtained from Department of Horticulture, Sunchon National University, Republic of Korea. Initially, seeds were sown in trays containing a nursery pot mixture at plant culture room (25±1°C temperature, 60% Relative humidity and 80–120 μmol/m
2/second light intensity). Twenty two days after sowing, the seedlings were transferred into the pot and kept at the glasshouse. For pathogenicity test, seedlings from each of the above mentioned inbred lines were used for screening against black rot disease.
A total of seven
Xanthomonas campestris pv.
campestris (
Xcc) races (race 1, 2, 3, 4, 5, 6 and 7) were inoculated in the aforementioned cabbage line for pathogenicity test which are shown in
Table 1. The
Xcc races (race 1–7) were collected from the School of Life Sciences, University of Warwick, UK. All the
Xcc races were grown on King’s medium B (KB) for 48 hours at 30°C (
King et al. 1954).
The cabbage inbred lines were inoculated at 35 days after sowing. The
Xcc races were sub-cultured on KB at 30°C for 48 hours before inoculation. Thereafter, the bacteria were scraped from the culture plates and suspended in 10 mL of sterile water. Finally, three youngest leaves of each plants were inoculated by clipping secondary veins with sterile forceps (maintained at least 10 inoculation points per leaf) followed by dipping into a bacterial suspension (10
8–10
9 CFU/mL) of
Xcc races and maintained high humidity (
Vicente et al. 2001). Three seedlings for each
Xcc race were used for the inoculation. The disease symptoms of inoculated leaves were evaluated at 2 and 3 weeks after inoculation (WAI). The disease reaction was scored at 2 WAI for each inoculated leaf based on a 0–9 scale (
Fig. 1) where 0 = no visible symptoms; 1 = small necrosis or chlorosis near the inoculation point; 3 = typical small V-shaped lesion with black veins; 5 = typical lesion half way to the middle vein; 7 = typical lesion succeeding to the middle vein; and 9 = lesion reaching the middle vein as previously described by
Vicente et al. (2002). The highly resistant (HR), resistant (R), susceptible (S) and highly susceptible (HS) lines were characterized based one the scales 0, 2–3, 5–7, and 9, respectively (
Fig. 1).
The young leaves of each cabbage lines were collected and immediately frozen in liquid nitrogen. Then, the leaf samples were maintained at −80°C until use. The genomic DNA was isolated using a commercial kit (DNeasy Plant Mini Kit, QIAGEN, Germany) following the manufacturer’s instructions. The integrity and purity of the DNA were assessed by gel electrophoresis (0.8% agarose) and Nanodrop ND-1000 (NanoDrop Technologies Inc., Wilmington, DE, USA), respectively.
Polymerase chain reaction (PCR)
Previously reported markers were used to screen the cabbage inbred lines for black rot resistance (
Table 2). The PCR amplification was performed in 20 μL reaction mixture, containing of 2× Prime Taq premix (Genet Bio, Republic of Korea), 10 pmoles of each forward and reverse primers (Macrogen Inc., Seoul, Republic of Korea) and 100 ng genomic DNA as template. The thermal cycle was set with an initial denaturation at 95°C for 5 minutes, followed by 30–35 cycles of denaturation at 95°C for 30 seconds, annealing at specific Tm to respective primer sets (
Table 2) between 52 and 61°C for 30 seconds, extension at 72°C for 30 seconds and a final extension at 72°C for 5 minutes. Then, the PCR products were subjected to agarose gel electrophoresis to visualize the bands. The concentration of agarose gel was varied based on amplicon size. Finally, results were compared with the bioassay results.
The polymorphic markers for black rot disease (BoGMS0971, BnGMS301, BoESSR291, BoESSR726 and OI10G06) were amplified by PCR using Phusion
® High-Fidelity DNA Polymerase (New England Biolabs) from resistant (R) and susceptible (S) cabbage lines. The polymorphic bands were extracted from the gel and purified using the Wizard SV gel and PCR cleanup system (Promega, Madison, WI, USA) from both resistant and susceptible lines for each of the above mentioned markers. Then, cloning was done using TOP cloner blunt kit (Enzynomics, Daejeon, Republic of Korea) according to the manufacturer’s instruction. Three independent PCR positive clones were sequenced with the universal primers (M13FpUC and M13RpUC) by ABI 3730XL DNA sequencer (Macrogen Inc., Seoul, Republic of Korea). Subsequently, the cloned sequences of R and S lines for each marker were aligned using the Clustal Omega online Multiple Sequence Alignment tool available at
https://www.ebi.ac.uk/Tools/msa/clustalo/.
RESULTS
Screening of cabbage inbred lines for the black rot resistance
The disease reaction results of 27 cabbage inbred lines for black rot disease resistance are shown in
Table 3 and
Fig. 2. The tested cabbage lines provide race-specific resistance against different
Xanthomonas campestris pv.
campestris (
Xcc) races. The result showed that cabbage line SCNU-C-4074 was resistant (
Fig. 2) against
Xcc race 1. A total of six cabbage lines were highly resistant to
Xcc race 2 (SCNU-C-3273, SCNU-C-3631, SCNU-C-3637, SCNU-C-3639, SCNU-C-4072 and SCNU-C-4073). Similarly, two lines SCNU-C-3273 and SCNU-C-4118 were resistant to
Xcc race 3. Likewise, lines SCNU-C-3270 (highly resistant) and SCNU-C-4118 (resistant) were resistant to
Xcc race 5. Two cabbage lines (SCNU-C-3470 and SCNU-C-41148) were shown to be resistant against
Xcc race 6. A total of four cabbage lines (SCNU-C-107, SCNU-C-3270, SCNU-C-3470 and SCNU-C-4059) were resistant to
Xcc race 7 of which three lines (SCNU-C-3270, SCNU-C-3470 and SCNU-C-4059) were highly resistant. On the other hand, all of the tested lines were susceptible for
Xcc race 4. However, some cabbage lines were resistant to more than one
Xcc races. For example, line SCNU-C-3270 shown to be resistant against race 5 and 7 (highly resistant), line SCNU-C-3273 against race 2 (highly resistant) and 3, and SCNU-C-4118 against race 3, 5 and 6.
A total of 10 previously published markers (9 SSR and 1 InDel) for black rot resistance were selected for this study based on reported QTL information. Among those reported markers, only five showed polymorphic amplification and were able to distinguish resistant (R) and susceptible (S) cabbage lines against black rot disease (
Table 3 and
Supplementary Fig. S1). Thereafter, the bioassay and molecular screening results were compared and those having more than 60% adaptability for each race were shown in
Table 3. The outcome of the comparison showed that the OI10G06 marker separates R and S lines with 83.3% adaptability, respectively for
Xcc race 1 followed by BnGMS301 (79.2%), BoESSR726 (79.2%) and BoESSR291 (66.7%). Similarly, BoESSR291 and OI10G06 can differentiate R and S lines against
Xcc race 2 with 73.9% adaptability followed by BoESSR726 (65.2%) and BoGMS0971 (60.9%). In case of race 3, BnGMS301 can differentiate R and S lines with 81.5% adaptability followed by BoESSR291 (77.8%), BoESSR726 (74.1%) and OI10G06 (70.1%). For race 4, BnGMS301 showed 79.2% match with the bioassay results followed by BoESSR726 (75%), OI10G06 (70.8%) and BoESSR291 (66.7%). In case of race 5, BnGMS301 showed 91.3% match with the phenotypic screening results followed by BoESSR726 (78.3%), BoESSR291 (73.9%) and OI10G06 (69.6%). For race 6, BnGMS301 and BoESSR726 can separated R and S cabbage lines with 81.5% adaptability with the bioassay results followed by BoESSR291 (70.4%) and OI10G06 (66.7%). In case of race 7, BnGMS301, BoESSR291, OI10G06 and BoESSR726 showed 72.0%, 68.0%, 68.0% and 64.0% match with the bioassay results, respectively.
The amplified polymorphic markers were further cloned and sequenced from R and S lines. Through sequencing, the genetic variations including insertion/deletions (InDels), single nucleotide polymorphisms (SNPs) between R and S lines for the polymorphic markers were detected (
Supplementary Fig. S2). The BoGMS0971 marker amplified a 351-bp DNA fragment for R lines and 330-bp for S lines. The sequence alignment results showed that there was a total 21-bp deletions from S lines compared to R lines. This marker located on the intergenic region between two genes (Bo8g092870 and Bo8g092880) (
Table 4). The BnGMS301 amplified 238-bp DNA fragment for S lines but 215-bp with total 27-bp deletion for R lines, which are located between two genes (Bo1g053300 and Bo1g053310). In case of the BoESSR291, the tested cabbage lines showed three different types of PCR bands. These bands were 114-bp DNA fragment for R lines and another two with InDels, where 21-bp deletions and 16-bp insertions from R line resulted in being susceptible to black rot disease. However, BoESSR291 showed high identity with a genomic fragment where putative promoter sequence of a gene encoding enzyme cystathionine gamma-synthase (Bo3g055310) was located. The BoESSR726 marker amplified a 238-bp DNA fragment for the R line while 228-bp for S line. There were 12-bp InDels for R line and 2-bp InDel for S line which are located in the intergenic region between genes with accession number Bo1g054960 and Bo1g054970. The OI10G06 marker amplified 127-bp DNA fragment for S lines and one with 18-bp deletions for R lines (109-bp), which are located in the intergenic region of two genes (Bo6g095520 and Bo6g095530) (
Table 4).
DISCUSSION
Black rot is considered as one of the main threats of cabbage production in the world and it is very difficult to control. The race-specific resistance in
Brassica crops is linked to the hypersensitive response at inoculated area with incompatible race, nonetheless occasionally partial symptom appeared (
Kamoun et al. 1992;
Ignatov et al. 1997). However, recent reports described
Xcc race 1, 4 and 6 as the major races while remaining as the minor races in
B. oleracea (
Ignatov et al. 1998;
Vicente et al. 2001;
Taylor et al. 2002;
Chidamba and Bezuidenhout 2012;
Lema et al. 2012;
Rouhrazi and Khodakaramian 2014). In
B. oleracea,
Taylor et al. (2002) reported that 43% of the tested accessions were resistant to one or more of the minor
Xcc races (2, 3, 5, and 6) and only one showed partial resistance to
Xcc races 1, 3, 5, and 6. They also noted that resistance is more common to
Xcc race 3 and 5 whereas very rare to race 1 in
B. oleracea. Moreover,
Xcc race 6 was predominant in
B. rapa (
Lema et al. 2015). But recently
Cruz et al. (2017) reported the existence of races 4, 6 and 7 (encompassing 21 out of 33 isolates) in Portugal. In this study, we identified novel cabbage lines with race-specific hypersensitive response against different races of
Xcc. The disease reaction results revealed that there were 1, 6, 2, 2, 2 and 4 cabbage inbred lines exhibiting resistance against
Xcc race 1, 2, 3, 5, 6 and 7, respectively while none of tested lines were resistant to race 4 (
Fig. 2 and
Table 3). This result is consistent with previous report as described the race-specific resistance against
Xcc in
Brassica species by
Ignatov et al. (1998). Besides,
Tonu et al. (2013) also reported black rot resistance in
B. oleracea in particular to
Xcc race 3 and race 5. It should be noted that the phenotypic screening results revealed SCNU-C-4059 as resistant lines against race 7 but none of the tested markers correlated with the phenotype except BoGMS0971 which showed only 36% adaptability with bioassay results (
Table 3).
The genetic inheritance of major black rot resistance genes has been elucidated in different
Brassica species including
B. rapa,
B. carinata and
B. napus containing A, BC and AC genome, respectively (
Guo et al. 1991;
Ignatov et al. 2000;
Vicente et al. 2002). However, the strong resistance source for
Xcc race 4 was reported to be originated from A genome while resistance to both races 1 and 4 from B genome, and
B. juncea (AB genome) was the most resistant species to races 1 and 4 (
Taylor et al. 2002). To date, several QTLs for black rot resistance have been described in
B. oleracea and other
B. species (
Camargo et al. 1995;
Cheng et al. 2009;
Kifuji et al. 2013;
Izzah et al. 2014;
Saha et al. 2014;
Lee et al. 2015;
Sharma et al. 2016). In this study, five reported SSR markers showed polymorphism in banding pattern for R and S lines of the tested cabbage lines. By using these markers, we were able to separate R and S lines which were consistent with bioassay based phenotypic screening results (
Table 3 and
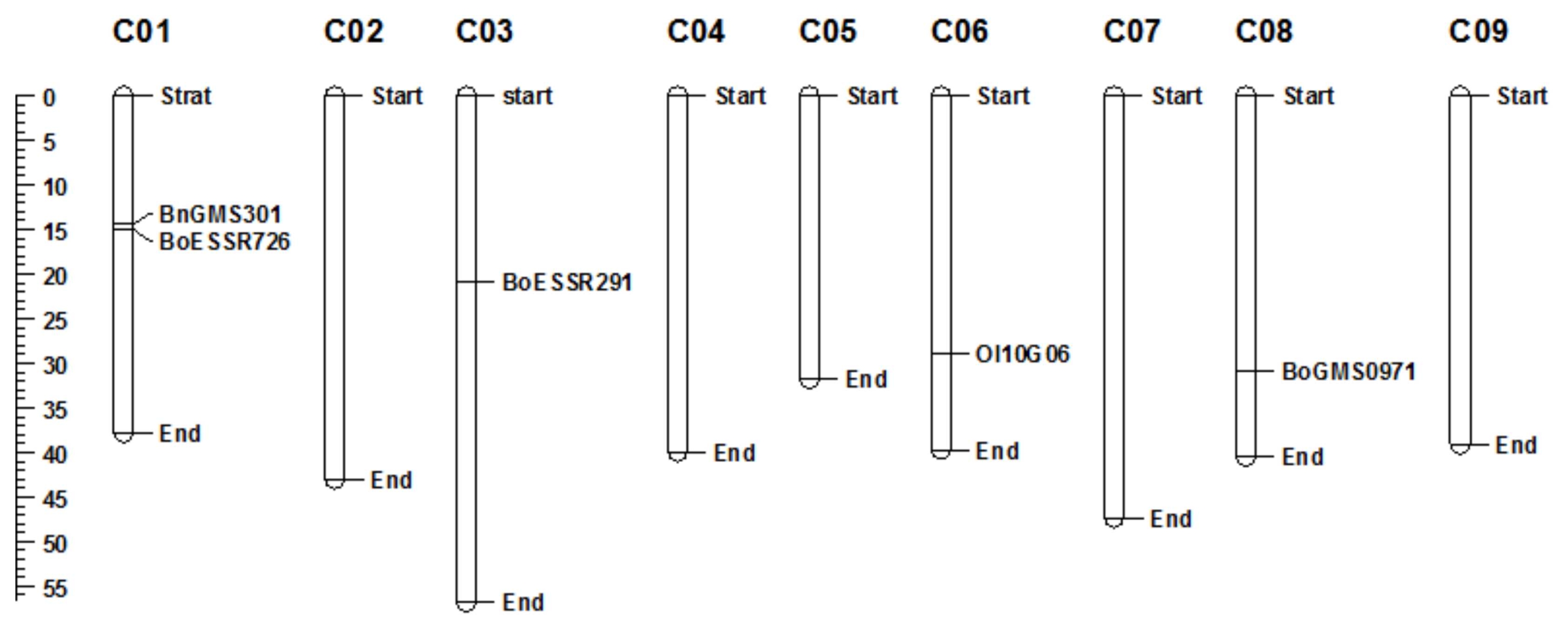
Fig. 2), nevertheless, none of them perfectly match with phenotypic data. These published markers were located on four different chromosomes such as BnGMS301 and BoESSR726 on chromosome C01, BoESSR291 on chromosome C03, OI10G06 on chromosome C06, and BoGMS0971 on chromosome C08 (
Fig. 3). Previous reports described several QTLs for black rot resistance on those chromosomes. For instance,
Lee et al. (2015) described four QTLs on three different chromosomes where BRQTL-C1_2 on chromosome C01 was the main QTL for black rot resistance in cabbage. Interestingly, the tested polymorphic marker BnGMS301 located on chromosome C01.
Tonu et al. (2013) found XccBo(Reiho)2 on chromosome C08 as the major locus for black rot resistance in cabbage and the SSR marker BoGMS0971 was closely linked to it. The remaining two SSR markers namely BoESSR291 and OI10G06 were linked to the minor QTLs (BRQTL-C3 and BRQTL-C6, respectively) (
Lee et al. 2015). Plant disease resistance governed by an interaction between specific disease resistance gene (
R) of the host plant and
avirulence gene (
avr) the pathogen and also known as “gene-for-gene” model for plant disease resistance (
Flor 1971;
Dangl and Jones 2001). The
R genes codes for only five classes of proteins and those encoded “nucleotide-binding site and leucine-rich-repeat” (NBS-LRR) proteins are the major
R genes in “gene-for-gene” plant resistance (
Van der Biezen and Jones 1998;
Dangl and Jones 2001). The NBS-LRR type
R genes were further subdivided into two classes based on N-terminal structures such as those having the intracellular signaling domains of the
Drosophila Toll and mammalian interleukin (IL)-1 receptors (TIR-NB-LRR), and others having coiled-coil domains (CC-NB-LRR) (
Dangl and Jones 2001). None of the tested markers were located in neither the
R genes nor any other annotated genes (
Table 5). Therefore, it is crucial to identify
R genes responsible for race-specific resistance in cabbage. Taking this into consideration, we performed crossing between R and S cabbage lines for each
Xcc race (race 1, 2, 3, 5, 6 and 7) to develop mapping population. Thereafter, with molecular mapping, the specific
R genes responsible for race-specific resistance could be elucidated.
Supplementary Information
ACKNOWLEDGEMENTS
This paper was supported by the Sunchon National University Research Fund in 2017. We thank Dr Joana G. Vicente, School of Life Sciences, University of Warwick, UK for providing Xanthomonas campestris pv. campestris races.
Fig. 1The disease reaction scoring criteria used in this study for black rot of cabbage. Scales: 0 = no visible symptoms; 1 = small necrosis or chlorosis near the inoculation point; 3 = typical small V-shaped lesion with black veins; 5 = typical lesion half way to the middle vein; 7 = typical lesion succeeding to the middle vein; and 9 = lesion reaching the middle vein (
Vicente et al. 2002). 0, high resistant (HR); 1–3, resistant (R); 5–7, susceptible (S); 9, highly susceptible (HS).

Fig. 2The resistant and susceptible cabbage lines against different races of Xanthomonas campestris pv. campestris (Xcc) at 2 weeks after inoculation. The letters a, b, c, d, e, f and g, represents Xcc race 1, 2, 3, 4, 5, 6 and 7, respectively.

Fig. 3Chromosomes location of five reported polymorphic markers on Brassica oleracea.
Table 1 Xanthomonas campestris pv. campestris (Xcc) races used for bioassay-based screening of cabbage lines.
Table 1
|
Sl. No. |
Bacterial race/strains |
Source |
Reference |
|
1 |
Xanthomonas campestris pv. campestris Race 1 (B100) |
UK |
, Vicente et al. 2001
|
|
2 |
Xanthomonas campestris pv. campestris Race 2 (3849A) |
US |
|
|
3 |
Xanthomonas campestris pv. campestris Race 3 (5212) |
UK |
|
|
4 |
Xanthomonas campestris pv. campestris Race 4 (CFBP 5817) |
UK |
|
|
5 |
Xanthomonas campestris pv. campestris Race 5 (3880) |
Australia |
|
|
6 |
Xanthomonas campestris pv. campestris Race 6 (6181) |
Portugal |
|
|
7 |
Xanthomonas campestris pv. campestris Race 7 (8450A) |
UK |
|
Table 2List of markers with their primer sequence used in the molecular screening for black rot resistance in cabbage.
Table 2
|
Sl. No. |
Markers |
Chromosome |
Primer sequence (5′……3′) |
Tm (°C) |
Size (bp) |
Marker type |
Reference |
|
1 |
BoESSR291 |
C03 |
F: AAGCTGGGGATGGAGAAGAT
R: GCACCTAATCGAACCCCTTA |
52 |
114 |
SSR |
, Izzah et al. 2014
|
|
2 |
BoESSR216 |
C01 |
F: GGTTTCCGCTATGTCCAGAA
R: CGGAAGAAGACGTTGAGGAG |
52 |
317 |
SSR |
, Izzah et al. 2014
|
|
3 |
BoESSR089 |
C01 |
F: ATGATCAGCGAAACCACTCC
R: TGATACATCCCGTTTGCTCA |
55 |
259 |
SSR |
, Izzah et al. 2014
|
|
4 |
BoESSR145 |
C01 |
F: GGGCGAGGATGGTTACTACA
R: TCATACCCCAAGGCTATTTT |
55 |
240 |
SSR |
, Izzah et al. 2014
|
|
5 |
BoESSR726 |
C01 |
F: CAATGGGTTACGCATGGTTT
R: CGTTTGTGAAACAGCCATTG |
55 |
228 |
SSR |
, Izzah et al. 2014
|
|
6 |
SSR739 |
C03 |
F: TAGGGTGAAAGGGAAGCTCA
R: CGCTAATAATGGCGCTAAGG |
52 |
182 |
SSR |
, Saha et al. 2014
|
|
7 |
BnGMS301 |
C01 |
F: AATATGCAGCATTCTAGACAAA
R: ATCATTCTCGTGATGACACA |
55 |
250 |
SSR |
, Cheng et al. 2009
|
|
8 |
BoGMS0971 |
C08 |
F: TAATCCGAACAACACGAA
R: CACCCAATAAGCGATGAG |
52 |
344 |
SSR |
, Li et al. 2011
|
|
9 |
OI10G06 |
C06 |
F: GACAAGTTCCCTTGTAATGGC
R:TGTAATCATCACACATTTTGGG |
52 |
109 |
SSR |
, Li et al. 2011
|
|
10 |
Iso0857_371bc |
C09 |
F: AAGGAATTCCCCAGGATGTC
R: TTGGCAACCCTAATGCTTTT |
51 |
477 |
InDel |
, Lee et al. 2015
|
Table 3Comparison of bioassay and molecular screening results for black rot resistance in cabbage.
Table 3
|
Sl. No. |
Lines |
Race 1 |
OI10G06 |
BnGMS301 |
BoESSR726 |
BoESSR291 |
Race 2 |
BoESSR291 |
OI10G06 |
BoESSR726 |
BoGMS0971 |
|
|
|
|
|
Pz)
|
Gy)
|
Pz)
|
Gy)
|
|
1 |
SCNU-C-106 |
HSu)
|
+ |
+ |
+ |
+ |
HS |
+ |
+ |
+ |
− |
|
2 |
SCNU-C-107 |
n/at)
|
− |
+ |
+ |
− |
HS |
− |
− |
+ |
+ |
|
3 |
SCNU-C-3122 |
Sv)
|
+ |
− |
+ |
+ |
HS |
+ |
+ |
+ |
+ |
|
4 |
SCNU-C-3270 |
S |
+ |
− |
+ |
+ |
S |
+ |
+ |
+ |
+ |
|
5 |
SCNU-C-3273 |
S |
+ |
+ |
+ |
− |
HRx)
|
− |
+ |
+ |
− |
|
6 |
SCNU-C-3305 |
HS |
+ |
+ |
+ |
+ |
S |
+ |
+ |
+ |
+ |
|
7 |
SCNU-C-3328 |
S |
+ |
− |
− |
+ |
HS |
+ |
+ |
− |
+ |
|
8 |
SCNU-C-3412 |
S |
+ |
+ |
+ |
+ |
HS |
+ |
+ |
+ |
− |
|
9 |
SCNU-C-3414 |
S |
− |
+ |
+ |
− |
n/a |
− |
− |
+ |
− |
|
10 |
SCNU-C-3449 |
HS |
+ |
+ |
+ |
+ |
HS |
+ |
+ |
+ |
+ |
|
11 |
SCNU-C-3470 |
n/a |
+ |
+ |
− |
+ |
n/a |
+ |
+ |
− |
+ |
|
12 |
SCNU-C-3631 |
S |
− |
+ |
+ |
− |
HR |
− |
− |
+ |
− |
|
13 |
SCNU-C-3637 |
S |
+ |
+ |
+ |
− |
HR |
− |
+ |
+ |
− |
|
14 |
SCNU-C-3639 |
S |
+ |
+ |
− |
+ |
HR |
+ |
+ |
− |
+ |
|
15 |
SCNU-C-4059 |
S |
+ |
+ |
+ |
+ |
S |
+ |
+ |
+ |
− |
|
16 |
SCNU-C-4066 |
S |
+ |
+ |
+ |
− |
S |
− |
+ |
+ |
− |
|
17 |
SCNU-C-4071 |
S |
− |
+ |
+ |
+ |
S |
+ |
− |
+ |
− |
|
18 |
SCNU-C-4072 |
n/a |
− |
+ |
+ |
+ |
HR |
+ |
− |
+ |
− |
|
19 |
SCNU-C-4073 |
S |
+ |
+ |
+ |
+ |
HR |
+ |
+ |
+ |
− |
|
20 |
SCNU-C-4074 |
Rw)
|
− |
− |
− |
+ |
n/a |
+ |
− |
− |
− |
|
21 |
SCNU-C-4085 |
S |
+ |
+ |
+ |
+ |
S |
+ |
+ |
+ |
− |
|
22 |
SCNU-C-4105 |
S |
+ |
+ |
− |
+ |
S |
+ |
+ |
− |
+ |
|
23 |
SCNU-C-4118 |
S |
+ |
− |
− |
− |
S |
− |
+ |
− |
+ |
|
24 |
SCNU-C-4161 |
HS |
+ |
+ |
+ |
+ |
S |
+ |
+ |
+ |
− |
|
25 |
SCNU-C-4168 |
S |
− |
+ |
+ |
− |
S |
− |
− |
+ |
− |
|
26 |
SCNU-C-4302 |
HS |
+ |
+ |
− |
+ |
HS |
+ |
+ |
− |
+ |
|
27 |
SCNU-C-4320 |
HS |
+ |
+ |
+ |
+ |
HS |
+ |
+ |
+ |
− |
|
Matched |
|
20.0 |
19.0 |
19.0 |
16.0 |
|
17.0 |
17.0 |
15.0 |
14.0 |
|
Not matched |
|
4.0 |
5.0 |
5.0 |
9.0 |
|
6.0 |
6.0 |
8.0 |
9.0 |
|
% Adaptability |
|
83.3 |
79.2 |
79.2 |
66.7 |
|
73.9 |
73.9 |
65.2 |
60.9 |
|
Sl. No. |
Lines |
Race 3 |
BnGMS301 |
BoESSR291 |
BoESSR726 |
OI10G06 |
Race 4 |
BnGMS301 |
BoESSR726 |
OI10G06 |
BoESSR291 |
|
|
|
|
|
Pz)
|
Gy)
|
Pz)
|
Gy)
|
|
1 |
SCNU-C-106 |
HS |
+ |
+ |
+ |
+ |
n/a |
+ |
+ |
+ |
+ |
|
2 |
SCNU-C-107 |
HS |
+ |
− |
+ |
− |
S |
+ |
+ |
− |
− |
|
3 |
SCNU-C-3122 |
HS |
− |
+ |
+ |
+ |
HS |
− |
+ |
+ |
+ |
|
4 |
SCNU-C-3270 |
HS |
− |
+ |
+ |
+ |
S |
− |
+ |
+ |
+ |
|
5 |
SCNU-C-3273 |
R |
+ |
− |
+ |
+ |
S |
+ |
+ |
+ |
− |
|
6 |
SCNU-C-3305 |
S |
+ |
+ |
+ |
+ |
HS |
+ |
+ |
+ |
+ |
|
7 |
SCNU-C-3328 |
HS |
− |
+ |
− |
+ |
HS |
− |
− |
+ |
+ |
|
8 |
SCNU-C-3412 |
HS |
+ |
+ |
+ |
+ |
S |
+ |
+ |
+ |
+ |
|
9 |
SCNU-C-3414 |
S |
+ |
− |
+ |
− |
S |
+ |
+ |
− |
− |
|
10 |
SCNU-C-3449 |
HS |
+ |
+ |
+ |
+ |
HS |
+ |
+ |
+ |
+ |
|
11 |
SCNU-C-3470 |
S |
+ |
+ |
− |
+ |
S |
+ |
− |
+ |
+ |
|
12 |
SCNU-C-3631 |
S |
+ |
− |
+ |
− |
S |
+ |
+ |
− |
− |
|
13 |
SCNU-C-3637 |
S |
+ |
− |
+ |
+ |
S |
+ |
+ |
+ |
− |
|
14 |
SCNU-C-3639 |
S |
+ |
+ |
− |
+ |
S |
+ |
− |
+ |
+ |
|
15 |
SCNU-C-4059 |
S |
+ |
+ |
+ |
+ |
S |
+ |
+ |
+ |
+ |
|
16 |
SCNU-C-4066 |
HS |
+ |
− |
+ |
+ |
HS |
+ |
+ |
+ |
− |
|
17 |
SCNU-C-4071 |
HS |
+ |
+ |
+ |
− |
S |
+ |
+ |
− |
+ |
|
18 |
SCNU-C-4072 |
HS |
+ |
+ |
+ |
− |
n/a |
+ |
+ |
− |
+ |
|
19 |
SCNU-C-4073 |
HS |
+ |
+ |
+ |
+ |
S |
+ |
+ |
+ |
+ |
|
20 |
SCNU-C-4074 |
HS |
− |
+ |
− |
− |
HS |
− |
− |
− |
+ |
|
21 |
SCNU-C-4085 |
HS |
+ |
+ |
+ |
+ |
HS |
+ |
+ |
+ |
+ |
|
22 |
SCNU-C-4105 |
HS |
+ |
+ |
− |
+ |
S |
+ |
− |
+ |
+ |
|
23 |
SCNU-C-4118 |
R |
− |
− |
− |
+ |
S |
− |
− |
+ |
− |
|
24 |
SCNU-C-4161 |
HS |
+ |
+ |
+ |
+ |
n/a |
+ |
+ |
+ |
+ |
|
25 |
SCNU-C-4168 |
HS |
+ |
− |
+ |
− |
S |
+ |
+ |
− |
− |
|
26 |
SCNU-C-4302 |
HS |
+ |
+ |
− |
+ |
S |
+ |
− |
+ |
+ |
|
27 |
SCNU-C-4320 |
HS |
+ |
+ |
+ |
+ |
HS |
+ |
+ |
+ |
+ |
|
Matched |
|
22.0 |
21.0 |
20.0 |
19.0 |
|
19.0 |
18.0 |
17.0 |
16.0 |
|
Not matched |
|
5.0 |
6.0 |
7.0 |
8.0 |
|
5.0 |
6.0 |
7.0 |
8.0 |
|
% Adaptability |
|
81.5 |
77.8 |
74.1 |
70.1 |
|
79.2 |
75.0 |
70.8 |
66.7 |
|
Sl. No. |
Lines |
Race 5 |
BnGMS301 |
BoESSR726 |
BoESSR291 |
OI10G06 |
Race 6 |
BnGMS301 |
BoESSR726 |
BoESSR291 |
OI10G06 |
|
|
|
|
|
Pz)
|
Gy)
|
Pz)
|
Gy)
|
|
1 |
SCNU-C-106 |
HS |
+ |
+ |
+ |
+ |
HS |
+ |
+ |
+ |
+ |
|
2 |
SCNU-C-107 |
HS |
+ |
+ |
− |
− |
HS |
+ |
+ |
− |
− |
|
3 |
SCNU-C-3122 |
S |
− |
+ |
+ |
+ |
HS |
− |
+ |
+ |
+ |
|
4 |
SCNU-C-3270 |
R |
− |
+ |
+ |
+ |
S |
− |
+ |
+ |
+ |
|
5 |
SCNU-C-3273 |
n/a |
+ |
+ |
− |
+ |
HS |
+ |
+ |
− |
+ |
|
6 |
SCNU-C-3305 |
S |
+ |
+ |
+ |
+ |
HS |
+ |
+ |
+ |
+ |
|
7 |
SCNU-C-3328 |
HS |
− |
− |
+ |
+ |
HS |
− |
− |
+ |
+ |
|
8 |
SCNU-C-3412 |
S |
+ |
+ |
+ |
+ |
S |
+ |
+ |
+ |
+ |
|
9 |
SCNU-C-3414 |
n/a |
+ |
+ |
− |
− |
HS |
+ |
+ |
− |
− |
|
10 |
SCNU-C-3449 |
HS |
+ |
+ |
+ |
+ |
S |
+ |
+ |
+ |
+ |
|
11 |
SCNU-C-3470 |
n/a |
+ |
− |
+ |
+ |
R |
+ |
− |
+ |
+ |
|
12 |
SCNU-C-3631 |
S |
+ |
+ |
− |
− |
HS |
+ |
+ |
− |
− |
|
13 |
SCNU-C-3637 |
HS |
+ |
+ |
− |
+ |
HS |
+ |
+ |
− |
+ |
|
14 |
SCNU-C-3639 |
HS |
+ |
− |
+ |
+ |
S |
+ |
− |
+ |
+ |
|
15 |
SCNU-C-4059 |
S |
+ |
+ |
+ |
+ |
S |
+ |
+ |
+ |
+ |
|
16 |
SCNU-C-4066 |
HS |
+ |
+ |
− |
+ |
HS |
+ |
+ |
− |
+ |
|
17 |
SCNU-C-4071 |
HS |
+ |
+ |
+ |
− |
HS |
+ |
+ |
+ |
− |
|
18 |
SCNU-C-4072 |
HS |
+ |
+ |
+ |
− |
HS |
+ |
+ |
+ |
− |
|
19 |
SCNU-C-4073 |
HS |
+ |
+ |
+ |
+ |
HS |
+ |
+ |
+ |
+ |
|
20 |
SCNU-C-4074 |
n/a |
− |
− |
+ |
− |
S |
− |
− |
+ |
− |
|
21 |
SCNU-C-4085 |
S |
+ |
+ |
+ |
+ |
HS |
+ |
+ |
+ |
+ |
|
22 |
SCNU-C-4105 |
HS |
+ |
− |
+ |
+ |
S |
+ |
− |
+ |
+ |
|
23 |
SCNU-C-4118 |
R |
− |
− |
− |
+ |
R |
− |
− |
− |
+ |
|
24 |
SCNU-C-4161 |
S |
+ |
+ |
+ |
+ |
HS |
+ |
+ |
+ |
+ |
|
25 |
SCNU-C-4168 |
HS |
+ |
+ |
− |
− |
HS |
+ |
+ |
− |
− |
|
26 |
SCNU-C-4302 |
HS |
+ |
− |
+ |
+ |
S |
+ |
− |
+ |
+ |
|
27 |
SCNU-C-4320 |
HS |
+ |
+ |
+ |
+ |
HS |
+ |
+ |
+ |
+ |
|
Matched |
|
21.0 |
18.0 |
17.0 |
16.0 |
|
22.0 |
22.0 |
19.0 |
18.0 |
|
Not matched |
|
2.0 |
5.0 |
6.0 |
7.0 |
|
5.0 |
5.0 |
8.0 |
9.0 |
|
% Adaptability |
|
91.3 |
78.3 |
73.9 |
69.6 |
|
81.5 |
81.5 |
70.4 |
66.7 |
|
Sl. No. |
Lines |
Race 7 |
BnGMS301 |
BoESSR291 |
OI10G06 |
BoESSR726 |
|
|
|
Pz)
|
Gy)
|
|
1 |
SCNU-C-106 |
HS |
+ |
+ |
+ |
+ |
|
2 |
SCNU-C-107 |
R |
+ |
− |
− |
+ |
|
3 |
SCNU-C-3122 |
HS |
− |
+ |
+ |
+ |
|
4 |
SCNU-C-3270 |
HR |
− |
+ |
+ |
+ |
|
5 |
SCNU-C-3273 |
n/a |
+ |
− |
+ |
+ |
|
6 |
SCNU-C-3305 |
S |
+ |
+ |
+ |
+ |
|
7 |
SCNU-C-3328 |
HS |
− |
+ |
+ |
− |
|
8 |
SCNU-C-3412 |
HS |
+ |
+ |
+ |
+ |
|
9 |
SCNU-C-3414 |
n/a |
+ |
− |
− |
+ |
|
10 |
SCNU-C-3449 |
S |
+ |
+ |
+ |
+ |
|
11 |
SCNU-C-3470 |
HR |
+ |
+ |
+ |
− |
|
12 |
SCNU-C-3631 |
HS |
+ |
− |
− |
+ |
|
13 |
SCNU-C-3637 |
HS |
+ |
− |
+ |
+ |
|
14 |
SCNU-C-3639 |
S |
+ |
+ |
+ |
− |
|
15 |
SCNU-C-4059 |
HR |
+ |
+ |
+ |
+ |
|
16 |
SCNU-C-4066 |
S |
+ |
− |
+ |
+ |
|
17 |
SCNU-C-4071 |
HS |
+ |
+ |
− |
+ |
|
18 |
SCNU-C-4072 |
HS |
+ |
+ |
− |
+ |
|
19 |
SCNU-C-4073 |
S |
+ |
+ |
+ |
+ |
|
20 |
SCNU-C-4074 |
HS |
− |
+ |
− |
− |
|
21 |
SCNU-C-4085 |
HS |
+ |
+ |
+ |
+ |
|
22 |
SCNU-C-4105 |
HS |
+ |
+ |
+ |
− |
|
23 |
SCNU-C-4118 |
HS |
− |
− |
+ |
− |
|
24 |
SCNU-C-4161 |
HS |
+ |
+ |
+ |
+ |
|
25 |
SCNU-C-4168 |
HS |
+ |
− |
− |
+ |
|
26 |
SCNU-C-4302 |
S |
+ |
+ |
+ |
− |
|
27 |
SCNU-C-4320 |
HS |
+ |
+ |
+ |
+ |
|
Matched |
|
18.0 |
17.0 |
17.0 |
16.0 |
|
Not matched |
|
7.0 |
8.0 |
8.0 |
9.0 |
|
% Adaptability |
|
72.0 |
68.0 |
68.0 |
64.0 |
Table 4The tested molecular markers and their location on Brassica oleracea genome.
Table 4
|
Maker name |
Locationz)
|
Description |
|
BoGMS0971 |
C8:31754531..31754753 |
Intergenic region (Bo8g092870 and Bo8g092880) |
|
BnGMS301 |
C1:15386406..15386643 |
Intergenic region (Bo1g053300 and Bo1g053310) |
|
BoESSR291 |
C3:21846613..21846644 |
Promoter region of a gene cystathionine gamma-synthase (Bo3g055310) |
|
BoESSR726 |
C1:15967972..15968174 |
Intergenic region (Bo1g054960 and Bo1g054970) |
|
OI10G06 |
C6:29898028..29898121 |
Intergenic region (Bo6g095520 and Bo6g095530) |
Table 5Genes around the tested polymorphic molecular markers in Brassica oleracea genome.
Table 5
|
Makers |
Gene ID |
Description |
|
|
NCBIz)
|
TAIRy)
|
Bolbasex)
|
|
BoGMS0971 |
Bo8g092870 |
V-type proton ATPase subunit D-like |
Vacuolar ATP synthase subunit D |
V-type proton ATPase subunit D-like |
|
Bo8g092880 |
Citrate synthase 2, peroxisomal-like |
Citrate synthase 1 |
Citrate synthase-like, large alpha subdomain |
|
BnGMS301 |
Bo1g053300 |
(E)-beta-ocimene synthase, chloroplastic |
Terpene synthase 02 |
Terpene synthase-like |
|
Bo1g053310 |
(E)-beta-ocimene synthase, chloroplastic |
Terpene synthase 02 |
Uncharacterized protein |
|
BoESSR291 |
Bo3g055310 |
Cystathionine gamma-synthase |
Cystathionine gamma-synthase |
Cystathionine gamma-synthase (CGS) |
|
BoESSR726 |
Bo1g054960 |
Uncharacterized |
ABC transporter family protein |
ABC transporter D family member 2, chloroplastic |
|
Bo1g054970 |
Uncharacterized |
Mitochondrial transcription termination factor family protein |
Mitochodrial transcription termination factor-related |
|
OI10G06 |
Bo6g095520 |
Uncharacterized |
Unknown protein |
Putative uncharacterized protein |
|
Bo6g095530 |
Protein argonaute 7-like |
Argonaute family protein |
Protein argonaute |
References
- Bain D. 1952. Reaction of brassica seedlings to blackrot. Phytopathology. 42: 316-319.
- Camargo L, Williams P, Osborn T. 1995. Mapping of quantitative trait loci controlling resistance of Brassica oleracea to Xanthomonas campestris pv. campestris in the field and greenhouse. Phytopathology. 85: 1296-1300.
- Cheng X, Xu J, Xia S, Gu J, Yang Y, Fu J, et al. 2009. Development and genetic mapping of microsatellite markers from genome survey sequences in Brassica napus. Theor Appl Genet. 118: 1121-113.
- Chidamba L, Bezuidenhout CC. 2012. Characterisation of Xanthomonas campestris pv. campestris isolates from South Africa using genomic DNA fingerprinting and pathogenicity tests. Eur J Plant Pathol. 133: 811-818.
- Cook A, Walker J, Larson R. 1952. Studies on the disease cycle of black rot of crucifers. Phytopathology. 42: 162-167.
- Cruz J, Tenreiro R, Cruz L. 2017. Assessment of diversity of Xanthomonas campestris pathovars affecting cruciferous plants in portugal and disclosure of two novel X. campestris pv. campestris races. J Plant Pathol. 99: 403-414.
- Dangl JL, Jones JD. 2001. Plant pathogens and integrated defence responses to infection. Nature. 411: 826-833.
- Dickson M, Hunter J. 1987. Inheritance of resistance in cabbage seedlings to black rot. HortScience. 22: 108-109.
- Fargier E, Manceau C. 2007. Pathogenicity assays restrict the species Xanthomonas campestris into three pathovars and reveal nine races within X. campestris pvcampestris. Plant Pathol. 56: 805-818.
- Ferreira M, Dias J, Mengistu A, Williams P. 1992. Screening of Portuguese cole landraces (Brassica oleracea L.) with Leptosphaeria maculans and Xanthomonas campestris pv. campestris. Euphytica. 65: 219-227.
- Flor HH. 1971. Current status of the gene-for-gene concept. Annu Rev Phytopathol. 9: 275-296.
- Guo H, Dickson M, Hunter J. 1991. Brassica napus sources of resistance to black rot in crucifers and inheritance of resistance. HortScience. 26: 1545-1547.
- Ignatov A, Kuginuki Y, Hidam K. 2000. Distribution and inheritance of race-specific resistance to Xanthomonas campestris pv. campestris in Brassica rapa and B napus. J Russ Phytopathol Soc. 1: 89-94.
- Ignatov A, Hida K, Kuginuki Y. 1999. Pathotypes of Xanthomonas campestris pv. campestris in Japan. Acta Phytopathol Entomol Hung. 34: 177-182.
- Ignatov A, Vicente J, Conway J, Roberts S, Taylor J. 1997. Identification of Xanthomonas campestris pv. campestris races and sources of resistance. ISHS Symposium on Brassicas In: 10th Crucifer Genetics Workshop; pp 23-27.
- Ignatov A, Kuginuki Y, Hida K. 1998. Race-specific reaction of resistance to black rot in Brassica oleracea. Eur J Plant Pathol. 104: 821-827.
- Izzah NK, Lee J, Jayakodi M, Perumal S, Jin M, Park B, et al. 2014. Transcriptome sequencing of two parental lines of cabbage (Brassica oleracea L. var. capitata L.) and construction of an EST-based genetic map. BMC Genomics. 15: 149
- Jensen BD, Vicente JG, Manandhar HK, Roberts SJ. 2010. Occurrence and diversity of Xanthomonas campestris pv. campestris in vegetable Brassica fields in Nepal. Plant Dis. 94: 298-305.
- Kamoun S, Kamdar HV, Tola E, Kado CI. 1992. Incompatible interactions between crucifers and Xanthomonas compestris involve a vascular hypersensitive response: role of the hrpK locus. Mol Plant-Microbe Interact. 5: 22-33.
- Kifuji Y, Hanzawa H, Terasawa Y, Nishio T. 2013. QTL analysis of black rot resistance in cabbage using newly developed EST-SNP markers. Euphytica. 190: 289-295.
- King EO, Ward MK, Raney DE. 1954. Two simple media for the demonstration of pyocyanin and fluorescin. J Lab Clin Med. 44: 301-307.
- Lee J, Izzah NK, Jayakodi M, Perumal S, Joh HJ, Lee HJ, et al. 2015. Genome-wide SNP identification and QTL mapping for black rot resistance in cabbage. BMC Plant Biol. 15: 32
- Lema M, Soengas P, Velasco P, Francisco M, Cartea M. 2011. Identification of sources of resistance to Xanthomonas campestris pv. campestris in Brassica napus crops. Plant Dis. 95: 292-297.
- Lema M, Cartea ME, Francisco M, Velasco P, Soengas P. 2015. Screening for resistance to black rot in a Spanish collection of Brassica rapa. Plant Breeding. 134: 551-556.
- Lema M, Cartea ME, Sotelo T, Velasco P, Soengas P. 2012. Discrimination of Xanthomonas campestris pv. campestris races among strains from northwestern Spain by Brassica spp. genotypes and rep-PCR. Eur J Plant Pathol. 133: 159-169.
- Li H, Chen X, Yang Y, Xu J, Gu J, Fu J, et al. 2011. Development and genetic mapping of microsatellite markers from whole genome shotgun sequences in Brassica oleracea. Mol Breed. 28: 585-596.
- Massomo SM, Nielsen H, Mabagala RB, Mansfeld-Giese K, Hockenhull J, Mortensen CN. 2003. Identification and characterisation of Xanthomonas campestris pv. campestris strains from Tanzania by pathogenicity tests, biolog, rep-PCR and fatty acid methyl ester analysis. Eur J Plant Pathol. 109: 775-789.
- Roberts S, Hiltunen L, Hunter P, Brough J. 1999. Transmission from seed to seedling and secondary spread of Xanthomonas campestris pv. campestris in Brassica transplants: effects of dose and watering regime. Eur J Plant Pathol. 105: 879-889.
- Rouhrazi K, Khodakaramian G. 2014. Genetic fingerprinting of Iranian Xanthomonas campestris pv. campestris strains inducing black rot disease of crucifers. Eur J Plant Pathol. 139: 175-184.
- Saha P, Kalia P, Sonah H, Sharma TR. 2014. Molecular mapping of black rot resistance locus Xca1bo on chromosome 3 in Indian cauliflower (Brassica oleracea var. botrytis L.). Plant Breeding. 133: 268-274.
- Schaad N, Dianese J. 1981. Cruciferous weeds as sources of inoculum of Xanthomonas campestris in black rot of crucifers. Phytopathology. 71: 1215-1220.
- Sharma BB, Kalia P, Singh D, Sharma TR. 2017. Introgression of black rot resistance from Brassica carinata to cauliflower (Brassica oleracea botrytis Group) through embryo rescue. Front Plant Sci. 8: 1255
- Sharma BB, Kalia P, Yadava DK, Singh D, Sharma TR. 2016. Genetics and molecular mapping of black rot resistance locus Xca1bc on chromosome B-7 in Ethiopian mustard (Brassica carinata A. Braun.). PLoS ONE. 11: e0152290
- Song E, Kim S, Noh T, Cho H, Chae S, Lee B. 2014. PCR-based assay for rapid and specific detection of the new Xanthomonas oryzae pv. oryzae K3a race using an AFLP-derived marker. J Microbiol Biotechnol. 24: 732-739.
- Taylor J, Conway J, Roberts S, Astley D, Vicente JG. 2002. Sources and origin of resistance to Xanthomonas campestris pv. campestris in Brassica genomes. Phytopathology. 92: 105-111.
- Tonu NN, Doullah MA, Shimizu M, Karim MM, Kawanabe T, Fujimoto R, et al. 2013. Comparison of Positions of QTLs Conferring Resistance to Xanthomonas campestris pv. campestris in Brassica oleracea. Am J of Plant Sci. 4: 11-20.
- Van der Biezen EA, Jones JD. 1998. Plant disease-resistance proteins and the gene-for-gene concept. Trends Biochem Sci. 23: 454-456.
- Vicente JG, Holub EB. 2013. Xanthomonas campestris pv. campestris (cause of black rot of crucifers) in the genomic era is still a worldwide threat to Brassica crops. Molecular Plant Pathology. 14: 2-18.
- Vicente JG, Taylor J, Sharpe A, Parkin I, Lydiate D, King G. 2002. Inheritance of race-specific resistance to Xanthomonas campestris pv. campestris in Brassica genomes. Phytopathology. 92: 1134-1141.
- Vicente JG, Conway J, Roberts S, Taylor J. 2001. Identification and origin of Xanthomonas campestris pv. campestris races and related pathovars. Phytopathology. 91: 492-499.
- . 1980. Black rot: a continuing threat to world crucifers. Plant Dis. 64: 736-742.
- Williams P, Staub T, Sutton J. 1972. Inheritance of resistance in cabbage to black rot. Phytopathology. 62: 247-252.