Abstract
The application of molecular markers in rice breeding facilitates the rapid screening of genotypes in early growth stages without phenotypic assessment. In the present study, we developed and validated high throughput Kompetitive Allele Specific PCR (KASP) assays for rice stripe virus (RSV) resistance genes. The newly developed RSV-KASP markers were compared with the gel-based InDel marker, Indel7. The results of the RSV-KASP assay and the Indel7 analysis were consistent. Due to their high accuracy, time saving attribute, high throughput features, and cost-effectiveness, KASP could be more suitable for RSV genotyping than other methods.
-
Key words: Rice stripe virus, KASP
INTRODUCTION
Rice stripe virus (RSV) is one of the most devastating rice (
Oryza sativa L.) pathogens in growing regions in East Asia including Korea (
Chung 1974), causing RSV disease, with subsequent significant declines in yield and quality. It is a persistent plant virus that relies mostly on insects for its transmission. RSV is transmitted by the small brown plant-hopper (SBPH) (
Laodelphax striatellus Fallen) and the SBPH isolate influences the infection rate of the virus (
Falk and Tsai 1998;
Cho et al. 2013). In Korea, over the last two decades, the regional hotspot of rice stripe virus (genus
Tenuivirus) has shifted from the southeastern coastal regions to the western coastal regions based on recent shifts in temperature trends in the spring season (
Lee et al. 2017).
The screening method used commonly in many Korean rice breeding programs to evaluate RSV-resistant rice varieties consists of inoculating plants with the pathogen mediated by SBPH (
Zhao et al. 2016), and then observing symptom development over time. However, the method requires specialized skills and experience for the assessment of disease severity based on symptom development alone in different rice varieties exhibiting diverse symptoms.
According to several reports, the use of molecular marker-assisted selection (MAS), associated with conventional breeding approaches, facilitates the identification of genotypes by breeders, with increased accuracy in determining genotypes that are associated with different economically important traits, which can improve rice breeding efficiency markedly (
Kwon et al. 2012a;
Cho et al. 2018).
Stv-a and Stv-b, have been reported to be loci related to the resistance to RSV in Japanese upland rice (Washio et al. 1968). Stv-a is located on chromosome 6, whereas Stv-b is located on chromosome 11.
Beginning in the 1960s,
Stv-bi was introduced by backcrossing from the indica paddy variety Modan to the
japonica paddy variety in Japan.
Stv-bi was subsequently used for breeding japonica rice varieties in Korea and Japan (
Kwon et al. 2012a).
The Indel7 marker developed by
Kwon et al. (2012a) in a previous study is a
Stv-bi gene linked marker located at 17.95 Mb on chromosome 11. The InDel7 marker is a co-dominant marker with a genotype and phenotype matching rate of 96.2%.
Recently, the application of Kompetitive Allele-Specific PCR (KASP), a detection method that can type single nucleotide polymorphisms (SNPs) and insertion-deletion mutations (InDels) at specific sites, has increased gradually in plant breeding programs. With respect to genotyping, KASP is based on terminal fluorescence reading, similar to the basis of
TaqMan probe detection. KASP-SNP markers are highly accurate, cost-effective, and highly flexible. The markers also have a high conversion rate and a wide range of applications. Several functional SNPs and InDels have been converted into functional markers (FMs) using KASP assays, which greatly improve the speed and efficiency of selection in plant breeding programs (
Neelam et al. 2013;
Rasheed et al. 2016).
Therefore, the aim of the current study was to convert the Indel7 marker linked to the RSV resistance gene into a more efficient KASP marker and facilitate their application in breeding programs through marker assisted selection (MAS).
MATERIALS AND METHODS
Plant materials
Two hundred and sixty-three Korean
japonica rice varieties, and 94 F
2 and 35 BC
1F
1 plants derived from crosses between Ilpum (P
1) and Saeilmi (P
2), the susceptible and resistant parental genotypes, respectively, carrying
Stv-bi originating from Modan (
Kwon et al. 2012b), a Sri-Lankan rice variety, were used in the present study to validate the gene-specific KASP markers for RSV-resistance.
We targeted the InDels between the resistant and susceptible haplotypes to develop RSV-specific KASP markers. The KASP marker, RSV-KASP, was designed at the target InDel using pipeline of LGC Genomics (LGC Ltd, Teddington, UK) (
https://www.biosearchtech.com). The genomic DNA was amplified using primers of two allele-specific forward primers and two RSV-KASP reverse primers (
Fig. 1). The list of primer sequences for KASP and InDel markers for RSV resistance gene, in addition to common primers used in the present study, are listed in
Table 1. The reaction volume of the 10-mL KASP assay consisted of 5 mL KASP Master mix, 0.14 mL KASP primer assay mix, and 5 mL of 10 ng/mL DNA template. The ABI 7500 FAST real-time PCR system (Applied Biosystems Inc.) was used to amplify and detect the fluorescent end-point reading. The following thermal cycling conditions were used: 94℃ for 15 minutes, 10 touchdown cycles of 94℃ for 20 seconds and 61-55℃ (dropping 0.7℃ per cycle) for 1 minute, followed by 26 cycles of 94℃ for 20 seconds and 55℃ for 1 minute. The fluorescent end-point reading was performed at 30℃ for 1 minute. The allele discrimination mode of 7500 v2.3 was used for genotype calling.
RESULTS AND DISCUSSION
An Indel7 marker closely linked to RSV-resistance gene has been previously developed for the detection of genotypes harboring
Stv-bi (
Kown et al. 2012a,
b). In the present study, the reliability of the KASP genotyping results was confirmed using InDel7 marker analysis (
Kwon et al. 2012b). The location of RSV resistance gene was detected on chromosome 11 (
Kwon et al. 2012a). The detection of RSV resistance gene in rice was carried out with a high throughput marker designed based on KASP reaction principles and the single base difference of the anti-sense material (
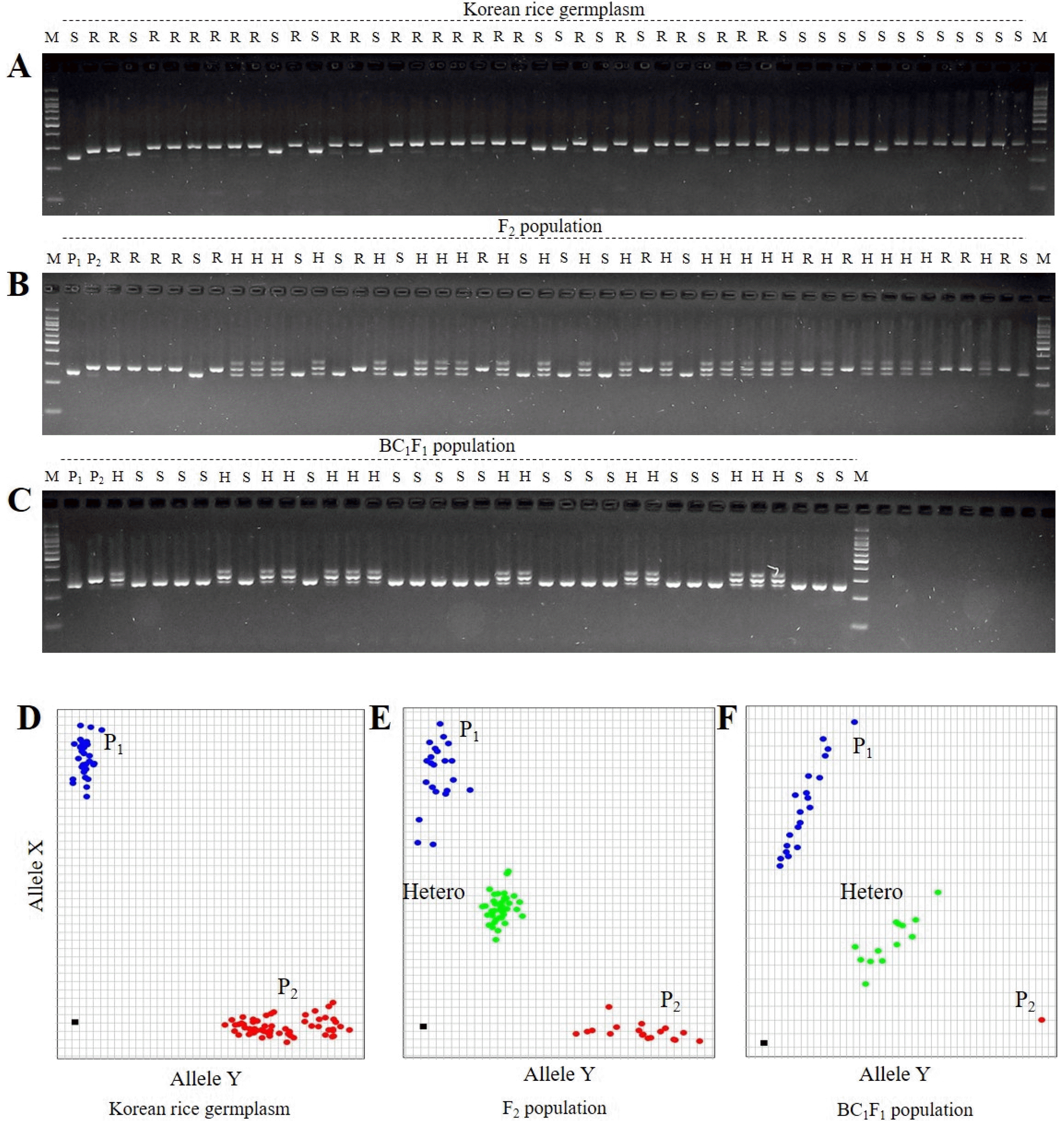
Fig. 1). According to the KASP genotyping results, the screened Korean varieties showed variability among them, showing either the resistance allele or the susceptible allele (
Fig. 2D). The comparison of resistant (R) and susceptible (S) genotypes based on allele-specific KASP-markers and the InDel-based markers, InDel7, closely linked to the RSV resistance gene, is illustrated in
Fig. 2A-C. The results show the separation of different genotypes, and the dots indicating genomic positions of each marker in each rice genotype were closely assembled. Therefore, the newly developed RSV-KASP marker could detect rice
Stv-bi genotypes.
The RSV-KASP assay accurately clustered genotypes into resistant and susceptible haplotype in the panel of 263 Korean
japonica rice varieties. The 263
japonica rice varieties were further classified using RSV-KASP markers. In early maturing varieties, 17 varieties were resistant, and 67 varieties were sensitive. In mid-maturing varieties, 52 varieties were resistant and 25 varieties were susceptible. In the mid–late maturing varieties, 76 varieties were resistant and 26 were susceptible (
Table 2). The RSV-KASP marker has been extensively applied in numerous breeding programs. In the F
2 population, the genotyping results revealed heterozygous and homozygous alleles of parents (
Fig. 2E), suggesting that the newly developed RSV-KASP marker could be used for MAS in early generations. Furthermore, RSV-KASP genotyping results of BC
1F
1 plants illustrated in
Fig. 2F indicate that the population was well separated into susceptible and heterozygous alleles, suggesting that RSV-KASP would be suitable for application in backcross breeding programs in rice. Considering the above results, RSV-KASP genotyping could be used to determine RSV resistance in rice in early generations.
RSV-KASP markers were developed for the selection of RSV resistance, targeting the InDel7 region in resistant and susceptible haplotypes, to facilitate the efficient and accurate genotyping of RSV resistance in rice. We used RSV-KASP markers to detect RSV resistant genotypes in F2 and BC1F1 segregating rice populations. In addition, RSV-KASP markers facilitated the detection of homozygous and heterozygous alleles in parents. Therefore, RSV-KASP genotyping could be applied in MAS of the RSV resistance gene in rice.
ACKNOWLEDGEMENTS
This work was supported by a grant from the Next-Generation Bio Green 21 Program (No. PJ01319602) and Rural Development Administration, Republic of Korea.
Fig. 1Sequence alignment in Indel7 region on chromosome 11 in RSV-resistant and RSV-susceptible genotypes. Their positions are indicated above the sequences. Arrows indicate the locations of the resistant allele (Third row Allele X and Reverse 1) and susceptible allele (Fourth row Allele Y) primers, respectively.

Fig. 2Comparison of genotypes between RSV-KASP marker and InDel marker Indel7 closely linked to RSV. (A) Genotype of Indel7 in 263 Korean japonica rice varieties visualized on 3% agarose gel after mixing. (B) F2 plants derived from a cross between Ilpum (P1) and Saeilmi (P2). (C) BC1F1 population derived from a cross of Ilpum and Saeilmi for marker assisted backcross. (D-F) Plots of R/S-specific RSV-KASP marker-PCR products for MAS of RSV. MAS: marker assisted selection, RSV: Rice stripe virus, R: Resistance, S: susceptible.
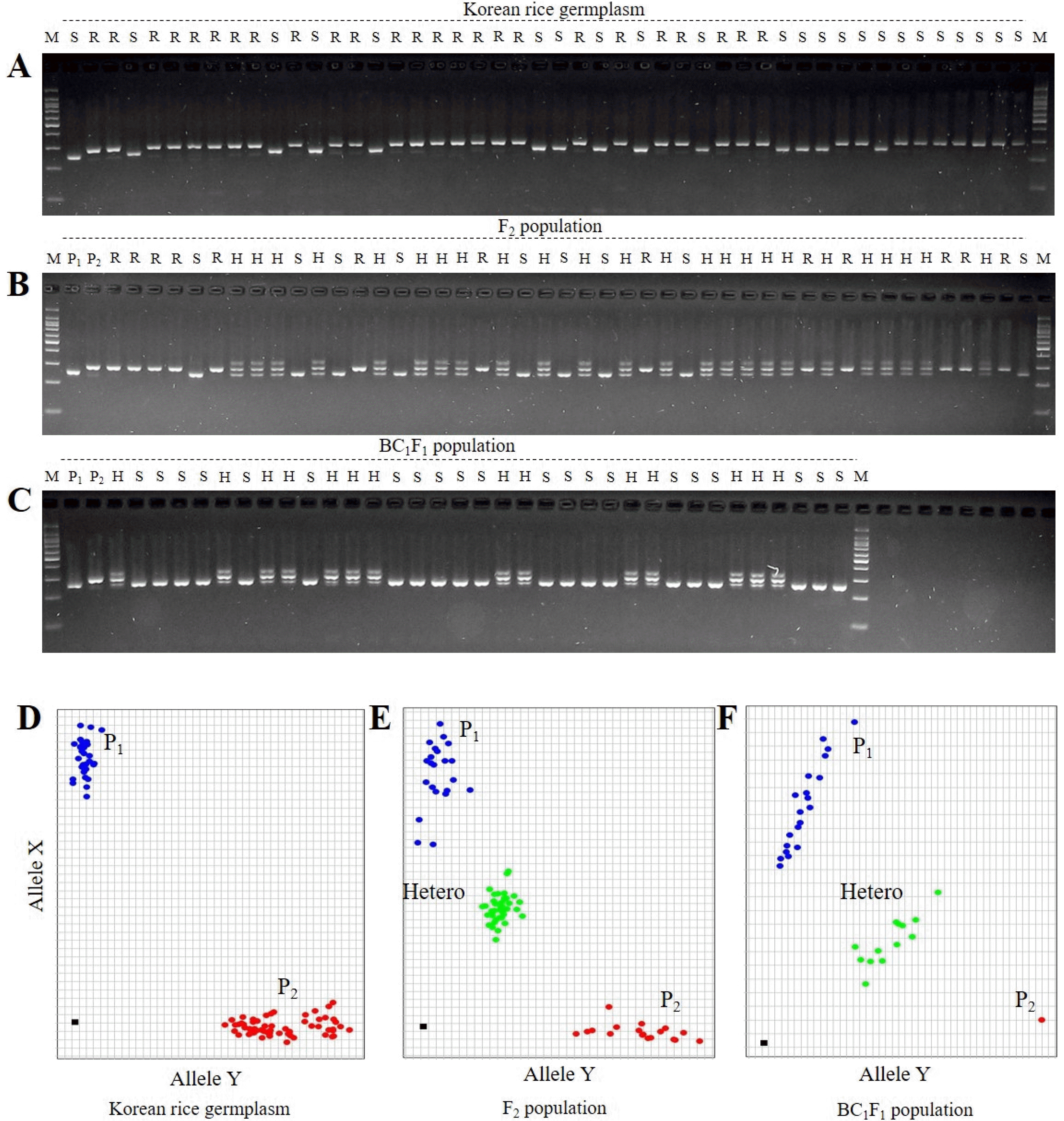
Table 1List of primer sequences used in the present study.
Table 1
|
Marker |
Primers |
Primer sequences (5ʹ to 3ʹ) |
SNP |
|
|
KASP |
Allele X |
AACGTGTAGTTGCCCGGCATC |
C |
Resistance |
|
Allele Y |
AACGTGTAGTTGCCCGGCATG |
G |
Susceptible |
|
Reverse 1 |
CAGCGAAATCCATGCCCTCGTAATT |
|
Reverse 2 |
CAGCGAAATCCATGCCCTCGTAATT |
|
InDel |
Indel7-F |
AAA TTC CAG TGC CCA AAA CC |
, Kwon et al. (2012)
|
|
Indel7-R |
TCC TCA TGG AGC TCA TCC AAG |
Table 2Classification of resistant and susceptible varieties using KASP marker for RSV resistance gene.
Table 2
|
Ecotype |
Resistant |
Susceptible |
|
Early maturing |
Danpyeng, Haedamssal, Hwawang, IS592BB, Jinkwang, Jogwang, Joil, Jonong, Joryeong, Jungmo1032, Jungmo1043, Keumo3, Manan, Pyeongwon, Sanhomi, Shinpyeong, Jopyeong (17 cultivars) |
Asemi, Asemi1, Baegilmi, BoSeog, Cheongbaekchal, CW92MR, Dunnae, Geumyoung, Goun, Guru, Haedeul, Handeul, Hanseol, Heugjinju, Hwangkeumbora, Jeogjinju, Jeogjinjuchal, Jinbu, Jinbuchal, Jinbuol, Jinhan, Jinmi, Jinok, Jinseolchal, Joami, Joan, Joeunheukmi, Jopum, Josaegheugchal, Joun, Junamjosaeng, Junghwa, Jungmo1024, Jungsan, Keumo, Manchoo, Manho, Manna, Namil, Namwon, Nukeunheugchal, Nunkeunheugchal1, Obong, Odae, Odae1, Ondami, Saeodae, Saesangju, Samcheon, Sandeuljinmi, Sangju, Sangjuchal, Seolbaek, Seolemi, Seongsan, Sinunbong, Sinunbong1, Sobaeg, Taebong, Unbaekchal, Unbong, Undoo, Unilchal, Unjang, Unkwang, Unmi, Weolbada (67 cultivars) |
|
Mid maturing |
Bodrami, Borami, Boseogchal, Cheongdam, Cheongnam, Cheongpum, Dabo, Daebo, Daepyeong, Dongbo, Donghaejinmi, Gancheok, Gangchan, Geonyang2, Haechanmulgyeol, Haeoreumi, Haepum, Haepyeong, Haepyeongchal, Heugkwang, Hwaan, Hwabong, Hwajin, Hwanong, Hwanseonchal, Hwaseong, Hwayeong, Janngan, Jungmo1034, Jungsaenggold, Keumo2, Keumobyeo 1, Manjong, Manpung, Manwol, Palgong, Pungmi, Samdeog, Samkwang1, Sampyeong, Sangbo, Sangnambatbyeo, Seohyangheukmi, Seonpum, Shinbaeg, Sinseonchal, Sobi, Suan, Suryeonjinmi, Yeongan, Yeongdeog, Youngbo (52 cultivars) |
Giho, Boseogheugchal, Cheonga, Cheongan, Daejin, Daeripbyeo 1, Dodamssal, Gangbaek, Geuman, Geunnun, Heugseol, Hongjinju, Jinpum, Juan, Migwang, Naepung, Nongan, Nunbora, Saegyehwa, Seoan, Seolhyangchal, Sinbo, Sura, Goami3, Seoan1 (25 cultivars) |
|
Mid late maturing |
Anbaek, Anmi, Aromi, Baekogchal, Boramchal, Boramchan, Cheongcheongjinmi, Cheonghaejinmi, Cheongho, Cheonghyangheukmi, Cheongun, Chilbo, Chindeul, Chinnong, Dacheong, Daean, Daecheong, Deuraechan, Dongan, Dongjin, Dongjin2, Geongganghongmi, Goami, Haiami, Hanam, Heughyang, Heugsujeong, Hoan, Hojin, Honong, Hopum, Huimangchan, Hwanam, Hwanggeummodeul, Hwangkeumnuri, Hwangrang, Hwansam, Hwasin, Hyeopum, Ilmil, Jeongjinju2, Jinbaek, Jinbo, Jinsumi, Junam, Jungmo1006, Keunpum, Kyehwa, Malgeumi, Manbaek, Manguem, Miho, Mihyang, Mipum, Misomi, MY298BB, MY299BK, Nanpyeong, Onnuri, Pungmi1, Saechilbo, Saeilmi, Saenuri, Saeshin, Samkwang, Seomyeong, Sindongjin, Sinjinbaek, Sodami, Sujin, Sukwang, Tamjin, Yangjo, Yechan, Younghojinmi, Youngjin (76 cultivars) |
Aranghangchal, Baegjinju, Baegjinju1, Baegseolchal, Daesan, Dami, danmi, Dongjin1, Dongjinchal, Geonyangmi, Goami2, Goami4, Gopum, Hangaru, Hanmaeum, Heugjinmi, Heugnam, Hopyung, Hyangnam, Ilpum, Manmi, Misiru, Saegoami, Saeilpum, Seolgaeng, Seopyeong(26 cultivars) |
References
- Cho WK, Lian S, Kim SM, Park S, Kim KH. 2013. Current insights into research on Rice stripe virus. Plant Pathol. J.. 29: 223-233.
- Cho JH, Lee JY, Kang JW, Kwon YH, Choi IR, Song YC, et al. 2018. Development of allele specific SNP marker for RTSV resistance gene at the tsv locus. Plant Breed. Biotech.. 6: 309-312.
- Chung BJ. 1974. Studies on the occurrence, host range, transmission, and control of rice stripe disease in Korea. Korean J. Appl. Entomol.. 13: 181-204.
- Falk BW, Tsai JH. 1998. Biology and molecular biology of viruses in the genus Tenuivirus. Annu. Rev. Phytopathol.. 36: 139-163.
- Hayano-Saito Y, Saito K, Hujii K, Touyama T, Tsuji T, Sugiura N, et al. 2000a. SCAR marker for selection of the rice stripe resistance gene Stvb-i. Breed. Res.. 2: 67-72.
- Hayano-Saito Y, Saito K, Nakamura S, Kawasaki S, Iwasaki M. 2000b. Fine physical mapping of the rice stripe resistance gene locus, Stvb-i. Theor. Appl. Genet.. 101: 59-63.
- Kwon TM, Lee JH, Park SK, Hwang UH, Cho JH, Kwak DY, et al. 2012a. Fine mapping and identification of candidate rice genes associated with qSTV11SG, a major QTL for rice stripe disease resistance. Theor. Appl. Genet.. 125: 1033-1046.
- Kwon TM, Lee JH, LeeSB , Park SK, Kim SY, Hwang UH, et al. 2012b. Screening of rice stripe resistance germplasm to develop new resistance allele in rice. Korean J. Breed. Sci.. 44: 282-289.
- Lee M, Lee S, Park JW, Son J, Lee DH, Ryoo M, et al. 2017. Prevalence of rice stripe virus can be altered by temperature and the virus-mediated development of insect vector, Laodelphax striatellus, in Korea. J. Asia-Pac. Entomol.. 20: 1145-1149.
- Neelam K, Brown-Guedira G, Huang L. 2013. Development and validation of a breeder-friendly KASPar marker for wheat leaf rust resistance locus Lr21. Mol. Breed.. 31: 233-237.
- Rasheed A, Wen W, Gao F, Zhai S, Jin H, Liu J, et al. 2016. Development and validation of KASP assays for genes underpinning key economic traits in bread wheat. Theor. Appl. Genet.. 129: 1843-1860.
- Zhao W, Yang P, Kang L, Cui F. 2016. Different pathogenicities of Rice stripe virus from the insect vector and from viruliferous plants. New Phytol.. 210: 196-207.
Citations
Citations to this article as recorded by

- Genome-Wide Characterization of the PaO Gene Family and Pyramiding Effects of Superior Haplotypes on Yield-Related Traits in Sorghum
Jinbiao Li, Haoxiang Li, Ruochen Zhang, Yizhong Zhang, Juanying Zhao, Xiaojuan Zhang, Huiyan Wang
Agronomy.2025; 15(11): 2493. CrossRef - KASP: a high-throughput genotyping system and its applications in major crop plants for biotic and abiotic stress tolerance
Bhawna Dipta, Salej Sood, Vikas Mangal, Vinay Bhardwaj, Ajay Kumar Thakur, Vinod Kumar, Brajesh Singh
Molecular Biology Reports.2024;[Epub] CrossRef - Pyramiding effects of favorable haplotypes of loci on major fiber yield and quality traits in Upland Cotton (Gossypium hirsutum L.)
Yingrui Zhao, Baojun Chen, Hongge Li, Jingjing Wang, Yinhua Jia, Zhaoe Pan, Daowu Hu, Zhen Peng, Yingxiao Li, Xu Gao, Peng Zhang, Liru Wang, Jun Peng, Shoupu He, Du Xiongming
Industrial Crops and Products.2024; 217: 118805. CrossRef - KASP mapping of QTLs for yield components using a RIL population in Basmati rice (Oryza sativa L.)
Hamza Ashfaq, Reena Rani, Naila Perveen, Allah Ditta Babar, Umer Maqsood, Muhammad Asif, Katherine A. Steele, Muhammad Arif
Euphytica.2023;[Epub] CrossRef - Comparative Analysis between the ITS TaqMan SNP Genotyping Assay and CAPS for the Rapid Molecular Identification of Zoysia japonica and Zoysia sinica, Related Hybrid Lines, and the Habitat Distribution of Each Species
Dae-Hwa Yang, Ok-Cheol Jeong, Yu-Ryang Kim, Mi-Jeong Kang, Yang-Ji Kim, Ji-Hi Son, Seong-Seop Han, Mi-Young Park, Il-Doo Jin, In-Ja Song, Min-Ji Hong, Hyeon-Jin Sun, Hong-Gyu Kang, Hyo-Yeon Lee
Horticultural Science and Technology.2023; 41(4): 463. CrossRef - Evaluation of Major Rice Varieties for Bakanae Disease Resistance in Korea
Sais-Beul Lee, Ju-Won Kang, Ji-Yoon Lee, Gi-Un Seong, Youngho Kwon, So-Myeong Lee, Nkulu Rolly Kabang, Jun-Hyeon Cho, Seong-Hwan Oh, Dongjin Shin, Jong-Hee Lee, Ki-Won Oh, Dong-Soo Park
Korean Journal of Breeding Science.2023; 55(2): 103. CrossRef - New Transcriptome-Based SNP Markers for Noug (Guizotia abyssinica) and Their Conversion to KASP Markers for Population Genetics Analyses
Sewalem Tsehay, Rodomiro Ortiz, Eva Johansson, Endashaw Bekele, Kassahun Tesfaye, Cecilia Hammenhag, Mulatu Geleta
Genes.2020; 11(11): 1373. CrossRef - Validation and Selection of Functional Allele-specific Molecular Markers to Analyze High-Molecular-Weight Glutenin Subunit Composition in Wheat
Dongjin Shin, Jin-Kyung Cha, So-Myeong Lee, Jong-Min Ko, Jong-Hee Lee
Korean Journal of Breeding Science.2020; 52(3): 235. CrossRef