Abstract
In this study, in order to understand the differentiation process of Perilla crop and related weedy types collected from South Korea, morphological variation between accessions of cultivated var. frutescens and related weedy types of var. frutescens and var. crispa was investigated by principal component analysis (PCA) using morphological characteristics, especially including seed traits such as seed size, seed hardness, seed color and seed germination rate. The first and second principal components accounted for 54.1% and 17.9% of the total variance, respectively. In the PCA analysis, most of the qualitative and quantitative traits contributed significantly to the positive or negative direction on the first axis. Thus, the first axis could be used mainly to distinguish between accessions of cultivated var. frutescens and weedy var. frutescens, and also between accessions of cultivated and weedy types of var. frutescens and weedy var. crispa. However, for several accessions it was not possible to discriminate clearly between accessions of cultivated and weedy types of var. frutescens and also between accessions of the two weedy types of var. frutescens and var. crispa. The results of the PCA analysis are thought to provide useful information for understanding the cultivation process of Perilla crop and the differentiation process of Perilla crop and related weedy types. Also, this study demonstrates the efficacy and utility of PCA analysis using morphological traits, including seed traits such as seed size, seed hardness, seed color and seed germination rate, in the study of morphological variation of Perilla crop and related weedy types.
-
Key words: Perilla crop, Cultivated and weedy types, Cultivation process, Seed characteristics, Principal component analysis
INTRODUCTION
Perilla crop (
Perilla frutescens L.) belongs to family Lamiaceae, and is widely cultivated and used mainly in East Asia countries.
Perilla crop is classified into two varieties (or cultivated types):
Perilla frutescens var.
frutescens and
Perilla frutescens var.
crispa in accordance with their morphological characteristics and usage conditions (
Makino 1961). The two cultivated types of
Perilla crop have a long history of cultivation in East Asia (
Li 1969;
Lee and Ohnishi 2001,
2003;
Nitta et al. 2003) where the cultivated var.
frutescens is used as both a leafy vegetable and an oil crop, while the cultivated var.
crispa is used as both a leafy vegetable and Chinese medicine crop (
Lee and Ohnishi 2001,
2003;
Nitta 2001;
Nitta et al. 2003). In South Korea, var.
frutescens is currently grown in many areas, whereas var.
crispa is often found growing as a weed in waste areas and roadsides and around farmers’ fields or houses (
Lee and Ohnishi 2001;
Lee et al. 2002;
Lee et al. 2002,
2015). The two cultivated types of
Perilla crop differ in some morphological characteristics. Cultivated var.
frutescens is taller and has larger seed size (>2 mm) than cultivated var.
crispa. The former shows soft or hard seeds, green leaves and stems, and non-wrinkled leaves; while the latter shows only hard and small seeds (seed size < 2 mm), exhibits red or green coloration on the leaves and stems, and has wrinkly or non-wrinkly leaves (
Makino 1961;
Lee and Ohnishi 2001;
Nitta 2001;
Nitta et al. 2003).
On the other hand, weedy plants of
Perilla crop were firstly reported in the two cultivated types of
Perilla crop in East Asia by
Lee and Ohnishi (2001,
2003),
Lee et al. (2002),
Nitta (2001),
Nitta and Ohnishi (1999), and
Nitta et al. (2003). Recently, many researchers have reported weedy plants of
Perilla crop in South Korea and China. For example, weedy plants of
Perilla crop grow naturally and are common found in habitats such as roadsides and wastelands, near rivers or streams, and around farmers’ fields or farmers’ houses in South Korea and China (
Lee and Ohnishi 2001,
2003;
Nitta et al. 2003,
2005;
Lee et al. 2002,
2015, 2018a;
Ma and Lee 2017;
Ma et al. 2017,
2019). These weedy plants exhibit green leaves and stems and the same fragrance as cultivate var.
frutescens (or little fragrance), but their seeds are smaller and harder than those of cultivate var.
frutescens (
Lee and Ohnishi 2001;
Sa et al. 2012,
2013;
Ma and Lee 2017). However, in southern and northern China, weedy plants of
Perilla crop grow wild (
Nitta et al. 2003,
2005;
Ma and Lee 2017;
Ma et al. 2017,
2019).
Wild species of the two cultivated types of
Perilla crop are unknown in East Asia, but it is necessary to understand the cultivation process of the two cultivated types of
Perilla crop and also the difference between
Perilla crop and related weedy types. In a previous report by
Sa et al. (2018b), it was reported that most accessions of cultivated var.
frutescens showed a high germination rate of 70% or more, while all accessions of weedy var.
frutescens and cultivated and weedy types of var.
crispa showed low germination rates. In addition, accessions of cultivated var.
frutescens were divided into two types (type I and type II) based on their germination rates (high and low): most accessions of type I showed a high germination rate of 70% or more, while some accessions of type II showed a low germination rate of 20% or less (
Sa et al. 2018b). Therefore,
Perilla crop and related weedy types are thought to exhibit different variations in seed-related properties, such as seed size, seed hardness, seed color, and seed germination rate.
By this time, many researchers have been researched to distinguish between
Perilla crop and related weedy types. The two cultivated types of
Perilla crop and related weedy types have the same number of chromosomes (2n=40) (
Yamane 1950;
Honda et al. 1994;
Nitta 2001). Also, cross experiments between the two cultivated types of
Perilla crop by
Nagai (1935), Honda
et al. (1990, 1994), and (
Lim et al. 2019) showed that they are mating with each other by artificial pollination. Furthermore, research by
Koezuka et al. (1985,
1986),
Honda et al. (1990),
Lee and Ohnishi (2001), and
Ma and Lee (2017) were unable to distinguish between the two cultivated types of
Perilla crop due to the presence of intermediate types such as weedy plants. Therefore, they reported that essential oil, anthocyanin, seed color, seed hardness, and plant fragrance could not be used as key characters to distinguish between
Perilla crop and related weedy types. Most accessions of cultivated var.
crispa and its weed type were not clearly separated by principal component analyses (PCAs) using 22 qualitative and quantitative characters (
Lee and Ohnishi 2001). Thus, it is still difficult to distinguish between accessions of
Perilla crop and related weedy types using morphological characteristics, and there is a need to understand the morphological variations in the accessions of
Perilla crop and related weedy types in East Asia.
In this study, in order to understand the differentiation process of Perilla crop and related weedy types, mor-phological variations between accessions of cultivated var. frutescens and related weedy types of var. frutescens and var. crispa collected from South Korea were investigated by principal component analysis (PCA) using mor-phological characteristics, especially seed traits such as seed size, seed hardness, seed color and seed germination rate.
MATERIALS AND METHODS
Plant materials and Morphological characteristic survey
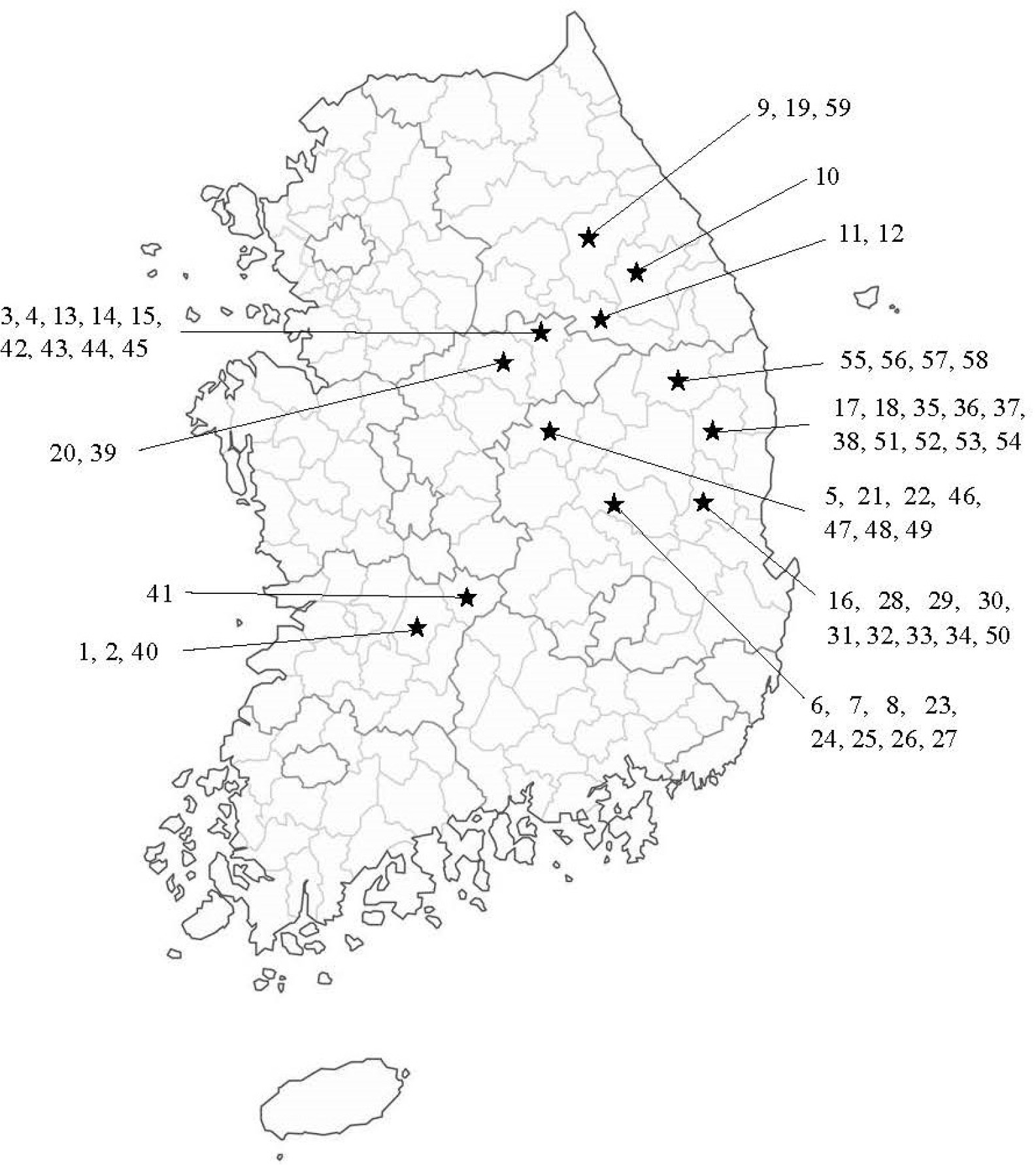
The materials for the present study consist of 59
Perilla accessions (19 cultivated var.
frutescens, 20 weedy var.
frutescens, 20 weedy var.
crispa) collected from South Korea. The number of accessions and the name of the collection place for the collected accessions of cultivated var.
frutescens and related weedy types of var.
frutescens and var.
crispa are shown in
Fig. 1 and
Table 1. In the case of germination experiments,
Sa et al. (2018b) completed a germination rate survey for the 59
Perilla accessions at one month after harvesting in the fall of 2017, and the experimental results reported by
Sa et al. (2018b) were used as the germination rate data of the 59
Perilla accessions (
Table 1).
Meanwhile, in our study, to evaluate the morphological variation of the 59
Perilla accessions that were used in the germination survey by
Sa et al. (2018b), in early May, 20 seeds of each
Perilla accession were sown in a seedling box and maintained in a glass greenhouse for a month. Ten seedling plants of each
Perilla accession were then transferred into the field in early June 2020 at the experimental farm of Kangwon National University, Chuncheon, Gangwon-do. In this study, we examined nine qualitative characters of each
Perilla accession, as shown in
Table 2.
Principal component analysis (PCA) was performed to detect any differences between and within the accessions of cultivated var.
frutescens and related weedy types of var.
frutescens and var.
crispa. Multivariate analysis was performed using the Microsoft Excel Statistical Analysis System Program and NTSYS-pc V2.1 Program (
Rohlf 1998). SPSS software was used to perform correlation analysis for one quantitative and nine qualitative characteristics.
RESULTS
Morphological difference between Perilla crop and related weedy types
The results for one quantitative and nine qualitative traits of 59 accessions of cultivated var.
frutescens and weedy types of var.
frutescens and var.
crispa are summarized in
Table 3.
For the one quantitative trait (seed germination rate, QN1), which was reported by
Sa et al. (2018b), 15 accessions of cultivated var.
frutescens showed a high germination rate of 70% or more and four accessions showed a low germination rate of less than 20%, ranging from 17 to 19%. All accessions of weedy var.
frutescens (20 accessions) and weedy var.
crispa (20 accessions) showed a low germination rate of less than 1%, ranging from 0 to 1%.
Among the nine qualitative traits, for seed size (QL1) all accessions of cultivated var. frutescens (19 accessions) had large seeds and all accessions of weedy var. frutescens (20 accessions) and var. crispa (20 accessions) had small seeds. For seed hardness (QL2), 18 accessions of cultivated var. frutescens had soft type and 1 accession had hard type, 1 accession of weedy var. frutescens had soft type and 19 accessions had hard type, and all 20 accessions of weedy var. crispa had hard type. For seed color (QL3), the accessions of cultivated var. frutescens showed white (1 accession), gray (4 accessions), light brown (10 accessions), and brown (4 accession) colors; the accessions of weedy var. frutescens showed only gray (4 accessions) and dark brown (16 accessions) colors; and the accessions of weedy var. crispa showed only light brown (1 accession) and dark brown (19 accessions) colors. For color of surface leaf (QL4), five accessions of cultivated var. frutescens showed light green color, 12 accessions showed green color, and 2 accessions showed deep green color. Meanwhile, four accessions of weedy var. frutescens showed light green color, 12 accessions showed green color, and four accessions showed deep green color; and the accessions of weedy var. crispa showed green (6 accessions), deep green (3 accessions), green/purple (7 accessions), and purple (4 accessions) colors. For color of the reverse side leaf (QL5), the accessions of cultivated var. frutescens showed light green (5 accessions) and green (14 accessions) colors; the accessions of weedy var. frutescens showed light green (8 accessions), green (9 accessions), and deep green (3 accessions) colors; and the accessions of weedy var. crispa showed green/purple (3 accession) and purple (17 ac-cessions) colors. For leaf size (QL6), the accessions of cultivated var. frutescens showed large (3 accessions), middle (14 accessions), and small (2 accessions) types; the accessions of weedy var. frutescens showed middle (15 accessions) and small (5 accessions) types; and the accessions of weedy var. crispa showed middle (17 accessions) and small (3 accessions) types. For the degree of pubescence (QL7), the accessions of cultivated var. frutescens showed slightly pubescent (7 accessions), pubescent (3 accessions), and heavily pubescent (9 accessions) degrees; the accessions of weedy var. frutescens showed slightly pubescent (4 accessions), pubescent (9 accessions), and heavily pubescent (7 accessions) degrees; and the accessions of weedy var. crispa showed only slightly pubescent (12 accessions) and pubescent (8 accessions) degrees. For stem color (QL8), the accessions of cultivated var. frutescens showed light green (4 accessions) and green (15 accessions) colors; the accessions of weedy var. frutescens showed light green (1 accession), green (17 accessions), and deep green (2 accessions) colors; and the accessions of weedy var. crispa showed purple (18 accessions) and green/purple (2 accessions) colors. For flower color (QL9), all accessions of cultivated (19 accessions) and weedy (20 accessions) types of var. frutescens showed only white color, while the accessions of weedy var. crispa showed white/purple (4 accessions) and purple (16 accessions) colors.
In this study, we performed a correlation analysis to detect the relationship between the one quantitative trait and nine qualitative traits in the 59 accessions of cultivated var.
frutescens and related weedy types (
Table 4). Among all combinations, the combinations between QN1 and QL1 (‒0.876**), between QN1 and QL2 (0.826**), between QL1 and QL2 (‒0.871**), between QL4 and QL5 (0.728**), and between QL5 and QL8 (0.895**) showed comparatively higher positive or negative correlation coefficients than the other combinations, with significance levels of 0.01. In addition, many combinations between the one quantitative trait and nine qualitative traits showed significant positive or negative correlations, with significance levels of 0.05 or 0.01. For example, in the case of traits related to seed characteristics, QN1 showed a significant negative correlation coefficient with QL1 (‒0.876**), QL4 (‒0.368**), QL5 (‒0.428**), QL6 (‒0.559**), and QL9 (‒0.550**) and a significant positive correlation coefficient with QL2 (0.826**), QL3 (0.445**), and QL7 (0.466**) at a significance level of 0.01 (
Table 4). QL1 showed a significant negative correlation coefficient with QL2 (‒0.871**), QL3 (‒0.584**), and QL7 (‒0.565**) and a significant positive correlation coefficient with QL4 (0.340**), QL5 (0.441**), QL6 (0.620**), and QL9 (0.568**) at a significance level of 0.01. QL2 showed a significant negative correlation coefficient with QL4 (‒0.410**), QL5 (‒0.491**), QL6 (‒0.611**), and QL9 (‒0.531**) and a significant positive correlation coefficient with QL3 (0.403**) and QL7 (0.583**) at a significance level of 0.01. QL3 showed a significant negative correlation coefficient with QL9 (‒0.261*) at a significance level of 0.05.
In the case of traits related to leaf characteristics, QL4 showed a significant positive correlation coefficient with QL5 (0.728**), QL6 (0.323*), QL8 (0.618**), and QL9 (0.517**) at a significance level of 0.01. QL5 showed a significant positive correlation coefficient with QL6 (0.409**), QL8 (0.895**), and QL9 (0.692**) at a sign-ificance level of 0.01. QL6 showed significant negative or positive correlation coefficients with QL7 (‒0.364**), QL8 (0.265*), and QL9 (0.463**) at a significance level of 0.05 or 0.01. In the case of traits related to plant characteristics, QL8 showed a significant positive correlation coefficient with QL9 (0.638**) at a significance level of 0.01. Among all combinations, in particular QN1 and QL1 (‒0.876**), QN1 and QL2 (0.826**), QL1 and QL2 (‒0.871**), QL4 and QL5 (0.728**), and QL5 and QL8 (0.895**) showed a very high correlation coefficient compared with the other traits at
P < 0.01 (
Table 4).
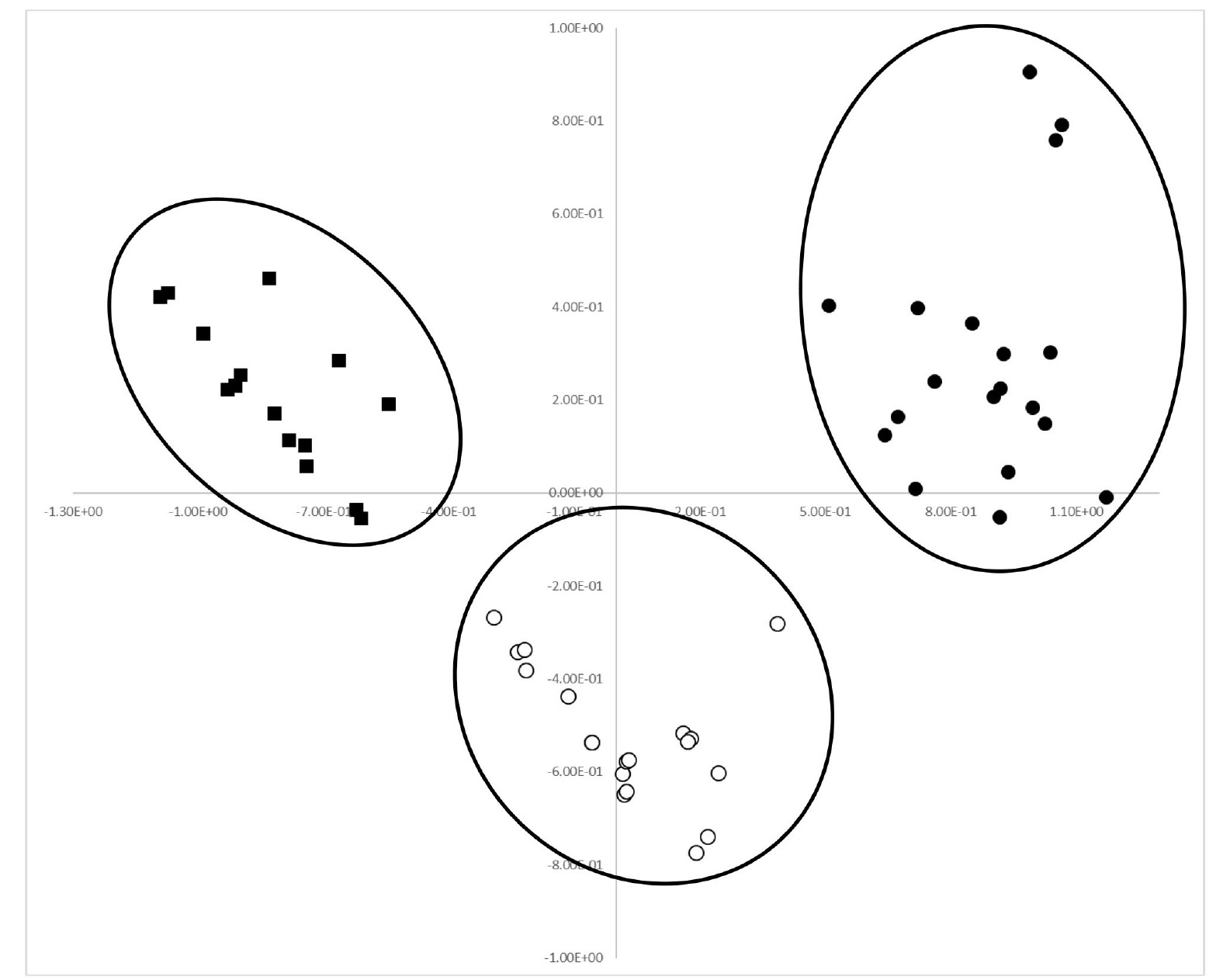
The first and second principal components were res-pectively accounted for 54.1% and 17.9% of the total variance (
Table 5). In PCA analysis, most of the qualitative and quantitative traits greatly contributed in the positive or negative direction on the first axis (
Table 5). In particular, those traits that are considered to be useful for discrimination between accessions of cultivated var.
frutescens and related weedy types contributed in the negative direction. Thus, the first axis can mainly be used to distinguish between accessions of cultivated and weedy types of var.
frutescens and also between accessions of cultivated var.
frutescens and weedy types of var.
frutescens and var.
crispa. Along axis 1 of the PCA analysis (
Fig. 2), most accessions of cultivated var.
frutescens and weedy types of var.
frutescens and var.
crispa were clearly separated, that is, all ac-cessions of cultivated var.
frutescens formed a group on the positive side, all accessions of weedy var.
frutescens formed a group in the middle between the cultivated var.
frutescens and weedy var.
crispa on the positive and negative sides, and all accessions of weedy var.
crispa formed a group on the negative side on the first axis (
Fig. 2). Therefore, most accessions of cultivated var.
frutescens and related weedy types were clearly separated by the PCA analysis.
DISCUSSION
Morphological difference of crop species within the region of their geographic distribution and cultivation area has received considerable interest in the study of crop species evolution (
Gould and Johnston 1972;
Wyatt and Antonovics 1981;
Ma and Lee 2017;
Ma et al. 2017,
2019;
Sa et al. 2018a). In the process of crop evolution, domestication is an evolutionary process through which domesticated crops become morphologically divergent from their wild species (
Schwanitz 1966;
Hancock 1992;
Harlan 1992;
Purugganan 2019). For example, seed dormancy is one of the largest differences between cultivated crops and the corresponding wild species. In general, in wild species seed dormancy is usually an evolutionary adaptation that prevents seeds from ger-minating under inappropriate ecological conditions that reduce the chances of seedling survival. An important function of seed dormancy is the inhibition or delay of germination. This prevents simultaneous germination of all seeds (
Foley and Fennimore 1998;
Sohindji et al. 2020). However, in most cultivated crops, seed dormancy is not necessary because germination of all seeds should occur simultaneously under the management of the farmers after sowing in the field. Therefore, seed dormancy is a strong choice during domestication by farmers; hence, it is not expected to be present in cultivated crops (
Hancock 1992). In most crops, seed dormancy is one important trait, among others such as seed size, seed shattering, and plant size, for distinguishing cultivated crops and their wild species.
In a previous study by
Sa et al. (2018b), the germination rate of 59
Perilla accessions collected from South Korea was investigated (
Table 1,
Fig. 1). According to their results, the accessions of cultivated P.
frutescens var.
frutescens showed two types in accordance with their germination rates. One type (type I, 15 accessions) showed a high germination rate of 72 to 86% and the other type (type II, 4 accessions) showed a low germination rate of 17 to 19% (
Table 1). These results indicate that, in the case of
Perilla crop, the dormancy of seeds varied in accordance with the accessions of cultivated var.
frutescens and related weedy types. The cultivation methods of the two cultivated types of
Perilla crop are different between South Korea and Japan. For example, in South Korea, cultivated var.
frutescens is cultivated by sowing seeds in spring in a field or seedling box, whereas in Japan seedlings of cultivated var.
crispa that germinate in the spring from seeds that were naturally shattered in the previous fall are transplanted into fields (
Nitta et al. 2003;
Sa et al. 2012). Because of this difference in the traditional cultivation methods of the two cultivated types of
Perilla crop in South Korea and Japan, seed traits, such as seed size, seed hardness, and seed germination rate, of the two cultivated types of
Perilla crop are now considered different. However, in our study, we identified some exceptional accessions of cultivated and weedy types of var.
frutescens with regard to their seed characteristics, as reported previously by
Lee and Ohnishi (2001,
2003). Specifically, most accessions of cultivated var.
frutescens showed comparatively high germination rates, but four accessions (16, 17, 18, 19) of cultivated var.
frutescens showed low germination rates (
Table 1). In addition, one accession of cultivated var.
frutescens showed large and hard seeds and a high germination rate, and one accession of weedy var.
frutescens showed small and soft seeds and a low germination rate. In the case of seed color, accessions of cultivated var.
frutescens showed brown, light brown, gray, and white, while accessions of weedy var.
frutescens showed only gray and dark brown and accessions of weedy var.
crispa showed only dark brown and light brown (
Table 3). These results suggest that the accessions of cultivated var.
frutescens have many variations and differences in their seed characteristics because of the cultivation process by farmers.
According to the result of the correlation analysis, among all combinations, in particular the combinations between QN1 and QL1 (‒0.876**), between QN1 and QL2 (0.826**), between QL1 and QL2 (‒0.871**), between QL4 and QL5 (0.728**), and between QL5 and QL8 (0.895**) show a very high correlation coefficient compared with the other traits at
P < 0.01 (
Table 4). In addition, in the case of traits related to seed characteristics, the combinations between QN1 and QL1 (‒0.876**), between QN1 and QL2 (0.826**), and between QL1 and QL2 (‒0.871**) show a significant negative or positive correlation coefficient compared with the other traits at
P < 0.01 (
Table 4). Also, in the case of traits related to leaf characteristics, the combinations between QL4 and QL5 (0.728**) and between QL5 and QL8 (0.895**) show a significant positive correlation coefficient compared with the other traits at
P < 0.01 (
Table 4). Therefore, among the traits investigated for two cultivated types of
Perilla crop and their weedy types, there was a high correlation in the traits related to seeds and leaves. These results indicate that the seed- and leaf-related traits are useful traits for discriminating between the two cultivated types of
Perilla crop and also between and within the two cultivated types of
Perilla crop and their weedy types. In addition, a previous study by
Ha et al. (2021) reported the iden-tification of SSR (simple sequence repeat) markers related to seed characteristics in 38
Perilla accessions, which were selected from 59
Perilla accessions used in this study. According to their results, a phylogenetic tree constructed using the unweighted pair group method with arithmetic mean (UPGMA) revealed that most accessions of cultivated var.
frutescens could be clearly distinguished from accessions of weedy types of var.
frutescens and var.
crispa by using 29 SSR markers (
Ha et al. 2021).
Meanwhile, the results of PCA analysis (
Fig. 2) show a clear distinction between the accessions of cultivated var.
frutescens and related weedy types, as previously reported by
Lee and Ohnishi (2001),
Sa et al. (2012,
2013), and
Ma and Lee (2017). However, it was not possible to dis-criminate clearly between several accessions of weedy types of var.
frutescens and var.
crispa and also between some accessions of cultivated and weedy types of var.
frutescens. That is, in our study, most accessions of weedy var.
frutescens were located in the middle of accessions of cultivated var.
frutescens and weedy var.
crispa on the first axis (
Fig. 2). These results are thought to provide useful information for understanding the cultivation process of
Perilla crop and the differentiation process of
Perilla crop and related weedy types.
In a previous study by
Lee and Ohnishi (2001) and
Sa et al. (2012,
2013), they were reported that the cultivated var.
crispa has not yet differentiated sufficiently from the weedy var.
crispa to be called a domesticated form; thus, the wild form has been used for cultivation in cultivated var.
crispa. This finding was attributed to all the accessions of cultivated and weedy types of var.
crispa having small seeds (below 2 mm) and seed dormancy, whereas the cultivated var.
frutescens had large seeds (above 2 mm) that were free from seed dormancy. In our study, the same results were obtained as those reported by
Lee and Ohnishi (2001) and
Sa et al. (2012). That is, most accessions of cultivated var.
frutescens had large seeds with soft seeds and had no dormancy, but most accessions of two weedy types of var.
frutescens and var.
crispa showed small seeds, strong dormancy, and hard seeds.
However, in our study, in the case of cultivated var.
frutescens, several accessions (16, 17, 18, 19) showed low germination rates (about 17-19%) and seed hardness (4, 16, 17, 18, 19). These results indicate that, in the case of cultivated var.
frutescens, some accessions are not completely differentiated from the weedy type to the cultivated type. Therefore, in the case of the native accessions of
Perilla crop cultivated in South Korea, it is considered that cultivated or weedy accessions are still cultivated (or mixed) together and used by farmers. In a previous report by
Nitta et al. (2003) and
Sa et al. (2012), the cultivation method and usage method of the two cultivated types of
Perilla crop differ between South Korea and Japan. Therefore, it is thought that the seed size, seed hardness, seed color, and seed dormancy of the two cultivated types of
Perilla crop show a large difference between the two types because of the difference in traditional cultivation methods in South Korea and Japan. Another piece of evidence comes from the phylogenetic tree or UPGMA dendrogram based on amplified fragment length polymorphism (AFLP) and SSR markers (
Lee and Ohnishi 2003;
Lee and Ohnishi 2003,
2015;
Ma et al. 2017,
2019;
Ha et al. 2021). In these studies, the accessions of cultivated and weedy types of var.
crispa could not be clearly discriminated from each other. Therefore, with the results of seed characteristics (seed size, seed dormancy) and the AFLP and SSR data, as in the previous reports by
Lee and Ohnishi (2001,
2003),
Sa et al. (2013,
2015,
2018a), and
Ha et al. (2021), we propose that cultivated var.
crispa has not yet differentiated sufficiently and the wild form is used for cultivation, even though wild species of
Perilla crop have not yet been identified in East Asia.
In conclusion, this study demonstrates the efficacy and utility of PCA analysis using morphological traits, including seed-related properties, such as seed size, seed hardness, seed color, and seed dormancy, in the study of morphological variation of Perilla crop and related weedy types. The results of PCA analysis help to clarify the genetic variation and relationship between Perilla crop and related weedy types, and to better understand the differentiation process of the two cultivated types of Perilla crop and their weedy types.
ACKNOWLEDGEMENTS
This study has been worked with the support of a research grant of Kangwon National University in 2020, and the Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education, Science, and Technology (#2016R1D1A1B01006461), and the Cooperative Research Program for Agriculture Science & Technology Develop-ment (project no. PJ014227032020 and PJ0142272020, PJ0151832020), Rural Development Administration, Republic of Korea.
Fig. 1Collection sites of 59 accessions of
Perilla crop and related weedy types in South Korea. See
Table 1 for the accession numbers.
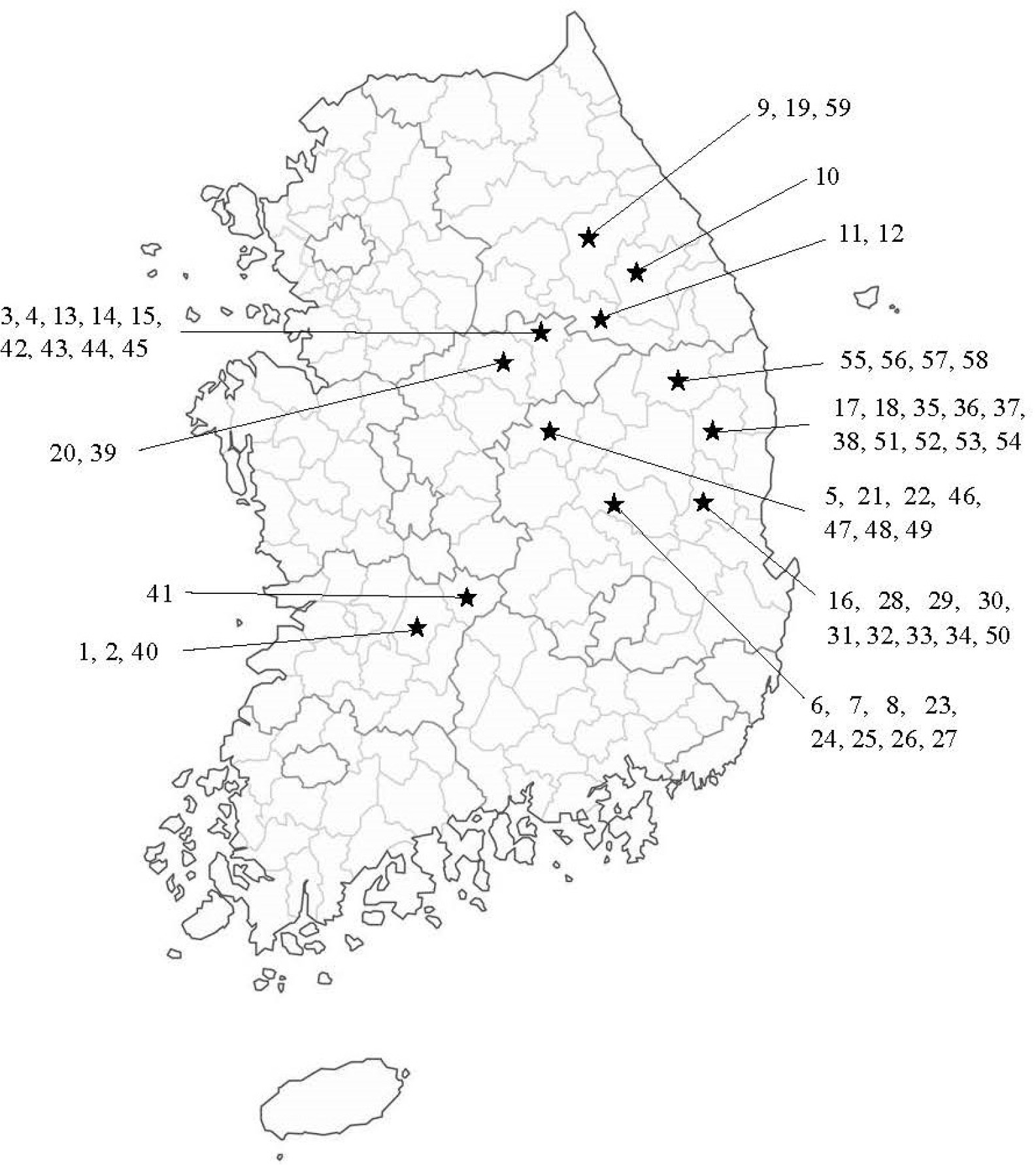
Fig. 2Projection of 59 accessions of Perilla crop and related weedy types in the first and second principal components (● = accessions of cultivated var. frutescens; ○ = accessions of weedy var. frutescens; ■ = accessions of weedy var. crispa).
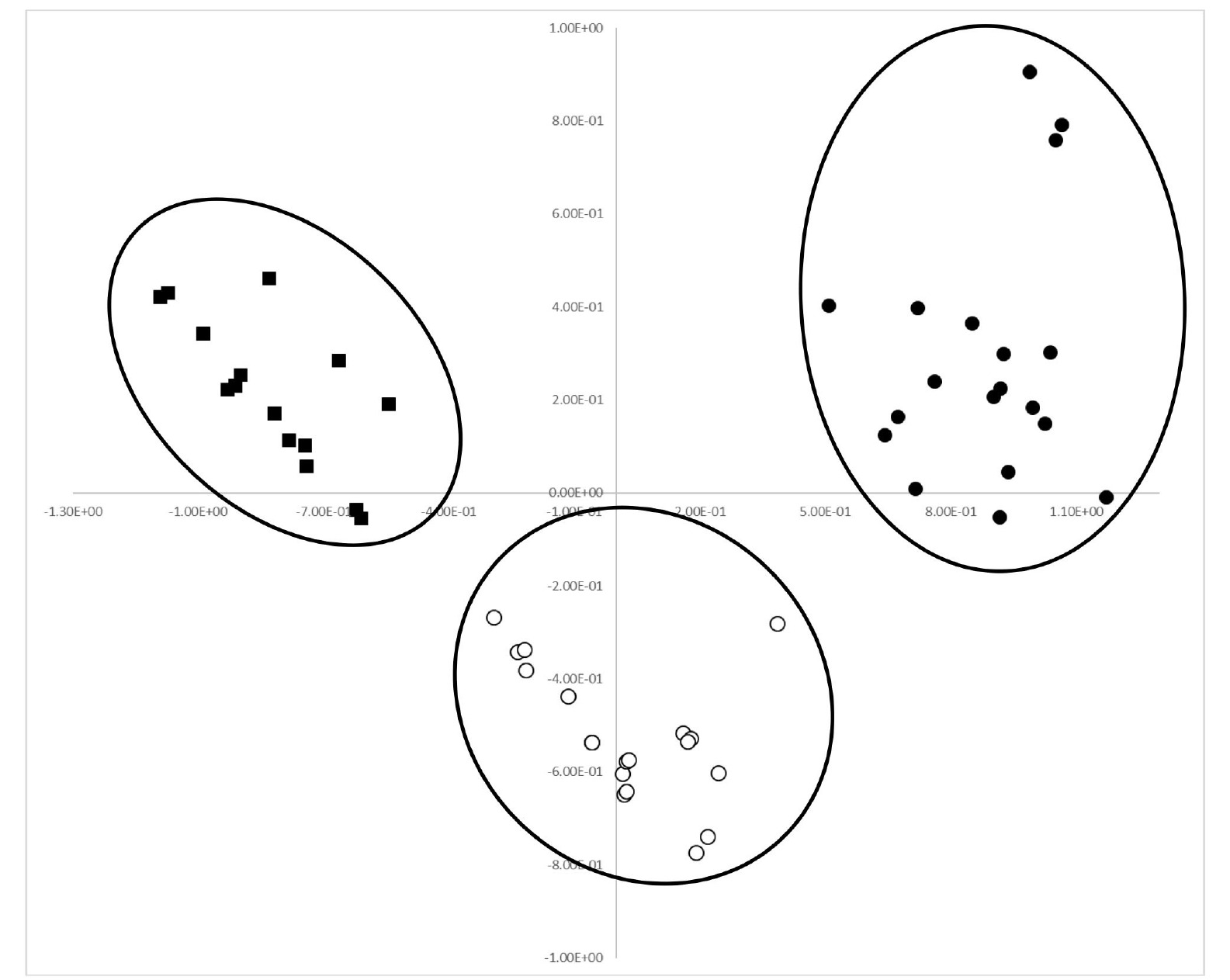
Table 1Accessions of cultivated and weedy types of Perilla crop surveyed in this study.
Table 1
|
Accession no. |
Source of material |
Type |
Germination rate (%)z)
|
|
|
Village, town or city |
Country |
|
1 |
Jinan-gun, Jeollabuk-do |
Korea |
Cultivated var. frutescens
|
72 |
|
2 |
Jinan-gun, Jeollabuk-do |
Korea |
Cultivated var. frutescens
|
79 |
|
3 |
Jecheon-si, Chungcheongbuk-do |
Korea |
Cultivated var. frutescens
|
80 |
|
4 |
Jecheon-si, Chungcheongbuk-do |
Korea |
Cultivated var. frutescens
|
76 |
|
5 |
Mungyeong-si, Gyeongsangbuk-do |
Korea |
Cultivated var. frutescens
|
85 |
|
6 |
Uiseong-gun, Gyeongbuk-do |
Korea |
Cultivated var. frutescens
|
79 |
|
7 |
Uiseong-gun, Gyeongbuk-do |
Korea |
Cultivated var. frutescens
|
80 |
|
8 |
Uiseong-gun, Gyeongbuk-do |
Korea |
Cultivated var. frutescens
|
78 |
|
9 |
Pyeongchang-gun, Gangwon-do |
Korea |
Cultivated var. frutescens
|
74 |
|
10 |
Jeongseon-gun, Gangwon-do |
Korea |
Cultivated var. frutescens
|
85 |
|
11 |
Yeongwol-gun, Gangwon-do |
Korea |
Cultivated var. frutescens
|
82 |
|
12 |
Yeongwol-gun, Gangwon-do |
Korea |
Cultivated var. frutescens
|
86 |
|
13 |
Jecheon, Chungcheongbuk-do |
Korea |
Cultivated var. frutescens
|
80 |
|
14 |
Jecheon, Chungcheongbuk-do |
Korea |
Cultivated var. frutescens
|
83 |
|
15 |
Jecheon, Chungcheongbuk-do |
Korea |
Cultivated var. frutescens
|
86 |
|
16 |
Cheongsong-gun, Gyeongsangbuk-do |
Korea |
Cultivated var. frutescens
|
19 |
|
17 |
Yeongyang-gun, Gyeongsangbuk-do |
Korea |
Cultivated var. frutescens
|
17 |
|
18 |
Yeongyang-gun, Gyeongsangbuk-do |
Korea |
Cultivated var. frutescens
|
18 |
|
19 |
Pyeongchang-gun, Gangwon-do |
Korea |
Cultivated var. frutescens
|
17 |
|
20 |
Chungju-si, Chungcheongbuk-do |
Korea |
Weedy var. frutescens
|
0 |
|
21 |
Mungyeong-si, Gyeongsangbuk-do |
Korea |
Weedy var. frutescens
|
0 |
|
22 |
Mungyeong-si, Gyeongsangbuk-do |
Korea |
Weedy var. frutescens
|
0 |
|
23 |
Uiseong-gun, Gyeongbuk-do |
Korea |
Weedy var. frutescens
|
1 |
|
24 |
Uiseong-gun, Gyeongbuk-do |
Korea |
Weedy var. frutescens
|
0 |
|
25 |
Uiseong-gun, Gyeongbuk-do |
Korea |
Weedy var. frutescens
|
0 |
|
26 |
Uiseong-gun, Gyeongbuk-do |
Korea |
Weedy var. frutescens
|
0 |
|
27 |
Uiseong-gun, Gyeongbuk-do |
Korea |
Weedy var. frutescens
|
0 |
|
28 |
Cheongsong-gun, Gyeongsangbuk-do |
Korea |
Weedy var. frutescens
|
0 |
|
29 |
Cheongsong-gun, Gyeongsangbuk-do |
Korea |
Weedy var. frutescens
|
2 |
|
30 |
Cheongsong-gun, Gyeongsangbuk-do |
Korea |
Weedy var. frutescens
|
0 |
|
31 |
Cheongsong-gun, Gyeongsangbuk-do |
Korea |
Weedy var. frutescens
|
0 |
|
32 |
Cheongsong-gun, Gyeongsangbuk-do |
Korea |
Weedy var. frutescens
|
0 |
|
33 |
Cheongsong-gun, Gyeongsangbuk-do |
Korea |
Weedy var. frutescens
|
0 |
|
34 |
Cheongsong-gun, Gyeongsangbuk-do |
Korea |
Weedy var. frutescens
|
0 |
|
35 |
Yeongyang-gun, Gyeongsangbuk-do |
Korea |
Weedy var. frutescens
|
0 |
|
36 |
Yeongyang-gun, Gyeongsangbuk-do |
Korea |
Weedy var. frutescens
|
0 |
|
37 |
Yeongyang-gun, Gyeongsangbuk-do |
Korea |
Weedy var. frutescens
|
0 |
|
38 |
Yeongyang-gun, Gyeongsangbuk-do |
Korea |
Weedy var. frutescens
|
0 |
|
39 |
Chungju-si, Chungcheongbuk-do |
Korea |
Weedy var. frutescens
|
2 |
|
40 |
Jinan-gun, Jeollabuk-do |
Korea |
Weedy var. crispa
|
0 |
|
41 |
Muju-gun, Jeollabuk-do |
Korea |
Weedy var. crispa
|
0 |
|
42 |
Jecheon-si, Chungcheongbuk-do |
Korea |
Weedy var. crispa
|
0 |
|
43 |
Jecheon-si, Chungcheongbuk-do |
Korea |
Weedy var. crispa
|
0 |
|
44 |
Jecheon-si, Chungcheongbuk-do |
Korea |
Weedy var. crispa
|
1 |
|
45 |
Jecheon-si, Chungcheongbuk-do |
Korea |
Weedy var. crispa
|
0 |
|
46 |
Mungyeong-si, Gyeongsangbuk-do |
Korea |
Weedy var. crispa
|
0 |
|
47 |
Mungyeong-si, Gyeongsangbuk-do |
Korea |
Weedy var. crispa
|
0 |
|
48 |
Mungyeong-si, Gyeongsangbuk-do |
Korea |
Weedy var. crispa
|
0 |
|
49 |
Mungyeong-si, Gyeongsangbuk-do |
Korea |
Weedy var. crispa
|
0 |
|
50 |
Cheongsong-gun, Gyeongsangbuk-do |
Korea |
Weedy var. crispa
|
0 |
|
51 |
Yeongyang-gun, Gyeongsangbuk-do |
Korea |
Weedy var. crispa
|
0 |
|
52 |
Yeongyang-gun, Gyeongsangbuk-do |
Korea |
Weedy var. crispa
|
1 |
|
53 |
Yeongyang-gun, Gyeongsangbuk-do |
Korea |
Weedy var. crispa
|
1 |
|
54 |
Yeongyang-gun, Gyeongsangbuk-do |
Korea |
Weedy var. crispa
|
0 |
|
55 |
Bonghwa-gun, Gyeongsangbuk-do |
Korea |
Weedy var. crispa
|
0 |
|
56 |
Bonghwa-gun, Gyeongsangbuk-do |
Korea |
Weedy var. crispa
|
0 |
|
57 |
Bonghwa-gun, Gyeongsangbuk-do |
Korea |
Weedy var. crispa
|
0 |
|
58 |
Bonghwa-gun, Gyeongsangbuk-do |
Korea |
Weedy var. crispa
|
1 |
|
59 |
Pyeongchang-gun, Gangwon-do |
Korea |
Weedy var. crispa
|
1 |
Table 2Characters used in the morphological analysis of cultivated and weedy types of Perilla crop.
Table 2
|
Abbreviation |
Character |
Unit or Category |
|
QN1 |
Seed germination rate |
0-100 (%) |
|
QL1 |
Seed size |
Large (> 2 mm)-1, small (2 mm <)-2 |
|
QL2 |
Seed hardness |
Hard-1, soft-2 |
|
QL3 |
Seed color |
Dark brown-1, brown-3, light brown-5, gray-7, white-9 |
|
QL4 |
Color of surface leaf |
Light green-1, green-3, deep green-5, green/purple-7, purple-9 |
|
QL5 |
Color of reverse side leaf |
Light green-1, green-3, deep green-5, green/purple-7, purple-9 |
|
QL6 |
Leaf size |
Large-1, middle-2, small-3 |
|
QL7 |
Degree of pubescence |
Slightly pubescent-1, pubescent-3, heavily pubescent-5 |
|
QL8 |
Stem color |
Light green-1, green-3, deep green-5, purple-7 |
|
QL9 |
Flower color |
White-1, white/purple-2, purple-3 |
Table 3Accession number for one quantitative and nine qualitative characters among 59 accessions of cultivated and weedy types of Perilla crop.
Table 3
|
Morphological Character |
Cultivated var. frutescens (n=19) |
Weedy var. frutescens (n=20) |
Weedy var. crispa (n=20) |
|
QN1 (Seed germination rate) |
High (15z)), low (4) |
Low (20) |
Low (20) |
|
QL1 (Seed size) |
Large (19) |
Small (20) |
Small (20) |
|
QL2 (Seed hardness) |
Hard (1), soft (18) |
Hard (19), soft (1) |
Hard (20) |
|
QL3 (Seed color) |
Brown (4), light brown (10), gray (4), white (1) |
Dark brown (16), gray (4) |
Dark brown (19), light brown (1) |
|
QL4 (Color of surface leaf) |
Light green (5), green (12), deep green (2) |
Light green (4), green (12), deep green (4) |
Green (6), deep green (3), green/purple (7), purple (4) |
|
QL5 (Color of reverse side leaf) |
Light green (5), green (14) |
Light green (8), green (9), deep green (3) |
Green/purple (3), purple (17) |
|
QL6 (Leaf size) |
Large (3), middle (14), small (2) |
Middle (15), small (5) |
Middle (17), small (3) |
|
QL7 (Degree of pubescence) |
Slightly pubescent (7), pubescent (3), heavily pubescent (9) |
Slightly pubescent (4), pubescent (9), heavily pubescent (7) |
Slightly pubescent (12), pubescent (8) |
|
QL8 (Stem color) |
Light green (4), green (15) |
Light green (1), green (17), deep green (2) |
Purple (18), green/purple (2) |
|
QL9 (Flower color) |
White (19) |
White (20) |
White/purple (4), purple (16) |
Table 4Pearson correlation coefficient for one quantitative and nine qualitative characteristics in 59 Perilla accessions.
Table 4
|
` |
QN1 |
QL1 |
QL2 |
QL3 |
QL4 |
QL5 |
QL6 |
QL7 |
QL8 |
QL9 |
|
QN1 |
|
−0.876**
|
0.826**
|
0.445**
|
−0.368**
|
−0.428**
|
−0.559**
|
0.466**
|
−0.234 |
−0.550**
|
|
QL1 |
|
|
−0.871**
|
−0.584**
|
0.340**
|
0.441**
|
0.620**
|
−0.565**
|
0.253 |
0.568**
|
|
QL2 |
|
|
|
0.403**
|
−0.410**
|
−0.491**
|
−0.611**
|
0.583**
|
−0.262*
|
−0.531**
|
|
QL3 |
|
|
|
|
−0.075 |
−0.174 |
−0.248 |
0.186 |
−0.093 |
−0.261*
|
|
QL4 |
|
|
|
|
|
0.728**
|
0.323*
|
−0.126 |
0.618**
|
0.517**
|
|
QL5 |
|
|
|
|
|
|
0.409**
|
−0.112 |
0.895**
|
0.692**
|
|
QL6 |
|
|
|
|
|
|
|
−0.364**
|
0.265*
|
0.463**
|
|
QL7 |
|
|
|
|
|
|
|
|
0.046 |
−0.186 |
|
QL8 |
|
|
|
|
|
|
|
|
|
0.638**
|
Table 5Cumulative variances of first and second principal components and the loadings of 10 quantitative and qualitative characters on each principal component.
Table 5
|
Morphological character |
Eigenvectors |
|
|
1 |
2 |
|
QL5 (Color of reverse side leaf) |
−0.849 |
0.367 |
|
QL2 (Seed hardness) |
−0.842 |
0.453 |
|
QL6 (Leaf size) |
−0.838 |
0.436 |
|
QL8 (Stem color) |
−0.836 |
−0.478 |
|
QL1 (Seed size) |
−0.691 |
0.402 |
|
QL3 (Seed color) |
−0.675 |
−0.289 |
|
QL7 (Degree of pubescence) |
0.401 |
0.455 |
|
QL4 (Color of surface leaf) |
0.455 |
−0.388 |
|
QN1 (Seed germination rate) |
0.783 |
0.488 |
|
QL9 (Flower color) |
0.817 |
0.433 |
|
Cumulative variance (%) |
54.1 |
17.9 |
References
- Foley ME, Fennimore SA. 1998. Genetic basis for seed dormancy. Seed Sci. Res.. 8: 173-182.
- Gould SJ, Johnston RF. 1972. Geographic variation. Ann. Rev. Ecol. Syst.. 3: 457-498.
- Ha YJ, Sa KJ, Lee JK. 2021. Identification of SSR markers associated with seed characteristics in Perilla (Perilla frutescens L.). Physiol. Mol. Biol. Plants.. 27: 93-105.
- Hancock JF. 1992. Plant evolution and the origin of crop species. Prentice Hall. Englewood Cliffs, New Jersey, U. S. A.
- Harlan JR. Chapman GP., 1992. Origins and processes of domestication. editor. Grass evolution and domestication. Cambridge University Press. Cambridge, U. K: pp. 159-175.
- Honda G, Koezuka Y, Tabata M. 1990. Genetic studies of fruit color and hardness in Perilla frutescens. Jpn. J. Breed.. 40: 469-474.
- Honda G, Yuba A, Kojima T, Tabata M. 1994. Chemotaxonomic and cytogenetic studies on Perilla frutescens var. citriodora ('Lemon Egoma'). Nat. Med.. 48: 185-190.
- Koezuka Y, Honda G, Sakamoto S, Tabata M. 1985. Genetic control of anthocyanin production in Perilla frutescens. Sho. Zasshi. 39: 228-231.
- Koezuka Y, Honda G, Tabata M. 1986. Genetic control of the chemical composition of volatile oils in Perilla frutescens. Phytochemistry. 25: 859-863.
- Lee JK, Nitta M, Kim NS, Park CH, Yoon KM, Shin YB, et al. 2002. Genetic diversity of Perilla and related weedy types in Korea determined by AFLP analyses. Crop Sci.. 42: 2161-2166.
- Lee JK, Ohnishi O. 2001. Geographical differentiation of morphological characters among Perilla crops and their weedy types in East Asia. Breed. Sci.. 51: 247-255.
- Lee JK, Ohnishi O. 2003. Genetic relationships among cultivated types of Perilla frutescens and their weedy types in East Asia revealed by AFLP markers. Genet. Resour. Crop Evol.. 50: 65-74.
- Li HL. 1969. The vegetables of ancient China. Econ. Bot.. 23: 253-260.
- Lim SE, Sa KJ, Ha YJ, Lee JK. 2019. Genetic analysis of F2 population derived from the cross between Perilla frutescens var. frutescens and var. crispa. Korean J. Breed. Sci.. 51: 184-189.
- Ma SJ, Lee JK. 2017. Morphological variation of two cultivated types of Perilla crop from different areas of China. Hortic. Sci. Technol.. 35: 510-522.
- Ma SJ, Sa KJ, Hong TK, Lee JK. 2017. Genetic diversity and population structure analysis in Perilla frutescens from Northern areas of China based on simple sequence repeats. Genet. Mol. Res.. 16: gmr16039746
- Ma SJ, Sa KJ, Hong TK, Lee JK. 2019. Genetic diversity and population structure analysis in Perilla crop and their weedy types from northern and southern areas of China based on simple sequence repeat (SSRs). Genes Genom.. 41: 267-281.
- Makino T. 1961. Makino's New Illustrated Flora of Japan. Hokuryukan. Tokyo, Japan.
- Nagai I. 1935. On "Shiso" and "Egoma". Agric. Hortic.. 10: 2265-2273. (In Japanese).
- Nitta M. 2001. Origin of Perilla crops and their weedy type. Ph.D Thesis Kyoto University. Kyoto, Japan, 78.
- Nitta M, Lee JK, Kobayashi H, Liu D, Nagamine T. 2005. Diversification of multipurpose plant, Perilla frutescens. Genet. Resour. Crop Evol.. 52: 663-670.
- Nitta M, Lee JK, Ohnishi O. 2003. Asian Perilla crops and their weedy forms: their cultivation, utilization and genetic relationships. Econ. Bot.. 57: 245-253.
- Nitta M, Ohnishi O. 1999. Genetic relationships among two Perilla crops, shiso and egoma, and the weedy type revealed by RAPD markers. Genes Genet. Syst.. 74: 43-48.
- Purugganan MD. 2019. Evolutionary insights into the nature of plant domestication. Curr. Biol.. 29: 705-714.
- Rohlf FJ. 1998. NTSYS-pc: numerical taxonomy and multivariate analysis system. Version: 2.02. Exter Software. Setauket, New York, U. S. A.
- Sa KJ, Choi SH, Ueno M, Lee JK. 2015. Genetic diversity and population structure in cultivated and weedy types of Perilla in East Asia and other countries as revealed by SSR markers. Hortic. Environ. Biotechnol.. 56: 524-534.
- Sa KJ, Choi SH, Ueno M, Park KC, Park YJ, Ma KH, et al. 2013. Identification of genetic variations of cultivated and weedy types of Perilla species in Korea and Japan using morphological and SSR markers. Genes Genom.. 35: 649-659.
- Sa KJ, Choi IY, Park KC, Lee JK. 2018a. Genetic diversity and population structure among accessions of Perilla frutescens (L.) Britton in East Asia using new developed microsatellite markers. Genes Genom.. 40: 1319-1329.
- Sa KJ, Hong TK, Park DH, Lee JK. 2018b. Variation in seed germination according to the storage period after harvest in the accessions of Perilla species. Korean J. Crop Sci.. 63: 48-56.
- Sa KJ, Kim JA, Lee JK. 2012. Comparison of seed characteristics between the cultivated and the weedy types of Perilla species. Hort. Environ. Biotechnol.. 53: 310-315.
- Schwanitz F. 1966. The origin of cultivated plants. Harvard University Press. Cambridge, Massachusetts, U. S. A: pp. 175.
- Sohindji FS, Sogbohossou DEO, Zohoungbogbo HPF, Houdegbe CA, Achigan-Dako EG. 2020. Understanding molecular mechanisms of seed dormancy for improved germination in traditional leafy vegetables: An overview. Agronomy. 10: 57
- Wyatt R, Antonovics J. 1981. Butterfly weed re-revisited: Spatial and temporal patterns of leaf shape variation in Asclepias tuberosa. Evolution. 35: 529-542.
- Yamane Y. 1950. Cytogenetic studies on the genus Perilla and Coleus I. chromosome numbers (in Japanese). Jpn. Genet.. 25: 220.