Abstract
Powdery mildew (PM), caused by the biotrophic fungus Podosphaera xanthii, drastically reduces the yield and quality of melon (Cucumis melo L.). Knowledge of the genetic control and high throughput molecular markers linked with resistance against this disease are essential for breeding programs. The bioassay study of the F1 and F2 populations derived from the parents, ‘PMR 5’ (♂) and ‘SCNU1154’ (♀) revealed a monogenic dominant nature of resistance to the devastating race, race 5. Besides, we developed three SNP based high resolution melting markers, PMm-HRM-1, PMm-HRM-2, and PMm-HRM-3, based on the previously identified SNPs on chromosome 12 and validated them using 8 melon lines and 137 F2 populations. Among these, the SNP of marker PMm-HRM-1 causes a missense mutation in the LRR region of MELO3C002393 and we were able to distinguish the resistant vs susceptible genotypes from eight diverse melon accessions and the segregating F2 population with more than 90% genotyping efficiency. The other two markers were based on intergenic SNPs and had more than 80% genotyping efficiency in F2 population. These markers will be helpful to melon breeders to develop melon cultivars resistant to P. xanthii race 5 via marker assisted breeding programs.
-
Key words: Melon, Powdery mildew, Race 5, SNP, HRM, MAS
INTRODUCTION
Melon (
Cucumis melo L.) is a fruit crop of Cucurbitaceae family that is widely grown across the world in both temperate and tropical regions due to its high flavor, taste, and nutritional value (Fernández‐Trujillo
et al. 2011). Melon fruit contains important nutritional compounds, including sugar, vitamins, minerals, and antioxidants that play significant roles in human health (
Lester 2008). The average production of melon is more than 33 million tons per year (FAOstat 2018). Every year, almost 25% of crops are lost owing to diseases caused by fungal, bacterial, viral, and insect pests (
Agrios 1997).
Powdery mildew (PM) is one of the most devastating and widespread diseases of Cucurbitaceae vegetables, caused by two biotrophic fungi,
Podosphaera xanthii or Golovinomyces cichoracearum, of which the former one has been reported in Korea (
Lee et al. 2014;
Kim et al. 2016a;
Kim et al. 2016b). Among the twenty-eight reported races of
P. xanthii that cause powdery mildew in melon (
McCreight 2006), races 1, 2, and 3 are prevalent in America, and races 0, 4, and 5 were identified in France (
Bardin et al. 1999). Previous studies characterized races 1, N1, N2, and 5 that cause powdery mildew in melon (
Hosoya et al. 1999) and later, seven different races (races 1, 2, 3, 4, 5, 6, and 7) have been reported (
Kuzuya et al. 2004) followed by a new race, N5 in Japan (
Kim et al. 2016b). Two new races, KN1 and KN2, have recently been identified in South Korea as causing severe disease on melon (
Kim et al. 2016a;
Kim et al. 2016b;
Hong et al. 2018). Despite moderate success in controlling
P. xanthii with agrochemicals, cost effectiveness, pathogenic resis-tance to fungicides as a result of long-term use, and environmental hazard are major concerns. This can be addressed by the development of resistance cultivars. However, for race 5, molecular markers and elite cultivars have yet to be reported.
In melon, twelve major quantitative trait loci (QTLs) associated with resistance to different races of powdery mildew have been identified (
Liu et al. 2010). In addition, many powdery mildew studies have been reported in melon (
Li et al. 2017). Genes and QTLs conferring resistance to powdery mildew have been reported on chromosomes 2, 5, and 12 (
Ning et al. 2014). Among them, Pm-R1-2 were against race 1 and Pm-R5 on LG V was against race 5 in the TGR-1551 line (Yuste‐Lisbona
et al. 2010). Using simple sequence repeat (SSR) markers, the QTL PmV.1 against races 1, 2, and 3 and QTL PmXII.1 against races 1, 2, and 5 on LG XII was identified in PI 124112 (
Perchepied et al. 2005).
Resistant (R) genes play important roles in resistance to plant disease (
Ellis et al. 2000;
Harris et al. 2013). R genes have been identified in several plant species, including Arabidopsis thaliana, Oryza sativa, and Cucumis sativus (
Meyers et al. 2003;
Monosi et al. 2004;
Wan et al. 2013). R genes are reliable candidate genes for disease resistance that can be easily converted into molecular markers through the use of SSRs, single nucleotide polymorphisms (SNPs), and insertion/deletions (InDels). Molecular markers have been instrumental in the selection of disease-resistant cultivars and accelerating breeding cycles. Among the many molecular marker methods, HRM analysis is a closed-tube method for measuring the fluorescence reduc-tion of intercalating dye during the dissociation of double- stranded DNA (
Vossen et al. 2009). HRM markers have been developed as a quick, accurate, and high throughput marker for plant trait, disease diagnostics, and SNP genoty-ping (
Chatzidimopoulos et al. 2014; Chatzidimopoulos
et al. 2019) Furthermore, this analysis has been widely used to study genetic diversity in several plant species using mole-cular markers such as SNPs and InDels (
Mackay et al. 2008;
Ganopoulos et al. 2011;
Distefano et al. 2012). There are few high throughput molecular markers for powdery mildew. We previously reported race 5-specific SNPs from whole-genome re-sequencing (WGR) on chromosomes 2, 5, and 12 and developed dCAPs markers based on three SNPs on chromosome 12 (
Natarajan et al. 2016;
Howlader et al. 2020). We converted these SNPs into HRM because HRM marker facilitate mass screening and is high throughput marker. Besides, the inheritance of resistance to
P. xanthii race 5 has not been investigated yet. In this study, we therefore aimed to determine the inheritance pattern of resistance to
P. xanthii race 5 in melon and develop high throughput HRM markers.
MATERIALS AND METHODS
Plant materials and growth conditions
The melon parental lines, ‘PMR 5’ (♂) and ‘SCNU1154’ (♀), resistant and susceptible to
Podosphaera xanthii race 5, respectively, were used to develop F
1 and F
2 generations. Besides, the susceptible lines (susceptible to
P. xanthii race 5) Edisto, WMR29, PMR 45, and NSL 74171; and resistant lines MR1, PI 414723, PI 124112, and NSL 86629 were used to validate the developed markers. All the seeds were obtained from the Department of Horticulture, Sunchon National University, Korea (
Kim et al. 2016a;
Kim et al. 2016b). Plants were grown in a plant culture chamber at 25 ± 2℃ and 60% relative humidity under long-day conditions (14 hours light and 10 hours dark cycle / 80-120 mmol∙m
‒2∙s
‒1 PPFD).
P. xanthii race 5 was obtained from the Department of Horticulture, Sunchon National University, Korea (
Kim et al. 2015) and was cultured on melon leaves in petri dishes on agar medium (Agar powder 6 g.L-1, Saccharose 10 g.L-1, D-Mannitol 20 g.L-1) and cultured fresh leaves were maintained in a plant culture chamber. Melon plants were inoculated with
P. xanthii race 5 when the third true leaf was fully open by hand using a spray bottle with a spore concentration of 5 × 10
5 spore/mL, whereas control plants were sprayed with plain water. To maintain over 95% humidity, inoculated plants were covered with a plastic cover.
Assessment of P. xanthii race 5 resistance and statistical analysis
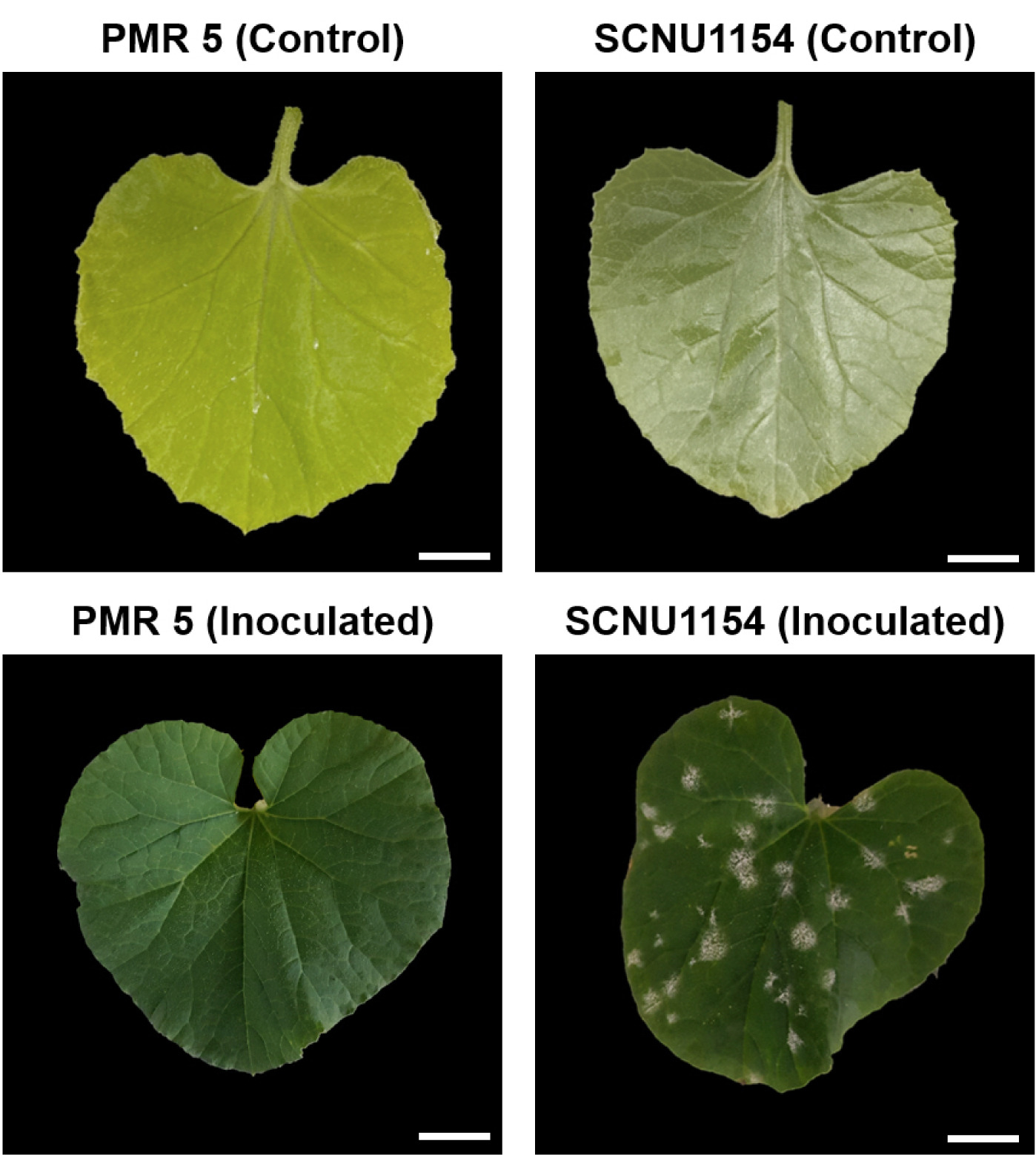
Inoculated leaves evaluated the disease symptoms at two weeks after inoculation (WAI). The plants were considered
P. xanthii race 5-susceptible when the white spores of PM covered more than 10% of the overall melon leaves (
Fig. 1). A chi-square (
x2) test for goodness-of-fit was used in the XLSTAT software (Addinsoft, New York, USA) to dete-rmine deviations of observed data from expected segre-gation ratios. Heat map was constructed through Heatmapper (
http://www.heatmapper.ca/; accessed March 9, 2022).
Genomic DNA (gDNA) was extracted from young leaves of ten melon plants using DNeasy Plant Mini Kit (QIAZEN, Hilden, Germany) according to the manufacturer’s instructions. The concentration and quality of total gDNA were determined using an ND-1000 Micro-spectrophotometer (NanoDrop Technologies Inc., Wilmington, DE, USA). High resolution melting (HRM) analysis combined with 3’-blocked and unlabeled oligonucleotide probe (HybProbe) specific to the SNP site was used to detect SNPs. The Primers used in this study were synthesized by Macrogen, Seoul, Korea (
Table 1). The gDNA was then used for HRM analysis with a LightCycler 96 instrument (Roche, Mannheim, Germany). HRM was performed in 10 mL reaction mixtures containing 1 mL at 5ng/mL, 0.1 mL forward primer, 0.5 mL reverse primer, 0.5 mL probe (10 pmol), 0.3 mL SYTO 9 fluorescent dye (Invitrogen, Thermo Fisher Scientific, USA), 5 mL HS prime LP premix (GENETBIO, Daejeon, Korea), and 2.6 mL DDW. HRM conditions were as follows: an initial preincubation at 95℃ for 300 s; followed by 40 cycle of 3 step amplification (95℃ for 10 seconds, 64℃ to 56℃ for 15 seconds under touch down and 72℃ for 15 seconds); four readings per ℃ at the final step after 60 seconds at 95℃, 60 seconds at 40℃, and 1 second at 97℃. HRM data was performed using LightCycler 96 software at 100% discrimination for delta Tm and curve shape with a 0.2 positive/negative threshold level.
RESULTS
Inheritance of resistance to P. xanthii race 5 in melon
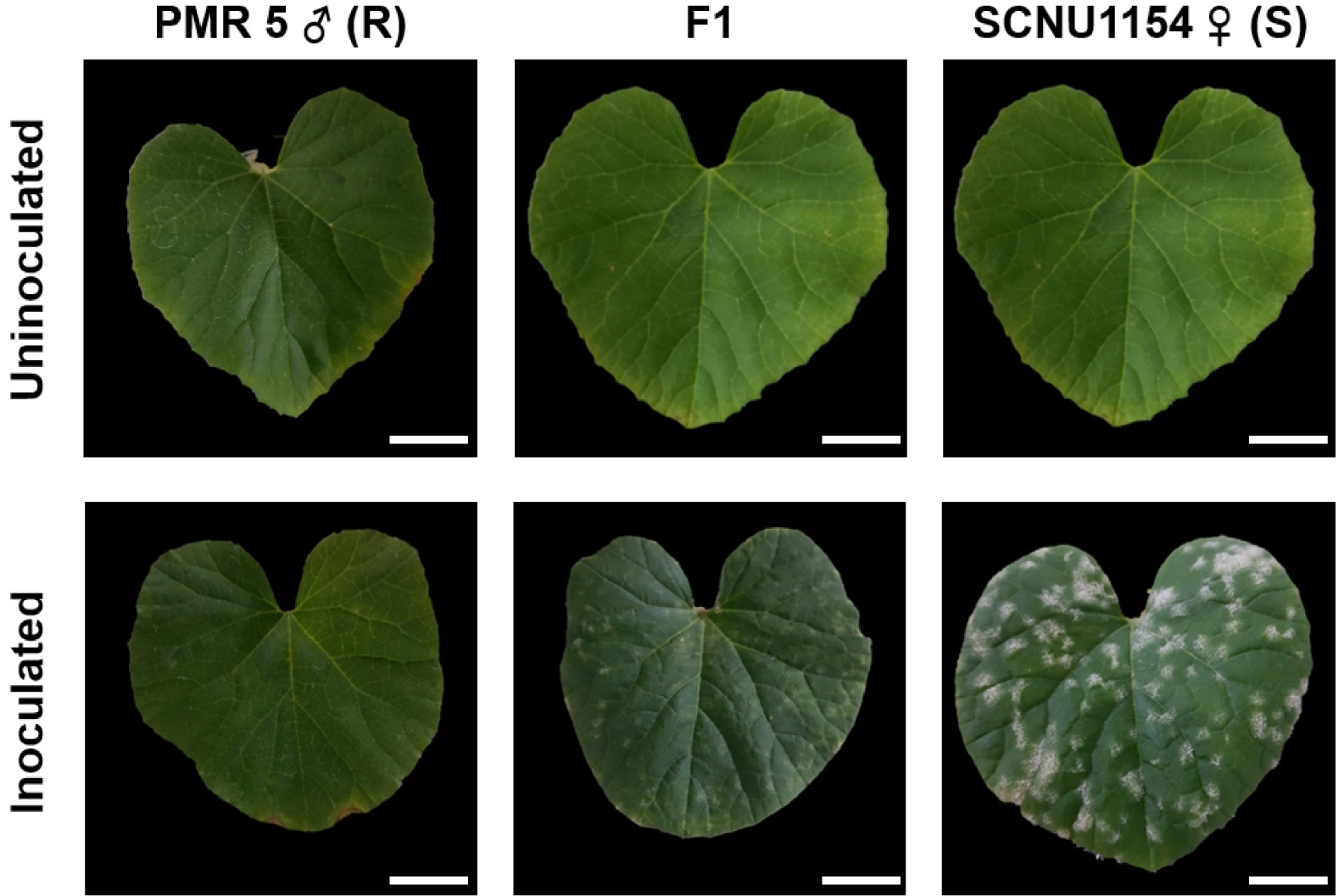
To determine whether the resistance trait to
P. xanthii race 5 is dominant or recessive, we used SCNU1154, PMR 5, and F
1 (PMR 5 × SCNU1154) generations. The suscep-tible parental line, SCNU1154, had more than 50% PM infected leaf area, while less than 10% of the leaf area was affected in resistant parental line, PMR 5 (
Fig. 2) by
P. xanthii race 5. Furthermore, bioassay results revealed that the F
1 (SCNU1154 × PMR 5) populations had less than 10% infected leaf area, indicating that resistance to
P. xanthii race 5 is inherited as a dominant trait. An F
2 population derived from a cross between the race 5-susceptbile melon line, SCNU1154 and -resistant line, PMR 5, to investigate the inheritance pattern of
P. xanthii race 5 resistance in melon. Among the 137
P. xanthii race 5 inoculated F
2 plants, 104 plants showed resistance and 33 plants showed susceptible reactions (
Table 2). A chi-square (
x2) test revealed that resistance to
P. xanthii race 5 seg-regates at a 3:1 (resistant: susceptible) ratio. These results suggest that a single dominant gene is involved in
P. xanthii race 5 resistance.
The identification of genes related to
P. xanthii race 5-resistant was necessary for the development of
P. xanthii race 5-resistant melon cultivars. Our research group recently reported potential candidates for three SNPs in
P. xanthii race 5 resistance genes on chromosome 12 (
Table 3) (
Natarajan et al. 2016;
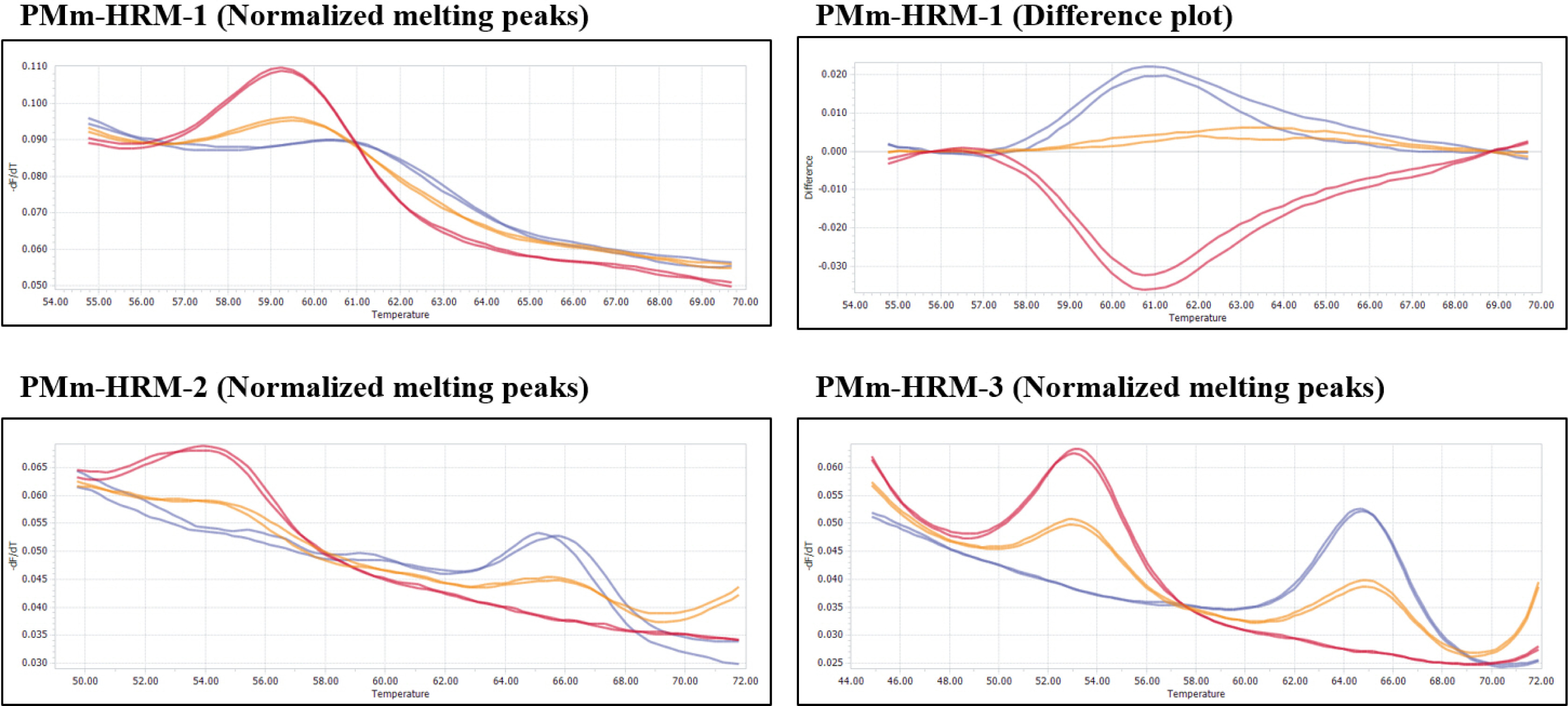
Howlader et al. 2020). However, these SNPs were validated only in melon PI accessions. Therefore, we developed three HRM markers, namely PMm-HRM-1, PMm-HRM-2, and PMm-HRM-3, using the PMR 5, SCNU1154, and F1 generations to validate these three SNPs (
Table 1 and
Fig. 3). These genotypic polymorphisms of HRM markers were consistent with
P. xanthii race 5 resistance phenotypes. Therefore, HRM mar-kers can be used to identify
P. xanthii race 5 resistance.
To validate the association of three HRM markers, PMm-HRM-1, PMm-HRM-2, and PMm-HRM-3 with resistance to
P. xanthii race 5, we used eight diverse melon accessions (
Supplementary Fig. 3 and
Supplementary Table 1) and 137 F
2 populations (
Supplementary Table 2). Among the eight diverse melon accessions, the HRM markers, PMm-HRM-2 and PMm-HRM-3 success-fully distinguished resistance vs susceptible genotypes of all, while the geno-typing of the marker, PMm-HRM-1 was miss-matched for only two genotypes (PI 124112 and NSL 86629) (
Supplementary Fig. 2 and
Supplementary Table 1). However, the genotypes Edisto, WMR29, PMR45, and NSL 74171 showed diffe-rent melting curve peaks for the marker PMm-HRM-3 (
Supplementary Fig. 3), which could be due to genetic differences of these two genotypes with other genotypes. HRM assay of the 137 F
2 individuals also indicated high detection (of resistance status) accuracy of 90.07%, 87.23%, and 82.27% for the developed markers, PMm-HRM-1, PMm-HRM-2, and PMm-HRM-3, respectively. High cor-relations between the phenotypic and genotypic assays with the diverse melon genotypes and 137 segregating F
2 individuals indicate the close linkage of these three SNPs, SNPR5-119, SNPR5-120, and SNPR5-121, with
P. xanthii race 5 resistance in melon (
Supplementary Table 2).
DISCUSSION
The use of molecular markers to select disease resistance genotypes has more advantages than traditional phenotypic screening. Target genes and disease-resistant chromosomal regions, or QTLs, can be identified using molecular markers linked to specific phenotypic traits (
Tanksley et al. 1989). SNPs are biallelic in nature and stable in most genomes because they are less susceptible to mutations than SSRs (
Kota et al. 2003). Furthermore, polymorphism of SSR markers in biological studies, particularly in fungal systems, is a critical constraint (
Mahfooz et al. 2016). SNPs can either regulate a phenotype directly or be linked to a phenotype as a result of linkage disequilibrium. As a result, SNPs could be used to create a more accurate high-density genetic map (
Li et al. 2015;
Kessuwan et al. 2016).
The yield and quality of melon are significantly reduced by many races of the powdery mildew (PM) fungus
P. xanthii; on the other hand, some cultivars show resistance to specific
P. xanthii races (
McCreight 2006;
Kim et al. 2016a;
Kim et al. 2016b;
Lebeda et al. 2016). To under-stand the PM resistance response, an interaction between the plant and the pathogenic
P. xanthii race is necessary. The development of melon cultivars with a gene for resistance specific to each
P. xanthii race may aid in the explanation of the pathogenesis of PM races and the interactions between genes and races (
Fukino et al. 2008). Therefore, the development of
P. xanthii race-specific SNP markers that are strongly associated with this desirable trait could be a useful tool for marker-assisted selection (MAS) in melon breeding programs.
We investigated the inheritance pattern of resistance to PM against a widespread
P. xanthii race 5 and developed high throughput molecular markers for selecting resistant lines. In our bioassay after inoculation with
P. xanthii race 5, the F
1 populations from a cross between
P. xanthii race 5-susceptbile (SCNU1154) and -resistant (PMR 5) lines were resistant, indicating the dominant nature of resistance to PM (
Fig. 3 and
Table 2). A phenotypic ratio of 3:1 (resistant: susceptible) was found in 137 F
2 plants, indicating a monogenic trait of resistance to
P. xanthii race 5 (
Supplementary Table 2). Various patterns of inheritance of resistance to powdery mildew in melon have been previously identified, including polygenic against race N1 (
Kim et al. 2016b), co-dominant against races 1, 2 and 5 (
Yuste-Lisbona et al. 2011), and dominant against races 1 and pxCh1 (
Fukino et al. 2008;
Liu et al. 2010). Develop-ment of markers, especially the high throughput ones, is essential for mass screening and speed breeding these days (
Thomson 2014;
Watson et al. 2018;
Wanga et al. 2021). Several SSR markers: CMBR150 and CMBR111 (
Fukino et al. 2008), CMN1-38 (
Fukino et al. 2007), CMBR120 (
Kim et al. 2016b), and TJ29 (
Gonzalo et al. 2005) CMCT170b and CMMS30-3 (
ÜNLÜ et al. 2022); SRAP marker: Pm-8 (
Liu et al. 2010) and CAPs markers: CAPS- Dde I (
Zhang et al. 2013), and PM2-CAPS and PM5-CAPS (
Yuste-Lisbona et al. 2011) were linked with linked to PM resistance against specific races (
Cui et al. 2022). These markers were mainly used to develop linkage maps and are of low throughput. These days, SNPs can easily be iden-tified and SNPs linked with specific phenotypic expres-sions can be used for developing high throughput geno-typing upon validation in diverse natural populations and in developed segregating populations (
Mammadov et al. 2012;
He et al. 2014;
Thomson 2014;
Yang et al. 2015).
Based on the whole genome re-sequencing of PM susceptible genotype, SCNU1154 and resistant genotypes, MR-1, and PMR 5, we have identified the genome-wide SNPs, of which 112 SNPs and 12 InDels were observed in powdery mildew responsive chromosomes 2, 5, and 12 (
Natarajan et al. 2016;
Howlader et al. 2020). Among these, three polymorphic SNPs, namely, SNPR5-119, SNPR5-120, and SNPR5-121 demonstrated race 5‑specific genetic varia-tion in melon (
Kim et al. 2016a;
Kim et al. 2016b). Among these, three SNPs, namely, SNPR5_119, SNPR5_120, and SNPR5_121 on chromosome 12 (closely located to the SSR marker CMBR150) were later identified to be asso-ciated with the phenotypic variation against race 5-specific susceptible (SCNU1154, PMR45, WMR29, and Edisto47) and resistant (PI 414723, PMR 5, and MR1) lines in
C. melo via derived cleaved amplified polymorphic sequence (dCAPS) assay, indicating their linkage with resistance against PM race-5 (
Howlader et al. 2020). Here, we designed HRM markers and validated their genotyping efficacy in diverse natural populations and the developed F2 segregating generations. Among these three markers, PMm-HRM-1 showed more than 90% accuracy in genoty-ping the resistance status of both melon accessions and F
2 segregating populations, as opposed to only ∼80% accuracy of the other two markers, PMm-HRM-2 and PMm-HRM-3, in F2 population (
Supplementary Table 1). The SNPs,
SNPR5-120 and
SNPR5-121, are located in the intergenic region between the genes,
MELO3C002316 and
MELO 3C002317. Whereas,
SNPR5-119 is located in the intra-genic region of the intron-less leucine-rich repeat (LRR) receptor-like protein kinase family gene,
MELO3C002393 on chromosome 12 (
Table 3). Plant
R genes are mainly encoded with nucleotide-binding site leucine-rich repeat (NBS-LRRs), LRR, receptor-like protein kinases (RLKs) and LRR-RLKs domains, all of which are involved in plant defense against pathogens (
Ellis et al. 2000;
Eitas and Dangl 2010;
Brotman et al. 2013;
Harris et al. 2013;
Kourelis and Van Der Hoorn 2018). The LRR domain provides pathogen recognition specificity (DeYoung and Innes 2006;
Eitas and Dangl 2010). Moreover, the NBS-LRR domain in plant immune receptors relies on the LRR domain to perform many important roles (
Padmanabhan et al. 2009), and mutations in the LRR domain may result in a susceptible phenotype.
SNPR5-119 results in a missense mutation (Aspartic acid [N] → Asparagine [D]) in the LRR region of
MELO3C002393, indicating its potential role in eventual protein product and thereby, influencing the ability to confer resistance against race 5 (
Supplementary Fig. 4). Taken together, these findings suggest that the SNP maker, PMm-HRM-1, can effectively be used in HRM assay to detect
P. xanthii race 5 resistant and susceptible melon genotypes and, hence, can be used as high through-put molecular markers in MAS based breeding programs.
CONCLUSIONS
In this study, we showed that P. xanthii race 5 resistance in melon line ‘PMR 5’ is regulated by a monogenic dominant gene. To validate the three SNPs, SNPR5-119, SNPR5-120, and SNPR5-121 on chromosome 12, we developed three HRM markers, ‘PMm-HRM-1’, ‘PMm- HRM-2’, and ‘PMm-HRM-3’, respectively. These three markers showed consistency with the phenotypic results of both diverse melon accessions and F2 populations. Among these, “PMm-HRM-1” marker showed high accuracy com-pared to other markers. We suggested three candidate genes associated with P. xanthii race 5 resistance as key genes for further candidate gene analysis. These findings will be helpful in the incorporation of P. xanthii race 5 resistance in commercial melon cultivars.
DECLARATIONS
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Availability of data and materials
The raw sequence data from this study have been de-po-sited in the publicly accessible National Center for Biotech-nology Information (NCBI,
https://www.ncbi.nlm.nih.gov/) database as PRJNA804585. The datasets supporting the conclusions of this article are included within the article and its additional files.
STATEMENT
All our experiments complied with local and national regulations.
Supplemental Materials
ACKNOWLEDGEMENTS
Not applicable.
-
CONFLICT OF INTEREST
The authors declare that they have no competing interests.
-
FUNDING
This work was supported by Korea Institute of Planning and Evaluation for Technology in Food, Agriculture and Forestry (IPET) through Digital Breeding Transformation Technology Development Program, funded by Ministry of Agriculture, Food and Rural Affairs (MAFRA) (Project number 322071-03) and Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education (2020R1A6A3A 13077074).
-
AUTHOR’S CONTRIBUTIONS
I.-S.N. and H.-J.J. conceptualized the work, J.-E.H. conducted all experiments and J.-E.H. & M.R.H. prepared the data, interpreted the results, and wrote the manuscript. All authors read and approved the final manuscript.
Fig. 1Phenotypes of melon lines PMR 5 (resistant) and SCNU1154 (susceptible) after inoculation with P. xanthii race 5. Scale bar = 2 cm.
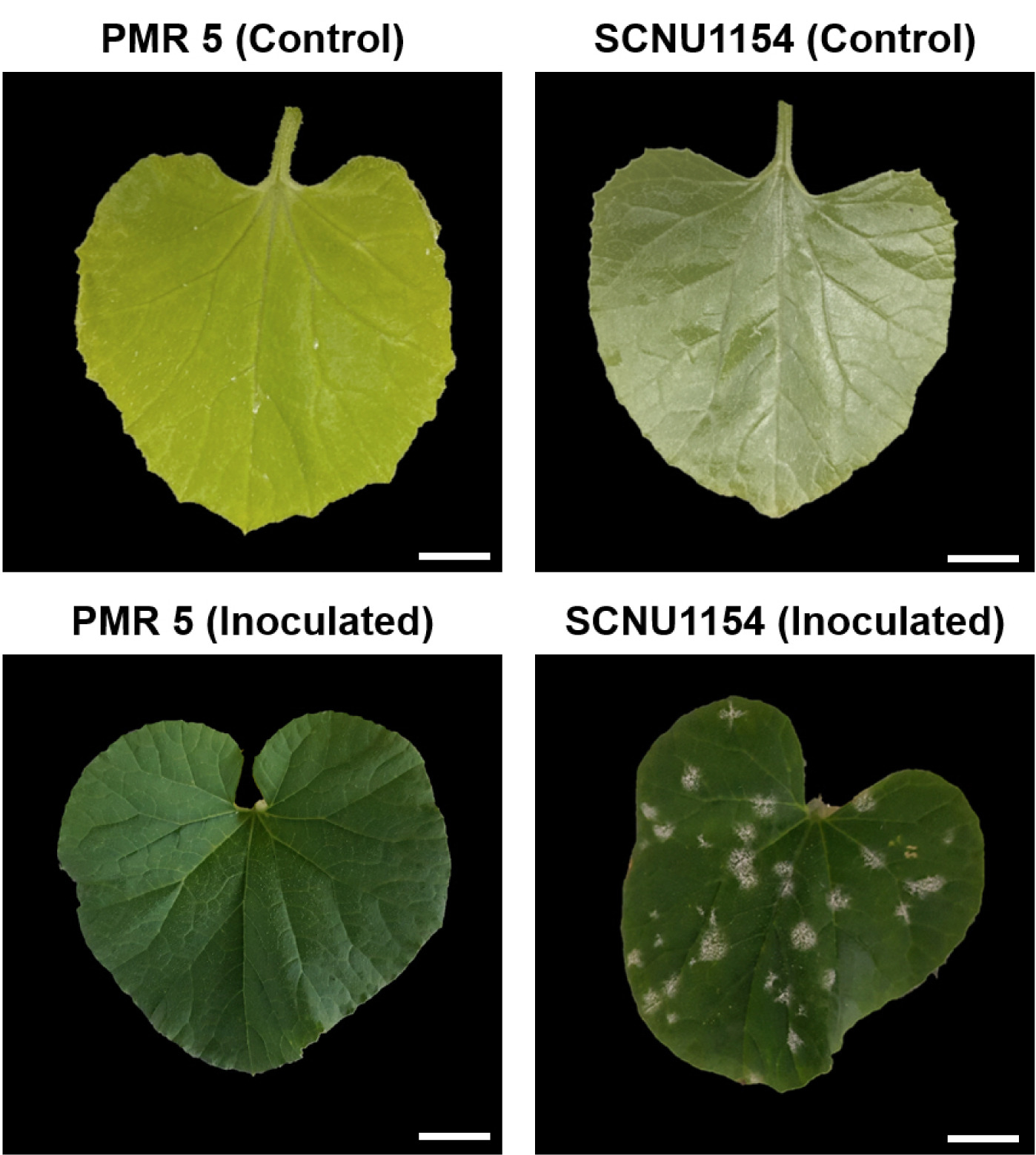
Fig. 2Phenotypes of the resistant and susceptible, and their F1 generation two-weeks after inoculation with P. xanthii race 5. R: Resistance, S: Susceptible. Scale bar = 2 cm.
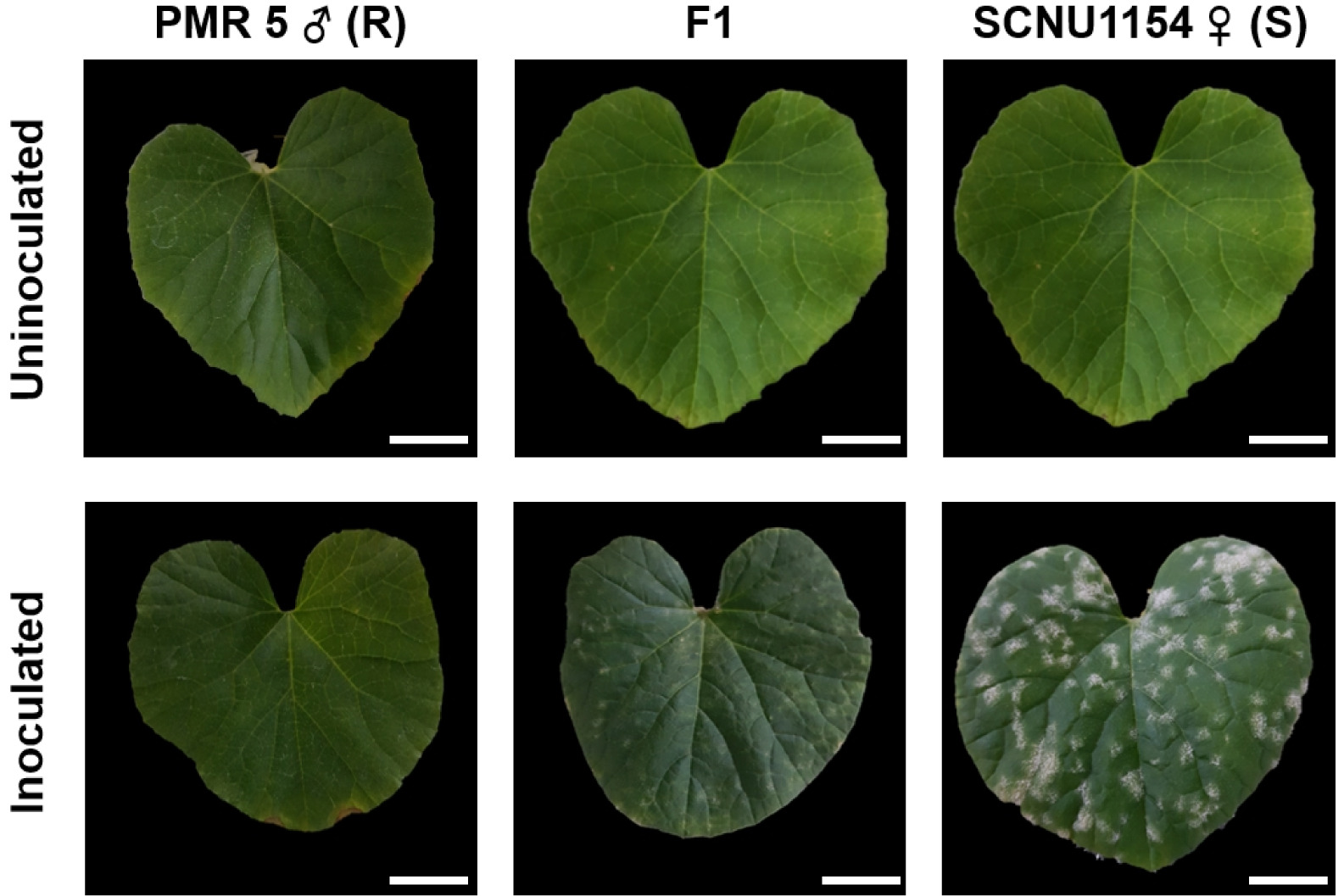
Fig. 3Melting curves of three HRM markers, PMm-HRM-1, PMm-HRM-2 and PMm-HRM-3, developed and validated in this study. Blue curves: Plants of ‘PMR 5’ genotype (resistant), red curves: Plants of ‘SCNU1154’ genotype (susceptible), Orange curves: heterozygous plants.
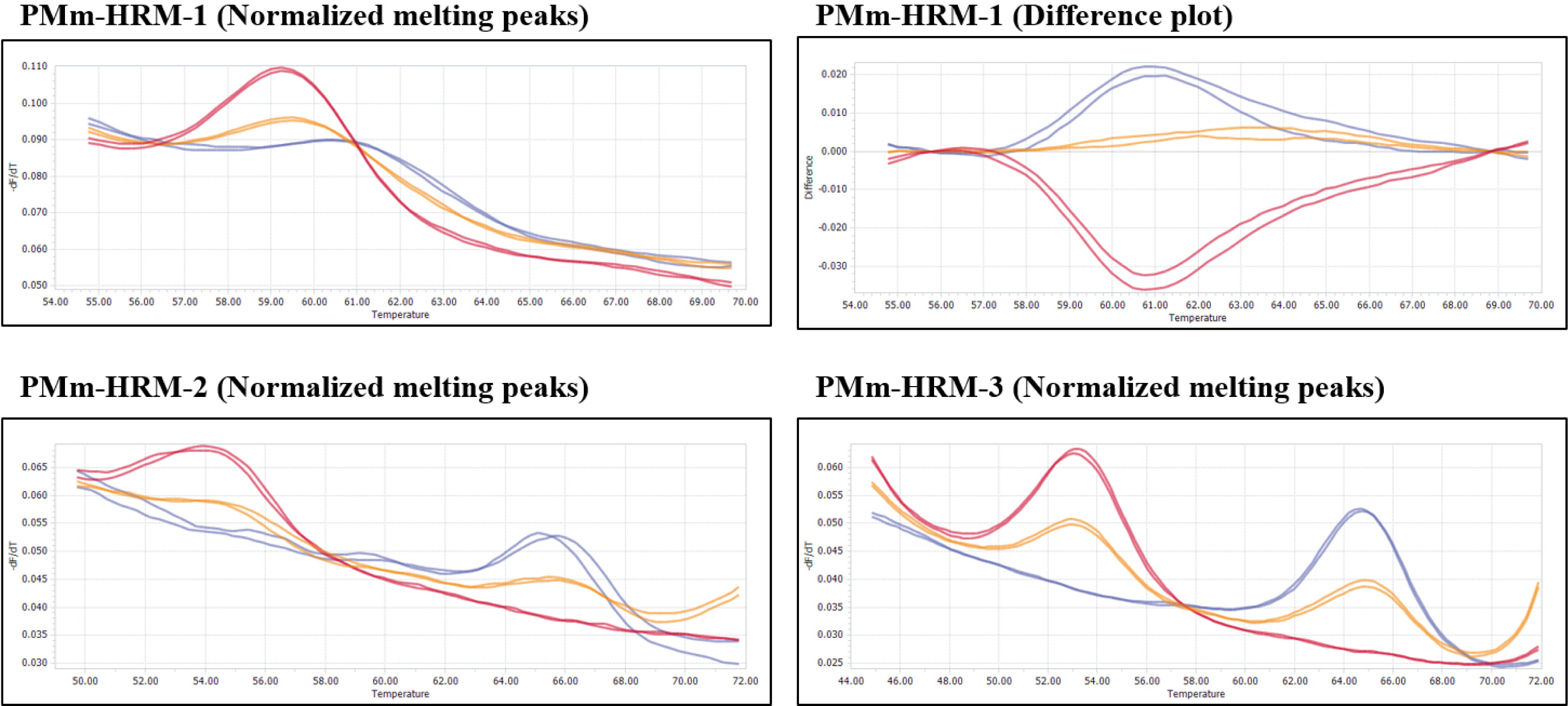
Table 1Primer and probe sequences used for discriminating the resistance vs susceptible genotypes via High Resolution Melting (HRM) assay.
Table 1
|
Marker name |
SNP name |
Primer (5’-3’) |
SNPs |
|
Direction |
Sequence |
|
PMm-HRM-1 |
SNPR5-119 |
Forward |
CAAATAAAGAACCTTCAGTTTGAATGG |
|
|
|
Reverse |
TGAGTTTAACAATTACCAAAATTCATGG |
|
|
|
Probe |
GTTTATCTTTTGCCTCGATGCGAAA |
T > C |
|
PMm-HRM-2 |
SNPR5-120 |
Forward |
TATTAGATAAGGAATCAGCCCTTCG |
|
|
|
Reverse |
GTTAGCCAAATGCAGATCAAATCAG |
|
|
|
Probe |
TGGATCTGAATGGTTTCCTGAAAC |
G > A |
|
PMm-HRM-3 |
SNPR5-121 |
Forward |
ATATTAGATAAGGAATCAGCCCTTCG |
|
|
|
Reverse |
CATATCATTTTACATATACCAACCTAGC |
|
|
|
Probe |
TTTCCTGAAACTGGGATTCCGTTTA |
T > A |
Table 2Inheritance of P. xanthii race 5 resistance in Cucumis melo.
Table 2
|
Crosses |
Generation |
Number of susceptible plants |
Number of resistant plants |
Expected
ratio (S:R) |
Chi-Square
(x2) |
P
|
|
SCUN1154 ♀ (S) |
P1
|
12 |
0 |
- |
- |
- |
|
PMR 5 ♂ (R) |
P2
|
0 |
12 |
- |
- |
- |
|
PMR 5 × SCUN1154 |
F1
|
0 |
12 |
- |
- |
- |
|
PMR 5 × SCUN1154 |
F2
|
33 |
104 |
1:3 |
0.06 |
0.805 |
Table 3Genomic location of SNPs and their variation with the melon genome.
Table 3
|
Chromosome |
SNP name |
SNP position |
Gene |
Ref*
|
SCNU1154 |
PMR 5 |
|
Chr12 |
SNPR5-119 |
23,192,595 |
Intragenic region (Exon) |
MELO3C002393 |
T |
C/C |
T/T |
|
Chr12 |
SNPR5-120 |
23,769,828 |
Intergenic region |
MELO3C002316-MELO3C002317 |
G |
A/A |
G/G |
|
Chr12 |
SNPR5-121 |
23,769,833 |
Intergenic region |
MELO3C002316-MELO3C002317 |
T |
A/A |
T/T |
References
- Agrios G. 1997. How plants defend themselves against pathogens. Plant Pathol.. 93: 114.
- Bardin M, Carlier J, Nicot P. 1999. Genetic differentiation in the French population of Erysiphe cichoracearum, a causal agent of powdery mildew of cucurbits. Plant Pathol.. 48: 531-540.
- Brotman Y, Normantovich M, Goldenberg Z, Zvirin Z, Kovalski I, Stovbun N, et al. 2013. Dual resistance of melon to Fusarium oxysporum races 0 and 2 and to Papaya ring-spot virus is controlled by a pair of head-to- head-oriented NB-LRR genes of unusual architecture. Mol Plant.. 6: 235-238.
- Chatzidimopoulos M, Ganopoulos I, Madesis P, Vellios E, Tsaftaris A. Pappas A C2014. High-resolution melting (HRM) analysis for rapid detection and characterization of Botrytis cinerea phenotypes resistant to fenhexamid and boscalid. Plant Pathol.. 63: 1336-1343.
- Chatzidimopoulos M, Ganopoulos I, Moraitou-Daponta E, Lioliopoulou F, Ntantali O, Panagiotaki P, et al. 2019. High-resolution melting (HRM) analysis reveals genotypic differentiation of Venturia inaequalis populations in Greece. Front. Ecol. Evol.. 7: 489
- Cui L, Siskos L, Wang C, Schouten HJ, Visser RG, Bai Y. 2022. Breeding melon (Cucumis melo) with resistance to powdery mildew and downy mildew. Hortic. Plant J.. 5: 545-561.
- Distefano G, Caruso M, La Malfa S, Gentile A, Wu SB. 2012. High resolution melting analysis is a more sensitive and effective alternative to gel-based platforms in analysis of SSR‒An example in Citrus. PLoS ONE. 7: e44202
- Eitas TK, Dangl JL. 2010. NB-LRR proteins: pairs, pieces, perception, partners, and pathways. Curr. Opin. Plant Biol.. 13: 472-477.
- Ellis J, Dodds P, Pryor T. 2000. Structure, function and evolution of plant disease resistance genes. Curr. Opin. Plant Biol.. 3: 278-284.
- Monforte AJ. Fernández‐Trujillo JP.Picó B.Garcia‐Mas J.Álvarez JM.2011. Breeding for fruit quality in melon. Breeding for fruit quality: 261-278..
- Fukino N, Ohara T, Monforte AJ, Sugiyama M, Sakata Y, Kunihisa M. 2008. Identification of QTLs for resistance to powdery mildew and SSR markers diagnostic for powdery mildew resistance genes in melon (Cucumis melo L.). Theor. Appl. Genet.. 118: 165-175.
- Fukino N, Sakata Y, Kunihisa M, Matsumoto S. 2007. Characterisation of novel simple sequence repeat (SSR) markers for melon (Cucumis melo L.) and their use for genotype identification. J. Hortic. Sci. Biotechnol.. 82: 330-334.
- Ganopoulos I, Argiriou A, Tsaftaris A. 2011. Microsatellite high resolution melting (SSR-HRM) analysis for authenticity testing of protected designation of origin (PDO) sweet cherry products. Food Control. 22: 532-541.
- Gonzalo M, Oliver M, Garcia-Mas J, Monfort A, Dolcet- Sanjuan R, Katzir N, et al. 2005. Simple-sequence repeat markers used in merging linkage maps of melon (Cucumis melo L.). Theor. Appl. Genet.. 110: 802-811.
- Harris CJ, Slootweg EJ, Goverse A, Baulcombe DC. 2013. Stepwise artificial evolution of a plant disease resistance gene. Proc Natl Acad Sci U S A.. 110: 21189-21194.
- He J, Zhao X, Laroche A, Lu Z-X, Liu H, Li Z. 2014. Genotyping-by-sequencing (GBS), an ultimate marker- assisted selection (MAS) tool to accelerate plant breeding. Front. Plant Sci.. 5: 484
- Hong Y-J, Hossain MR, Kim H-T, Park J-I, Nou I-S. 2018. Identification of two new races of Podosphaera xanthii causing powdery mildew in melon in South Korea. Plant Pathol J.. 34: 182
- Hosoya K, Narisawa K, Pitrat M, Ezura H. 1999. Race identification in powdery mildew (Sphaerotheca fuliginea) on melon (Cucumis melo) in Japan. Plant Breed.. 118: 259-262.
- Howlader J, Hong Y, Natarajan S, Sumi KR, Kim H-T, Park J-I, et al. 2020. Development of powdery mildew race 5-specific SNP markers in Cucumis melo L. using whole- genome resequencing. Hortic. Environ. Biotechnol.. 61: 347-357.
- Kessuwan K, Kubota S, Liu Q, Sano M, Okamoto N, Sakamoto T, et al. 2016. Detection of growth-related quantitative trait loci and high-resolution genetic linkage maps using simple sequence repeat markers in the kelp grouper (Epinephelus bruneus). Mar. Biotechnol.. 18: 57-84.
- Kim H-t, Park J-i, Ishikawa T, Kuzuya M, Horii M, Yashiro K, et al. 2015. Development of molecular marker to select resistant lines and to differentiate the races related to powdery mildew in melon (Cucumis melo L.). Plant Biotechnol. J.. 42: 284-289.
- Kim H-tPark J-iNou I-s2016a. Identification of fungal races that cause powdery mildew in melon (Cucumis melo L.) and selection of resistant commercial melon cultivars against the identified races in Korea. Plant Biotechnol. J.. 43: 58-65.
- Kim H-T, Park J-I, Robin AHK, Ishikawa T, Kuzuya M, Horii M, et al. 2016b. Identification of a new race and development of DNA markers associated with powdery mildew in melon. Plant Breed Biotechnol.. 4: 225-233.
- Kota R, Rudd S, Facius A, Kolesov G, Thiel T, Zhang H, et al. 2003. Snipping polymorphisms from large EST collections in barley (Hordeum vulgare L.). Mol. Genet. Genom.. 270: 24-33.
- Kourelis J, Van Der Hoorn RA. 2018. Defended to the nines: 25 years of resistance gene cloning identifies nine mecha-nisms for R protein function. Plant Cell. 30: 285-299.
- Kuzuya M, Yashiro K, Tomita K. 2004. Melon [Cucumis melo] breeding for resistance to powdery mildew in respect to its races. Proceedings of Vegetable and Tea Science (Japan)..
- Lebeda A, McCreight JD, Coffey MD. Křístková ESedláková B2016. Cucurbit powdery mildews: methodology for objective determination and denomination of races. Eur. J. Plant Pathol.. 144: 399-410.
- Lee JH, Jang KS, Lee WJ, Choi YH, Choi GJ. 2014. Resistance of cucurbits to Podosphaera xanthii race 1. Korean J. Hortic. Sci. Technol.. 32: 673-683.
- Lester GE. 2008. Antioxidant, sugar, mineral, and phytonutrient concentrations across edible fruit tissues of orange- fleshed honeydew melon (Cucumis melo L.). J. Agric. Food Chem.. 56: 3694-3698.
- Li B, Zhao Y, Zhu Q, Zhang Z, Fan C, Amanullah S, et al. 2017. Mapping of powdery mildew resistance genes in melon (Cucumis melo L.) by bulked segregant analysis. Sci. Hortic.. 220: 160-167.
- Li C, Bai G, Carver BF, Chao S, Wang Z. 2015. Single nucleotide polymorphism markers linked to QTL for wheat yield traits. Euphytica. 206: 89-101.
- Liu L, Chen Y, Su Z, Zhang H, Zhu W. 2010. A sequence- amplified characterized region marker for a single, domi-nant gene in melon PI 134198 that confers resistance to a unique race of Podosphaera xanthii in China. HortScience. 45: 1407-1410.
- Mackay JF, Wright CD, Bonfiglioli RG. 2008. A new approach to varietal identification in plants by micro-satellite high resolution melting analysis: application to the verification of grapevine and olive cultivars. Plant Methods. 4: 8
- Mahfooz S, Singh SP, Rakh R, Bhattacharya A, Mishra N, Singh PC, et al. 2016. A comprehensive characterization of simple sequence repeats in the sequenced Trichoderma genomes provides valuable resources for marker develop-ment. Front. Microbiol.. 7: 575
- Mammadov J, Aggarwal R, Buyyarapu R, Kumpatla S. 2012. SNP markers and their impact on plant breeding. Int J Plant Genomics.. 2012: 11
- McCreight JD. 2006. Melon-powdery mildew interactions reveal variation in melon cultigens and Podosphaera xanthii races 1 and 2. J. Am. Soc. Hortic. Sci. . 131: 59-65.
- Meyers BC, Kozik A, Griego A, Kuang H, Michelmore RW. 2003. Genome-wide analysis of NBS-LRR–encoding genes in Arabidopsis. Plant Cell. 15: 809-834.
- Monosi B, Wisser R, Pennill L, Hulbert S. 2004. Full-genome analysis of resistance gene homologues in rice. Theor. Appl. Genet.. 109: 1434-1447.
- Natarajan S, Kim H-T, Thamilarasan SK, Veerappan K, Park J-I, Nou I-S. 2016. Whole genome re-sequencing and characterization of powdery mildew disease-associated allelic variation in melon. PloS one. 11: e0157524
- Ning X, Wang X, Gao X, Zhang Z, Zhang L, Yan W, et al. 2014. Inheritances and location of powdery mildew resis-tance gene in melon Edisto47. Euphytica. 195: 345-353.
- Padmanabhan M, Cournoyer P. Dinesh‐Kumar S2009. The leucine‐rich repeat domain in plant innate immunity: a wealth of possibilities. Cell. Microbiol.. 11: 191-198.
- Perchepied L, Bardin M, Dogimont C, Pitrat M. 2005. Relationship between loci conferring downy mildew and powdery mildew resistance in melon assessed by quanti-tative trait loci mapping. Phytopathology. 95: 556-565.
- Tanksley SD, Young ND, Paterson AH, Bonierbale M. 1989. RFLP mapping in plant breeding: new tools for an old science. technology. 7: 257-264.
- Thomson MJ. 2014. High-throughput SNP genotyping to accelerate crop improvement. Plant Breed. Biotech.. 2: 195-212.
- Polat I, YILDIRIM A, Onus AN. ÜNLÜ A2022. Mapping quantitative trait loci and developing first molecular marker for race 5 of Podosphera xanthii resistance in melon (Cucumis melo L.). Turk. J. Bot.. 46: 123-133.
- Vossen RH, Aten E, Roos A, den Dunnen JT. 2009. High‐Resolution Melting Analysis (HRMA)—More than just sequence variant screening. Hum. Mutat.. 30: 860-866.
- Wan H, Yuan W, Bo K, Shen J, Pang X, Chen J. 2013. Genome-wide analysis of NBS-encoding disease resis-tance genes in Cucumis sativus and phylogenetic study of NBS-encoding genes in Cucurbitaceae crops. BMC Genom.. 14: 1-15.
- Wanga MA, Shimelis H, Mashilo J, Laing MD. 2021. Opportunities and challenges of speed breeding: A review. Plant Breed.. 140: 185-194.
- Watson A, Ghosh S, Williams MJ, Cuddy WS, Simmonds J, Rey MD, et al. 2018. Speed breeding is a powerful tool to accelerate crop research and breeding. Nat. Plants.. 4: 23-29.
- Yang H, Li C, Lam HM, Clements J, Yan G, Zhao S. 2015. Sequencing consolidates molecular markers with plant breeding practice. Theoretical and Applied Genetics. 128: 779-795.
- Yuste-Lisbona FJ, Capel C, Capel J, Lozano R. Gómez-Guillamón MLLópez-Sesé AI2011. Codominant PCR-based markers and candidate genes for powdery mildew resistance in melon (Cucumis melo L.). Theor. Appl. Genet.. 122: 747-758.
- Yuste‐Lisbona FLópez‐Sesé AGómez‐Guillamón M2010. Inheritance of resistance to races 1, 2 and 5 of powdery mildew in the melon TGR‐1551. Plant Breed.. 129: 72-75.
- Zhang C, Ren Y, Guo S, Zhang H, Gong G, Du Y, et al. 2013. Application of comparative genomics in developing markers tightly linked to the Pm-2F gene for powdery mildew resistance in melon (Cucumis melo L.). Euphytica. 190: 157-168.