Abstract
Polyploid plants, such as hexaploid Solanum nigrum, present new challenges and opportunities for trait improvement due to their complex genome structures and homoeologous gene interactions. Gene editing of homoeologs provides a powerful tool to investigate the inheritance and phenotypic effects of mutations in polyploid systems. Here, we investigated the inheritance and phenotypic traits of homoeolog-edited triple and double mutants in hexaploid S. nigrum. Triple mutant genotypes (sns and snsp) were used for analyzing inheritance patterns through backcrossing and self-pollination, revealing independent segregation of homoeologous variants and consistent genotypic ratios across generations. Yield and related traits were evaluated in fixed double mutants (sns1 sns2, sns1 sns3, sns2 sns3) under greenhouse and open-field conditions. Open-field cultivation significantly enhanced yield, primarily through increased fruit weight per inflorescence, whereas greenhouse cultivation showed more modest improvements. Other traits such as plant weight and Brix values remained largely unchanged. Yield improvements were stable across F3 to F5 generations, with minimal genotype-specific variability. These findings highlight the potential of homoeolog-targeted editing in polyploid plants to achieve stable and environment-specific yield enhancements.
-
Key words: Hexaploid, S. nigrum, Inheritance, Homoeolog editing, Yield traits
Introduction
However, polyploid inheritance is more complex than that of diploids. In polyploid genomes, homoeologous genes, which are derived from different ancestral species, interact dynamically to maintain functional balance. This phenomenon, often referred to as subgenome dominance, underscores the complexity of editing polyploid genomes (
Bird et al. 2018). Targeted homoeolog editing not only introduces novel allele combinations but also affects dose sensitivity, a critical factor in phenotypic expression. Dose sensitivity arises from the varying contribution of multiple homoeologs to specific traits, where the loss or modification of one homoeolog can be compensated—or exacerbated—by others (
Birchler et al. 2012). This interaction can result in non-linear phenotypic effects, particularly in traits governed by gene dosage (
Lee et al. 2023).
Polyploid inheritance is shaped by interactions between subgenomes, which contribute to phenotypic stability and adaptability (
Comai 2005). A phenomenon known as "subgenome dominance," where one subgenome shows higher gene expression levels, can influence trait inheritance and shift over generations, affecting breeding outcomes (
Wang et al. 2022). Subgenomic interactions, including those between homologous and homoeologous genes, create novel allele combinations that enhance phenotypic diversity and impact traits like disease resistance and stress tolerance (
Deb et al. 2023). However, polyploid genomes often exhibit genetic redundancy, where multiple homoeologous copies buffer the effects of mutations, obscuring their contributions (
De Smet et al. 2012). Additionally, gene dosage effects and subgenome interplay can lead to unexpected phenotypic outcomes, complicating the interpretation of inheritance patterns (
Lee et al. 2023). This complexity underscores the need for focused analyses of genetic and phenotypic outcomes in polyploid crops to optimize trait selection.
Despite advance in genome editing tools, significant challenges remain in applying CRISPR/Cas technology to polyploid crops, particularly in effectively utilizing mutations after targeting multiple genes (homoeologs). This underscores the need for further studies to elucidate the complex inheritance patterns of homologs and homoeologs in polyploid genomes. Furthermore, verifying the inheritance and fixation of multiple mutated genotypes across generations necessitates yield and phenotypic analysis at each stage. This study aims to systematically examine the inheritance patterns of mutation-induced genotypes created through gene editing in hexaploid S. nigrum, focusing on how these genotypes are passed down across generations.
Materials and Methods
Plant material
S. nigrum (accession NIBRGR0000189638) were used in this study (
Heo et al. 2022). The seeds of
sns and
snsp mutant series were obtained from those developed by (
Lee et al. 2023). Plants were grown in a greenhouse under long-day conditions (14-16 hours of light at 26-28℃ and 8-10 hours of dark at 18-20℃, with 40%-60% relative humidity) supplemented with artificial light from 200 W halogen lamps. Seeds were directly sown in the soil in 50-cell plastic flats in greenhouse. Seedlings that were 4 weeks old after germination were transplanted to pots in the greenhouse and to the field.
Plant genotyping
Deep sequencing, as previously described (
Oh et al. 2020), was employed to identify CRISPR/Cas9-induced mutations in the T0 transformants. Briefly, Deep sequencing was performed using Illumina MiniSeq (Illumina, USA) with amplicons carrying indices and sequencing adaptors, which were attached through three rounds of PCR. The mutation alleles were analyzed using the Cas-Analyzer web tool, which is part of the CRISPR RGEN Tools (
https://www.rgenome.net/). From the BC1F2 generation onward, the genotypes of the
sns mutants were verified using the Cleaved Amplified Polymorphic Sequences (CAPS) method (
Konieczny et al. 1993), which utilizes the Xcm1 recognition site specifically localized in gRNA2 of
SnS. The target sites were amplified by PCR using primers that were specific to each of the three
S genes and capable of amplifying the XcmI recognition site. The PCR product was digested with XcmI, where the WT was cleaved by the enzyme, while the mutant remained undigested, allowing for genotyping. The
snsp mutants contains large deletions in each gene. To validate the genotype, PCR was performed using primers specific to each
SnSP. The PCR products were separated by size using 3% agarose gel electrophoresis.
The S. nigrum yield trial was conducted under two different cultivation conditions. Under the first condition, plants were grown in a greenhouse, where pruning was performed to retain only two main shoots, and sufficient nutrients were supplied. Under the second condition, the plants were cultivated in the field without pruning. In the field, organic fertilizer was applied to the soil prior to transplanting, and drip irrigation was implemented afterward. The sns mutant plants were cultivated under both greenhouse and field conditions, while the snsp mutant plants were cultivated under two field conditions.
Plant yield trials
The yield trial was conducted in the Wonkwang University greenhouse (July 2022), the Ochang field (July 2022), and the Sacheon field (July 2024). Harvests were performed when the majority of plants in a trial had 70% black fruit. Fruit yield was estimated through a calculation that multiplied the average weight of inflorescences by the total number of inflorescences. The plant weight, including the fruits but excluding the roots, was measured. Only inflorescences containing black fruits were counted to determine the total number of inflorescences. Brix and 10-fruit weight were measured using randomly selected fruits. Plants that were damaged or affected by disease were excluded from the analysis.
Statistical analyses
Statistical calculations were performed using Prism software 10 (GraphPad Software, San Diego, CA, USA). Statistical analyses were carried out using a one-way analysis of variance (ANOVA) followed by Tukey's correction for multiple comparisons. Different letters indicate significant differences between groups at a significance level of p<0.05.
Phenotyping
When more than 70% of the fruits on the plant had matured, the inflorescences and plant shoots were photographed using an EOS850D camera (Canon, Tokyo, Japan). Representative plants were selected to effectively illustrate the phenotypic characteristics of the sns mutants, which exhibit an increase in inflorescence branches, and the snsp mutants, which show a reduction in internode length between inflorescences.
Results
Inheritance study of co-edited homoeolog triple mutants in hexaploid S. nigrum
In a previous study, we identified 8911 homoeolog groups of three copies in the hexaploid
S. nigrum genome, and we obtained T0 generation of co-edited
sns and
snsp triple mutants (
Lee et al. 2023;
Park et al. 2020). To analyze the inheritance of the mutations induced in the hexaploid
S. nigrum homoeolog group, we first performed backcrosses of
sns triple mutants with wild type in the T0 generation to determine the pattern of inheritance of each of the genetic variants, and confirmed the segregation of each homoeolog variant in the F2 generation through self-pollination of F1 generation individuals. We first identified the triple
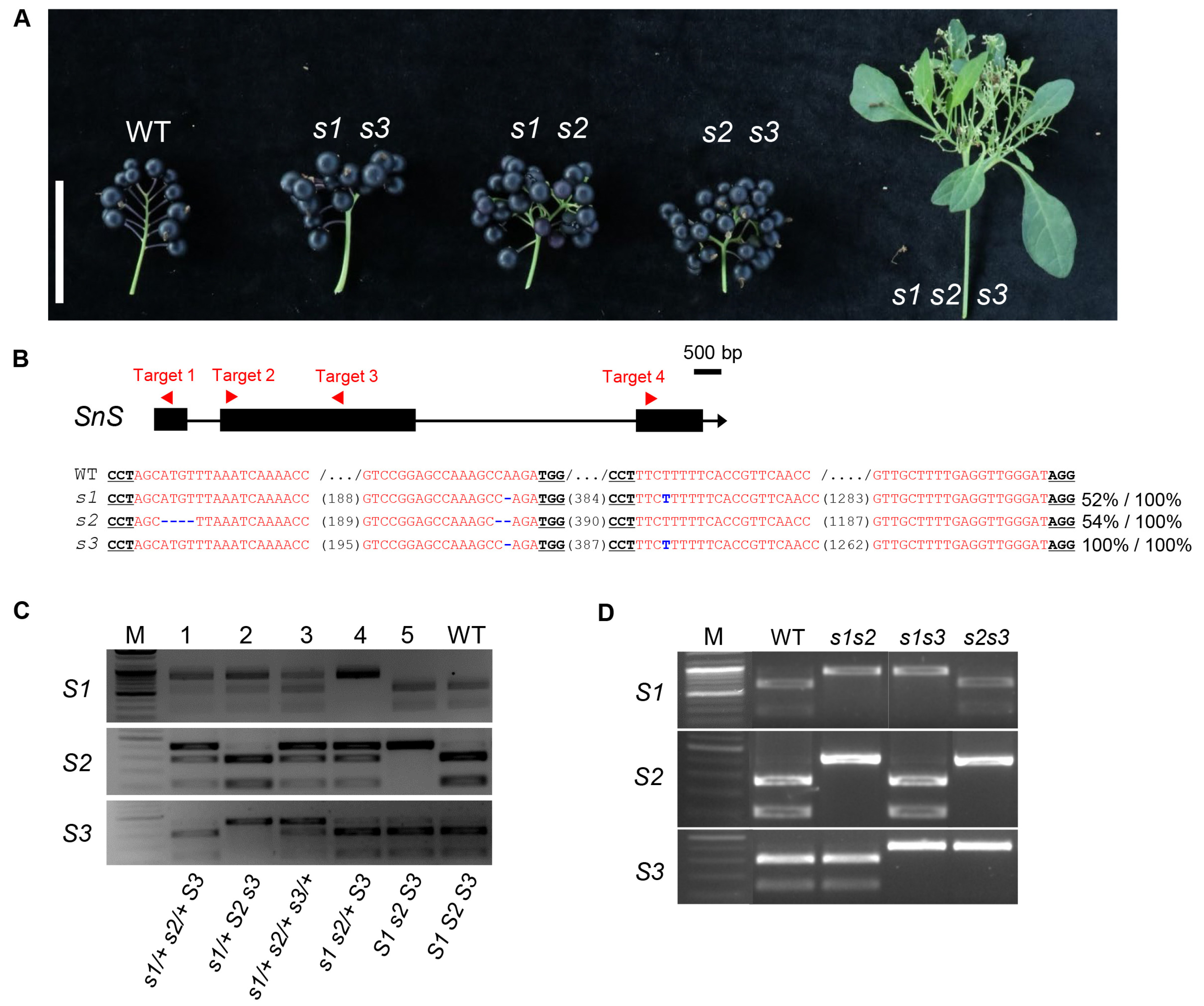
sns mutation in the T0 generation by amplicon sequencing, selecting three T0 individuals with >95% sum of mutation alleles induced in each homoeologs (
Fig. 1). The three T0 individuals were then backcrossed to the wild type, and one F1 plant was selected in which the T-DNA insert was removed and all three homoeologs were heterozygous with the deletion allele (
Kim et al. 2022). Subsequently, 237 F2 seeds obtained through self-pollination of selected F1 plant were sown to determine the genotypic segregation ratio of the variants derived from the three homoeologs, which showed a ratio of approximately 1:2:1 genotype for each allele. Specifically, 82 wild-type, 117 heterozygous, and 38 homozygous mutants were analyzed for
sns1; 72 wild-type, 111 heterozygous, and 54 homozygous mutants were analyzed for
sns2; and 52 wild-type, 129 heterozygous, and 56 homozygous mutants were analyzed for
sns3. In addition, 4, 4, and 2 double mutants were obtained for
sns1 sns2,
sns1 sns3, and
sns2 sns3, respectively, and 0 for
sns1 sns2 sns3 were analyzed (
Table 1). Notably, the knock-out mutant of the diploid tomato
SnS ortholog was reported to exhibit an embryo-defective phenotype, suggesting that
sns1 sns2 sns3 may also display similar defects in embryo development (
Hendelman et al. 2021).
We also conducted a genetic segregation ratio analysis by genotyping the F2 population through backcrossing wild type with T0 generation of
snsp-edited triple mutants followed by self-pollination of the F1 generation (
Supplementary Fig. 1). The
snsp mutation rate, segregated from the F2 generation, was found to be 16.4% wild-type, 57.5% heterozygous, and 26.1% homozygous mutants in
snsp1; 23.7% wild-type, 50.5% heterozygous, and 25.8% homozygous mutants in
snsp2; and 26.8% wild-type, 50.5% heterozygous, and 22.7% homozygous mutants in
snsp3. The
snsp double mutation rate was also similar each other, such as 0.6% in
snsp1 snsp2, 1% in
snsp1 snsp3, and 0.6%
in snsp2 snsp3, and the triple mutation rate was 2% (
Table 1). These results indicate that each of the gene-editing-induced homoeologous mutant genotypes is being inherited independently in hexaploid. To determine whether the fixed homozygous mutant genotypes were maintained in the F3 generation, a series of three double-mutant progenies were sown to determine the genotypes of eight individuals each, and we found that the genotypes remained the same as those in the parental generation (
Fig. 1).
To investigate the impact of environmental conditions on the phenotypic traits of F3 fixed double mutants (
sns1 sns2,
sns1 sns3,
sns2 sns3), we evaluated their fruit yields under two cultivation systems: greenhouse (hydroponics with pruning) and open-field (fertilization without pruning; see methods). Seeds were sown in March, the optimal growing season for
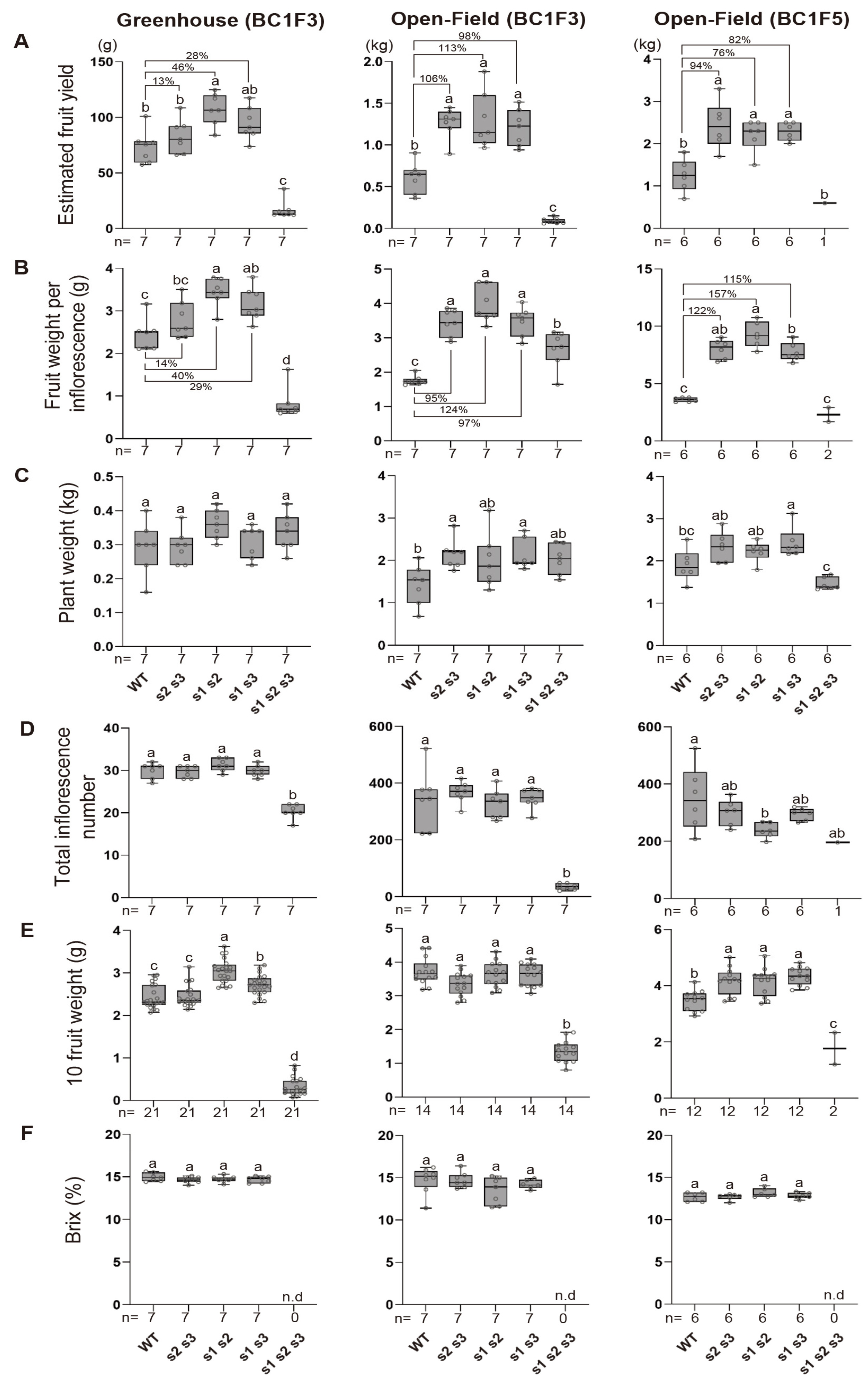
S. nigrum in South Korea, and fruit yields were measured in July. The open-field system significantly boosted yields, with all double mutants achieving over a 100% increase compared to wild-type controls. Although yield improvements varied among genotypes, all double mutants demonstrated substantial enhancements (
Fig. 2A). Under greenhouse conditions, the double mutants of
S. nigrum also produced higher fruit yields compared to the wild type. However, the yield increase was less pronounced than under the open field conditions. Among the double mutants,
sns1 sns2 exhibited the highest yield enhancement, achieving approximately an 46% increase over the wild type. Interestingly,
sns2 sns3 demonstrated higher average yields in greenhouse conditions than the one of wild type, but the increase was not statistically significant (
Fig. 2A).
Major traits contributing to the yield improvement of
S. nigrum include fruit weight per inflorescence, plant weight, total inflorescence number, and total fruit weight estimated. We measured these traits under both greenhouse and open-field conditions, using the same settings as the yield assessment, and compared the differences between the two cultivation systems. Interestingly, the fruit weight per inflorescence in the double mutants was significantly higher than in the wild type under both cultivation systems (
Fig. 2B). When comparing the trait improvement ratios relative to the wild type, the fruit weight per inflorescence showed a much greater increase in open-field conditions than in the greenhouse (
Fig. 2B). For example, in the
sns2 sns3, the fruit weight per inflorescence increased by 14% under greenhouse conditions but dramatically rose to 95% under open-field conditions. Similar patterns were observed in the other two double mutants (
Fig. 2B). Other yield-related traits, including plant weight, total inflorescence number, and 10-fruit weight, showed no significant differences in the double mutants compared to the wild type, regardless of the growing conditions (
Figs. 2C-
2E). Similarly, Brix values, which indicate sugar accumulation in the fruit, remained consistent across all genotypes and cultivation environments, showing no variation from the wild type (
Fig. 2F).
Taken together, these results indicate that the primary factor contributing to the enhanced yield in sns-edited variants of hexaploid S. nigrum is the increase in fruit weight per inflorescence. Additionally, the findings confirm that yield and yield-related traits in these edited variants can vary depending on the cultivation environment.
Inheritance of phenotypic traits across generations
To investigate whether fixed homoeolog mutations in polyploid
S. nigrum lead to phenotypic differences across generations, we compared the changes in yield and yield-related traits in the F3 and F5 generations of homoeolog-fixed double mutants (
sns1 sns2,
sns1 sns3,
sns2 sns3). Compared to the control, the yield improvement in F3 double mutants was similar to that observed in the F5 double mutants (
Fig. 2A). All three double mutant genotypes demonstrated approximately a 82% increase in yield compared to the wild type, with consistent yield enhancement ratios across both F3 and F5 generations of the double mutants (
Fig. 2A). Notably, there was minimal variability among individuals within the same genotype across generations. The yield variability among the three double mutant genotypes was measured as 0.18 in the F3 generation and 0.28 in the F5 generation, indicating that phenotypic traits related to yield were well-preserved across generations (
Table 2).
One of the most critical factors affecting yield, the fruit weight per inflorescence, showed significantly higher values in double mutants than the wild type, with consistent patterns in both the F3 and F5 generations (
Fig. 2B). Other yield-related traits, such as plant weight, total inflorescence number, 10-fruit weight, and Brix values, showed no significant changes in the double mutants compared to the wild type, nor were there substantial differences between generations (
Figs. 2C-
2F).
Similarly, we analyzed the yield and yield-related traits of
snsp1 snsp2 and
snsp1 snsp3 double mutants across the F3 and F4 generations under open-field conditions. These
snsp double mutants consistently displayed semi-determinate stem growth traits in both generations (
Supplementary Fig. 1). Traits such as fruit yield, plant weight, and Brix values showed minimal differences compared to wild-type ones. However, the triple mutant
snsp1 snsp2 snsp3 exhibited a significant reduction in these traits. The overall variation patterns among genotypes were highly consistent across F3 and F4 generations (
Supplementary Fig. 2).
In conclusion, the inheritance of mutations in hexploid S. nigrum resulted in consistent phenotypic traits across generations. However, yield-related traits under specific environmental conditions were influenced by the additive or partially lost functions of homoeologs, which might lead to increased variability in yield-related traits depending on the combination of mutant genotypes.
Discussion
This study investigates the inheritance patterns and phenotypic traits of homoeolog-edited mutants in hexaploid S. nigrum, focusing on the segregation of mutations and their impact on yield-related traits. The findings provide valuable insights into the potential of homoeolog-targeted editing for crop improvement and the stability of phenotypic expression across generations.
The results demonstrate that homoeologous mutations in
S. nigrum segregate independently and follow Mendelian ratios in the F2 generation, as evidenced by the approximately 1:2:1 genotypic segregation ratio of wild-type, heterozygous, and homozygous mutants across all three homoeologs (
sns1,
sns2,
sns3). These inheritance patterns confirm that targeted mutagenesis using CRISPR/Cas technologies can achieve stable transmission of edited alleles in polyploid crops. Furthermore, the consistency of genotypes in the F3 generation highlights the robustness of gene editing in creating stable mutant lines suitable for breeding programs. Similar findings have been reported in other polyploid crops, such as wheat, where homoeolog editing has enabled the development of stable traits through precise allele manipulation (
Howells et al. 2018;
Zhang et al. 2019).
The study reveals that yield-related traits, including fruit weight per inflorescence, are consistently enhanced in
sns homoeologous double mutants across the F3 and F5 generations, with approximately a 100% yield increase compared to the wild type. This phenotypic stability underscores the reliability of targeted homoeolog editing in maintaining desirable traits over multiple generations. Importantly, the minimal variability observed within and between genotypes across generations suggests that these traits are well-preserved and relatively unaffected by potential genetic recombination. These findings align with previous studies demonstrating stable phenotypic expression in polyploid crops with fixed homoeolog mutations (
He et al. 2022;
Lee et al. 2023;
Mathur et al. 2021).
Environmental conditions significantly influenced the expression of yield-related traits, particularly under open-field and greenhouse cultivation systems. While both systems showed increased yields in
sns homoeologous double mutants compared to the wild type, the open-field system resulted in a greater enhancement than greenhouse growth system, primarily driven by increased fruit weight per inflorescence. This indicates that certain environmental factors, such as fertilization and pruning practices, may modulate the expression of yield-enhancing traits. The variability in trait expression across environments highlights the importance of considering genotype-by-environment interactions in polyploid breeding programs. For instance,
sns2 sns3 demonstrated a dramatic increase in total fruit yield under open-field conditions but only a modest enhancement in greenhouse growth condition. This finding is consistent with studies in polyploid wheat and potato, where environmental factors were shown to influence trait expression in edited genotypes (
Qu et al. 2024;
Zhao et al. 2022).
The study confirms that homoeolog-targeted editing can generate stable and predictable trait improvements in hexaploid S. nigrum. However, the findings also underscore the complexity of polyploid breeding, where interactions between environmental factors can lead to variable outcomes. The observed consistency in phenotypic expression across generations provides a strong foundation for integrating gene-edited mutants into breeding programs. Furthermore, the potential for environment-specific trait enhancements offers opportunities for optimizing crop performance under diverse agricultural conditions. Future research could focus on dissecting the molecular mechanisms underlying trait stability and environmental responsiveness in edited polyploid crops. Additionally, expanding the scope of homoeolog editing to include other yield-related traits, such as stress tolerance and disease resistance, could further enhance the utility of this approach in polyploid breeding.
Acknowledgments
This work was supported by a grant from the New Breeding Technologies Development Program (RS-2024-00322420) to S.J.P., and also supported by the National Research Foundation of Korea grant numbers (RS- 2024-00356396, 2020R1A6A1A03044344) E.S.L. and H.Y.K., also supported by the Glocal University 30 Project Fund of Gyeongsang National University in 2024.
Fig. 1Inheritance of segregants backcrossed from co-edited s triple mutant in hexaploid S. nigrum. (A) Inflorescence branching of the s double and triple mutants from the BC1F5 population. The triple mutant rarely develops mature black fruits from flowers. Size bar, 5 cm. (B) CRISPR-Cas9-generated mutations in gene loci of s triple mutant used for backcrossing. The gene structure and four sgRNA targeting sites are depicted in the upper diagram. sgRNA-targeted sequences are highlighted in red. Each protospacer- adjacent motif (PAM) sequence is underlined. Numbers in parentheses represent gap lengths between sgRNAs. Each mutation is denoted in blue. Percent values indicate the proportion of each allele relative to the total. (C) Example of genotyping by cleaved amplified polymorphic sequence (CAPS) markers of segregants from the BC1F2 population. (D) Example of genotyping by CAPS markers of s double mutants from the BC1F3 population. M, DNA size marker. WT, wild type.
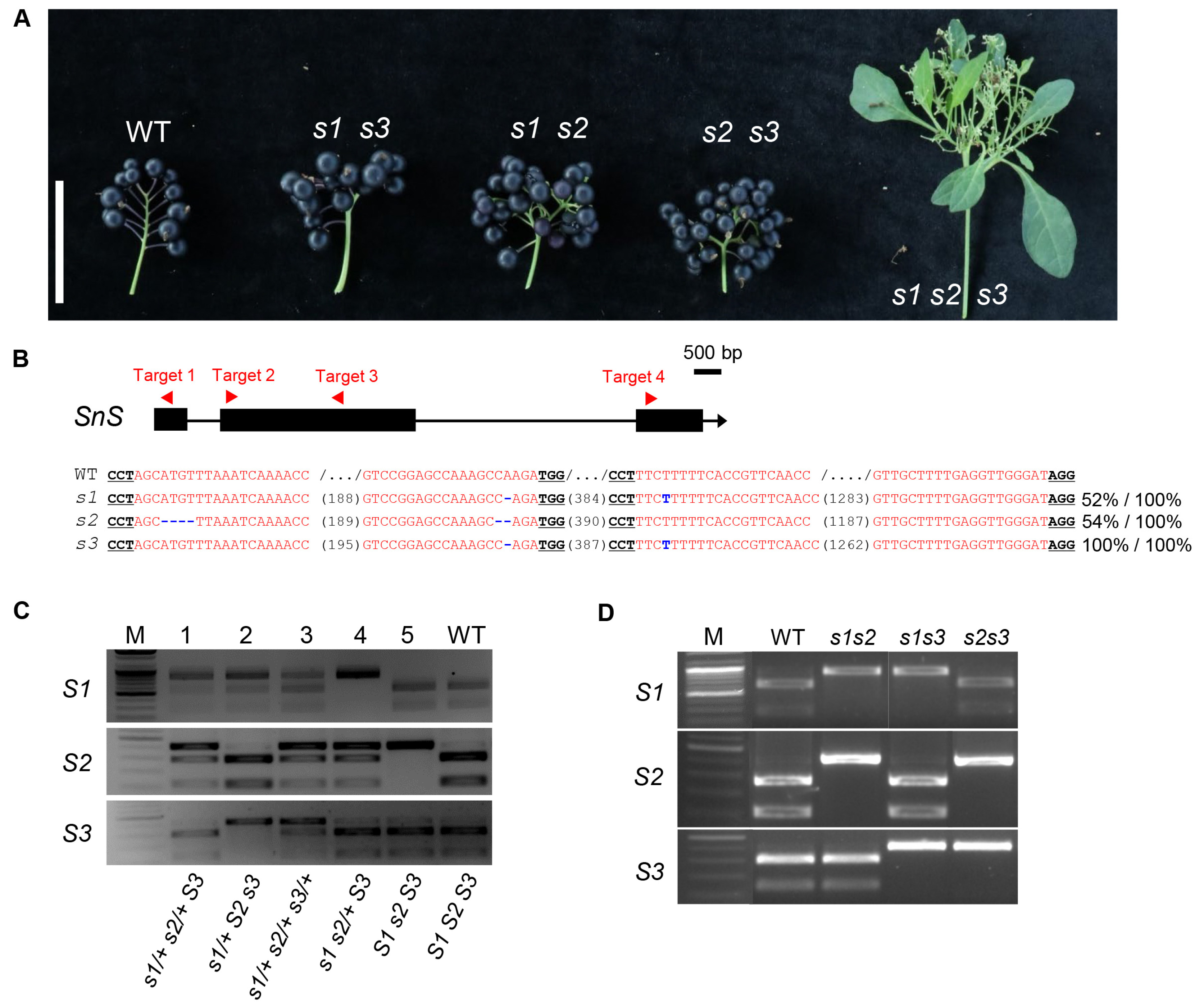
Fig. 2Productivity trials of the inherited s mutants under greenhouse and open-field conditions. (A-F) Estimated fruit yield, fruit weight per inflorescence, plant weight, total inflorescence number, 10 fruit weight, and Brix were measured in WT, s double and triple mutants. n represents the number of plants. Percent value indicates the extent of the increase in the trait of the double mutant compared to the WT. Box plot depicts the 25th, 50th, and 75th percentiles, and different letters indicate statistically significant differences. The whiskers indicate all data points (One-way ANOVA, p<0.05).
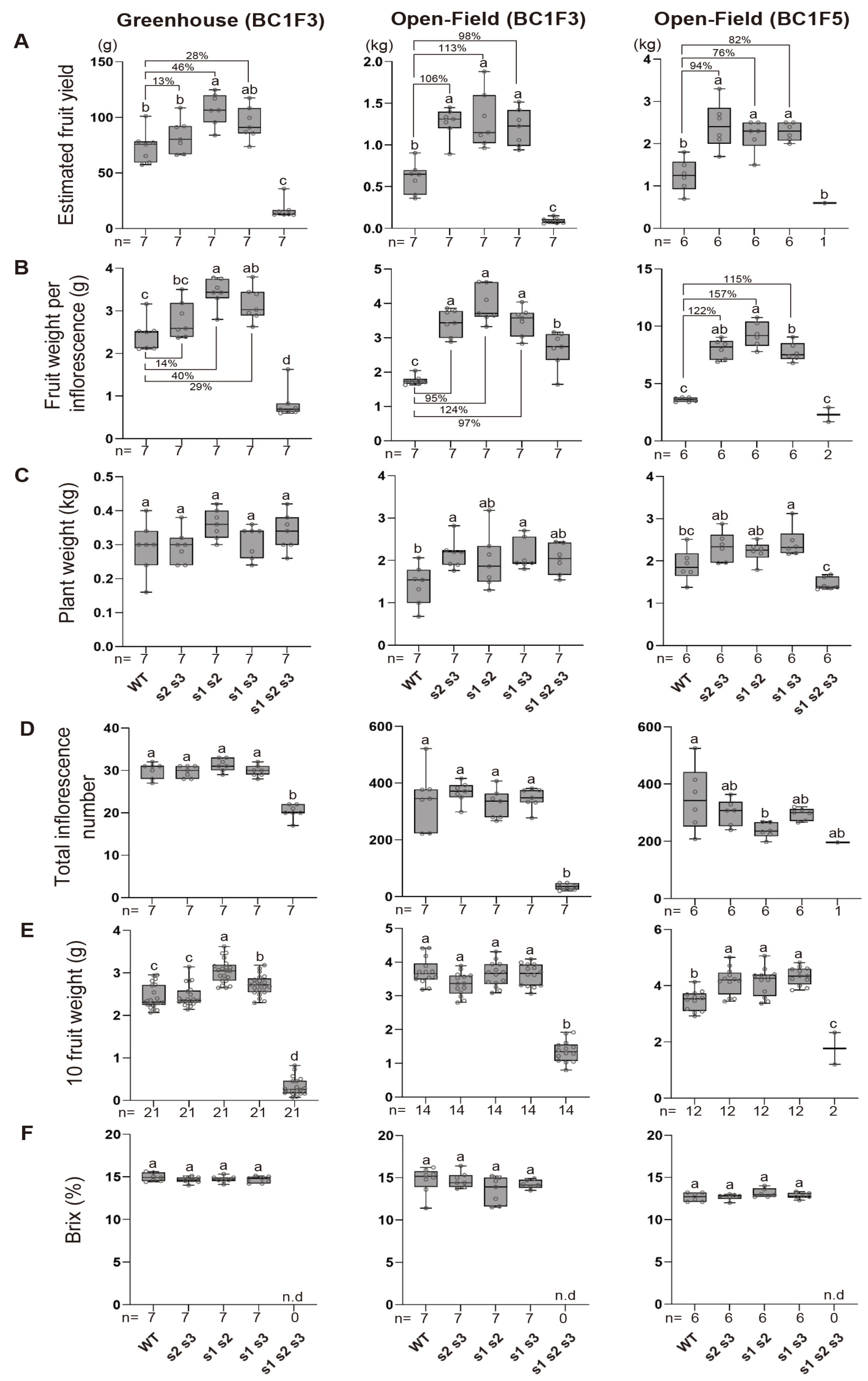
Table 1Distribution of genotypes and allelic proportions in homoeologous segregation
Table 1
|
Genotype |
Independent segregation in single homoeologue |
|
Segregation of homozygote alelle in three homoeologs |
Total
plants |
|
A1A1 / A1a1 / a1a1 |
A2A2 / A2a2 / a2a2 |
A3A3 / A3a3 / a3a3 |
A1A1A2A2A3A3 |
a1a1a2a2A3A3 |
a1a1A2A2a3a3 |
A1A1a2a2a3a3 |
a1a1a2a2a3a3 |
|
SnS |
82(34.6)z/ 117(49.4) / 38(16) |
72(30.4) / 111(46.8) / 54(22.8) |
52(22) / 129(54.4) / 56(23.6) |
|
3(1.2) |
4(1.6) |
4(1.6) |
2(0.8) |
0(0) |
237 |
|
SnSP |
49(16.4) / 172(57.5) / 78(26.1) |
71(23.7) / 151(50.5) / 77(25.8) |
80(26.8) / 151(50.5) / 68(22.7) |
|
4(1.3) |
2(0.6) |
3(1) |
2(0.6) |
6(2) |
299 |
Table 2Variability of standard deviation (SD) in fruit yield and yield-related traits in BC1F3 and BC1F5 populations
Table 2
|
Trait |
Cultivation condition |
WT |
s2 s3 |
s1 s2 |
s1 s3 |
s1 s2 s3 |
Variability of double mutantsz |
|
Estimated fruit yield (kg) |
BC1F3_Greenhouse |
73.52±13.76 |
83.15±14.06 |
107.67±13.27 |
94.63±13.76 |
16.81±7.88 |
13.70 |
|
BC1F3_Open-Field |
0.61±0.17 |
1.26±0.17 |
1.3±0.31 |
1.21±0.2 |
0.09±0.03 |
0.18 |
|
BC1F5_Open-Field |
1.25±0.35 |
2.43±0.51 |
2.2±0.34 |
2.28±0.2 |
0.6±0 |
0.28 |
|
|
Fruit weight per inflorescence (g) |
BC1F3_Greenhouse |
2.44±0.35 |
2.79±0.4 |
3.43±0.3 |
3.16±0.37 |
0.82±0.34 |
0.35 |
|
BC1F3_Open-Field |
1.76±0.13 |
3.44±0.36 |
3.95±0.47 |
3.48±0.38 |
2.67±0.49 |
0.37 |
|
BC1F5_Open-Field |
3.6±0.17 |
8.01±0.78 |
9.28±1.01 |
7.75±0.74 |
2.3±0.62 |
0.66 |
|
|
Plant weight (kg) |
BC1F3_Greenhouse |
0.29±0.07 |
0.29±0.04 |
0.36±0.04 |
0.31±0.04 |
0.34±0.05 |
0.05 |
|
BC1F3_Open-Field |
1.42±0.43 |
2.15±0.32 |
1.99±0.59 |
2.12±0.33 |
2.03±0.32 |
0.40 |
|
BC1F5_Open-Field |
1.9±0.35 |
2.33±0.32 |
2.22±0.22 |
2.44±0.32 |
1.46±0.14 |
0.27 |
|
|
Total inflorescence number |
BC1F3_Greenhouse |
30±1.69 |
29.71±1.28 |
31.29±1.39 |
29.86±1.25 |
20.29±1.58 |
1.44 |
|
BC1F3_Open-Field |
344±94.95 |
367.57±34.69 |
325.43±47.96 |
345.86±33.71 |
34.43±10.24 |
44.31 |
|
BC1F5_Open-Field |
349.67±103.38 |
300.67±41.57 |
237.83±24.37 |
294.67±19.46 |
196±0 |
37.76 |
|
|
10 fruit weight (g) |
BC1F3_Greenhouse |
2.43±0.27 |
2.45±0.25 |
3.04±0.27 |
2.71±0.23 |
0.33±0.21 |
0.25 |
|
BC1F3_Open-Field |
3.72±0.37 |
3.35±0.32 |
3.63±0.35 |
3.63±0.31 |
1.36±0.31 |
0.33 |
|
BC1F5_Open-Field |
3.47±0.34 |
4.17±0.46 |
4.14±0.48 |
4.34±0.3 |
1.77±0.57 |
0.43 |
|
|
Brix (%) |
BC1F3_Greenhouse |
14.93±0.44 |
14.63±0.35 |
14.7±0.35 |
14.66±0.34 |
NA |
0.37 |
|
BC1F3_Open-Field |
14.7±1.45 |
14.73±0.86 |
13.47±1.47 |
14.22±0.47 |
NA |
1.06 |
|
BC1F5_Open-Field |
12.67±0.48 |
12.68±0.33 |
13.15±0.49 |
12.82±0.32 |
NA |
0.41 |
References
- Birchler JA, Veitia RA. 2012. Gene balance hypothesis: connecting issues of dosage sensitivity across biological disciplines. Proc Natl Acad Sci U S A. 109: 14746-14753.
- Bird KA, VanBuren R, Puzey JR, Edger PP. 2018. The causes and consequences of subgenome dominance in hybrids and recent polyploids. New Phytol. 220: 87-93.
- Chen L, Li C, Li B, Zhou X, Bai Y, Zou X, et al. 2024. Evolutionary divergence of subgenomes in common carp provides insights into speciation and allopolyploid success. Fundam Res. 4: 589-602.
- Comai L. 2005. The advantages and disadvantages of being polyploid. Nat Rev Genet. 6: 836-846.
- De Smet R, Van de Peer Y. 2012. Redundancy and rewiring of genetic networks following genome-wide duplication events. Curr Opin Plant Biol. 15: 168-176.
- Deb SK, Edger PP, Pires JC, McKain MR. 2023. Patterns, mechanisms, and consequences of homoeologous exchange in allopolyploid angiosperms: a genomic and epigenomic perspective. New Phytol. 238: 2284-2304.
- He F, Wang W, Rutter WB, Jordan KW, Ren J, Taagen E, et al. 2022. Genomic variants affecting homoeologous gene expression dosage contribute to agronomic trait variation in allopolyploid wheat. Nat Commun. 13: 826
- Hendelman A, Zebell S, Rodriguez-Leal D, Dukler N, Robitaille G, Wu X, et al. 2021. Conserved pleiotropy of an ancient plant homeobox gene uncovered by cis-regulatory dissection. Cell. 184: 1724-1739.e16.
- Heo J, Bang WY, Jeong JC, Park SC, Lee JM, Choi S, et al. 2022. The comparisons of expression pattern reveal molecular regulation of fruit metabolites in S. nigrum and S. lycopersicum. Sci Rep. 12: 5001
- Howells RM, Craze M, Bowden S, Wallington EJ. 2018. Efficient generation of stable, heritable gene edits in wheat using CRISPR/Cas9. BMC Plant Biol. 18: 215
- Kim K, Lee ES, Warsadiharja SM, Lee J, Kang HY, Jang JI, et al. 2024. A new quantitative method to validate the expression of homoeologous genes in polyploids. Plant Biotechnol. Rep. 18: 967-973.
- Kim MJ, Beak HK, Choi JE, Lee ES, Kim K, Kim CM, et al. 2022. Simple methods for selection of T-DNA-free segregants from offspring of gene-edited Solanum nigrum. Plant Biotechnol. Rep. 16: 257-264.
- Konieczny A, Ausubel FM. 1993. A procedure for mapping Arabidopsis mutations using co-dominant ecotype-specific PCR-based markers. Plant J. 4(2): 403-410.
- Lee ES, Heo J, Bang WY, Chougule KM, Waminal NE, Hong NT, et al. 2023. Engineering homoeologs provide a fine scale for quantitative traits in polyploid. Plant Biotechnol J. 21: 2458-2472.
- Li C, Shu Y, Hu Q. 2023. Editorial: Genome editing technology in polyploid crops. Front Plant Sci. 14: 1340455
- Mathur J, Khare PB, Panwar A, Ranade SA. 2021. Analysis of genetic variability amongst polyploid genotypes of pteris vittata L. from various geographic locales of India. Front. Ecol. Evol. https://doi.org/10.3389/fevo.2021.613847.
- Oh Y, Lee B, Kim H, Kim SG. 2020. A multiplex guide RNA expression system and its efficacy for plant genome engineering. Plant Methods. 16: 37
- Park S, Lee E, Heo J, Kim DH, Chun HJ, Kim MC, et al. 2020. Rapid generation of transgenic and gene-edited Solanum nigrum plants using Agrobacterium-mediated transformation. Plant Biotechnol. Rep. 14: 497-504.
- Qu L, Huang X, Su X, Zhu G, Zheng L, Lin J, et al. 2024. Potato: From functional genomics to genetic improvement. Mol Hortic. 4: 34
- Ricroch AS, Ricroch A, Eriksson D, Miladinović D, Sweet J, Van Laere K, et al. 2024. A roadmap for plant genome editing. Springer Nature. p. 561.
- Schaart JG, van de Wiel CCM, Smulders MJM. 2021. Genome editing of polyploid crops: Prospects, achievements and bottlenecks. Transgenic Res. 30: 337-351.
- Wang Z, Yang J, Cheng F, Li P, Xin X, Wang W, et al. 2022. Subgenome dominance and its evolutionary implications in crop domestication and breeding. Hortic Res. 9: uhac090
- Zhang D, Zhang Z, Unver T, Zhang B. 2021. CRISPR/Cas: A powerful tool for gene function study and crop improvement. J Adv Res. 29: 207-221.
- Zhang S, Zhang R, Gao J, Gu T, Song G, Li W, et al. 2019. Highly efficient and heritable targeted mutagenesis in wheat via the agrobacterium tumefaciens-mediated CRISPR/Cas9 system. Int J Mol Sci. 20: 4257
- Zhao J, Sun L, Gao H, Hu M, Mu L, Cheng X, et al. 2022. Genome-wide association study of yield-related traits in common wheat (Triticum aestivum L.) under normal and drought treatment conditions. Front Plant Sci. 13: 1098560