Abstract
Pre-harvest sprouting is a major physiological problem in rice caused by prolonged rainfall and high humidity during the harvest period, and it is one of the most important targets in current rice breeding programs. In this study, the effect of cold and freezing storage on the pre-harvest sprouting rate was investigated using ten rice varieties under four different treatments. The result showed storage treatments of panicle samples used for germinate evaluation had no significant influence on the pre-harvest sprouting rate. These findings may enhance the efficiency of mass screening for pre-harvest sprouting and support the development of tolerant rice varieties.
-
Key words: Pre-harvest sprouting, Cold, Freezing, Storage, Rice
Introduction
Rice (
Oryza sativa L.) is one of the world’s most important food crops, and its quality and yield are affected by various environmental factors during the growing season. Prolonged rainfall and high humidity during the harvest period can induce pre-harvest sprouting (PHS), the premature germination of seeds within the panicle (
Tai et al. 2021). PHS is a serious physiological problem that reduces appearance and milling yield, leading to yield loss (
Kim et al. 2008). Due to its direct impact on yield and quality, it is considered a key trait for rice breeding. Accordingly, a variety of cultivation and breeding approaches to reduce PHS occurrence have been developed, and PHS test methods have been introduced to objectively evaluate their effectiveness (
Baek et al. 2014; Zhao et al. 2022;
Shalem Raju et al. 2024). The conventional PHS test method is generally performed immediately after harvesting panicles at 40 days after heading. However, in large-scale evaluations, the variation in heading dates among samples makes it impractical to maintain a uniform assessment schedule. Under such circumstances, harvested panicles are often stored at room temperature, which may expose them to environmental factors that could alter their pre-harvest sprouting rate prior to evaluation. Therefore, establishing a standardized assessment method that accounts for storage conditions is necessary.
In this study, we evaluated the PHS rates of panicle samples across ten rice varieties to understand the effect of cold and freezing storage treatments. This approach aimed to compare PHS responses of rice varieties under different storage conditions and to evaluate the effectiveness of storage treatment as a means of standardizing PHS testing across varieties.
Materials and Methods
Plant Materials and Cold and Freezing Storage Treatments
Ten japonica rice varieties, including early maturing (‘Boreumi’, ‘MY362VP’, ‘Joil’, ‘Joun’, ‘Jopyeong’, and ‘Baromi2’) and mid-late maturing (‘Namchan’, ‘Nampyeong’, ‘Samkwang’, and ‘Chamdongjin’) were cultivated at the National Institute of Crop and Food Science, Rural Development Administration (Wanju, Republic of Korea) in 2023 and used in this experiments. Rice panicles were harvested at physiological maturity (40 days after heading) and immediately pretreated in a 37℃ dry oven for 48 h to remove surface moisture. To determine the effect of storage treatments on seed germination, panicles were stored under four different conditions: NT (No treatment, without storage), T1 (two weeks at -20℃), T2 (four weeks at -20℃), and T3 (two weeks at 4℃), respectively.
PHS Evaluation
For the PHS test, seven panicles from each variety were selected as biological replicates. Harvest panicles were placed on the steel tray with paper towel, and an upper layer of paper towel was added to cover the panicles. All samples were then incubated in a dew chamber at 25℃ with over 95% relative humidity. After 7 days for incubation, the PHS rate (%) was calculated as (germinated seeds /total filled seeds) × 100 per panicle. Germination was evaluated as the emergence of the coleoptile at least 2 mm through the hull. For each variety, the average PHS was calculated from seven panicles and used for statistical analyses. The varieties with an average PHS rate below 20% were classified as PHS-tolerant group, and varieties showing an average of 20% or higher were classified as PHS-susceptible group. Data values were summarized as mean ± standard deviation across seven replicates in each variety. All statistical analyses were conducted in R, using ANOVA with Tukey’s HSD for post hoc comparisons (
p < 0.05) (
R Core Team, 2020).
Results and Discussion
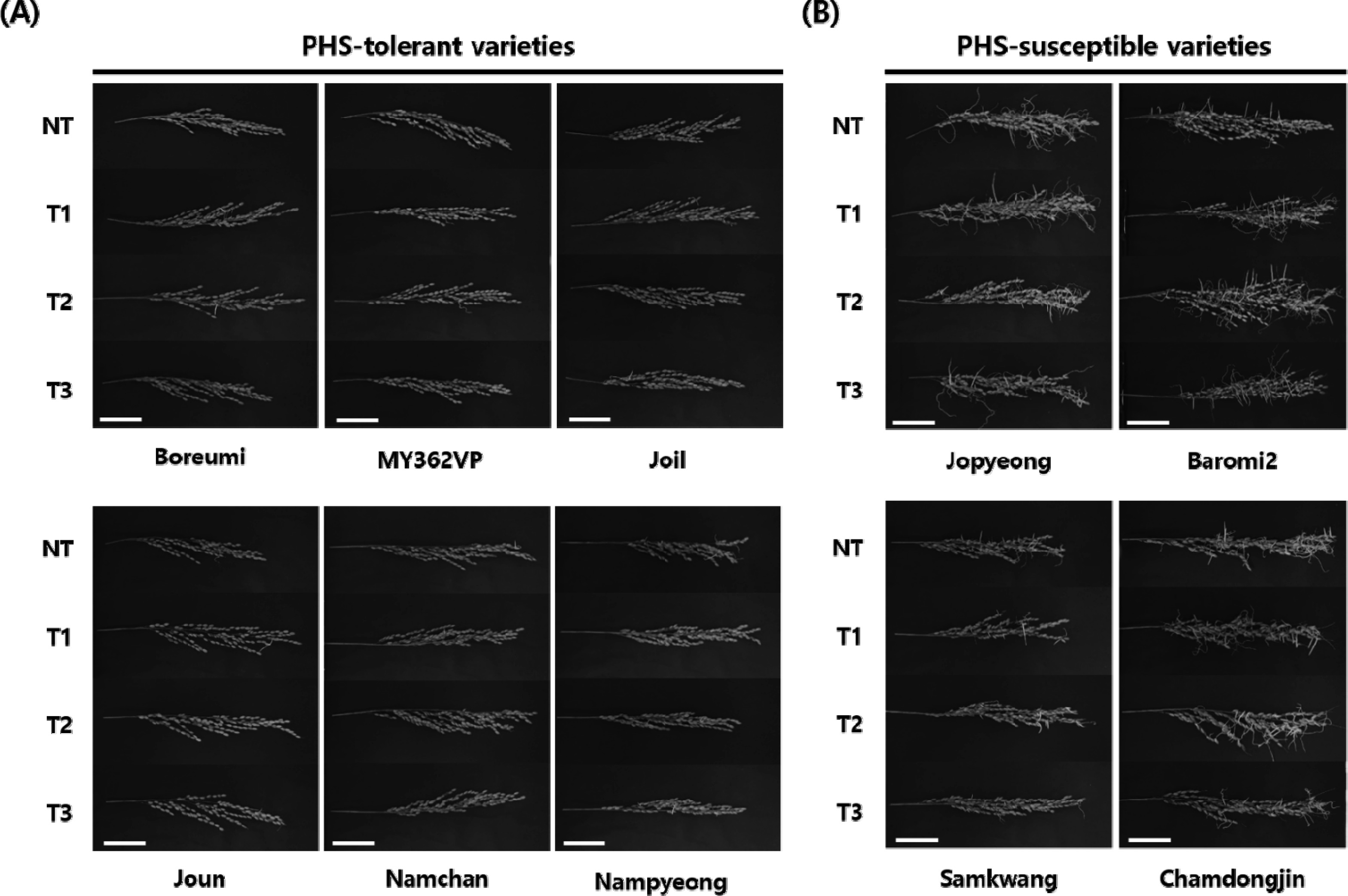
Rice panicles were evaluated under four storage conditions (NT, T1, T2, and T3), and PHS rate was calculated for each variety with condition combination. Ten rice varieties were classified into two phenotypic groups, six PHS-tolerant (‘Boreumi’, ‘MY362VP’, ‘Joil’, ‘Joun’, ‘Namchan’, and ‘Nampyeong’) and four PHS-susceptible (‘Jopyeong’, ‘Baromi2’, ‘Samkwang’, and ‘Chamdongjin’), respectively (
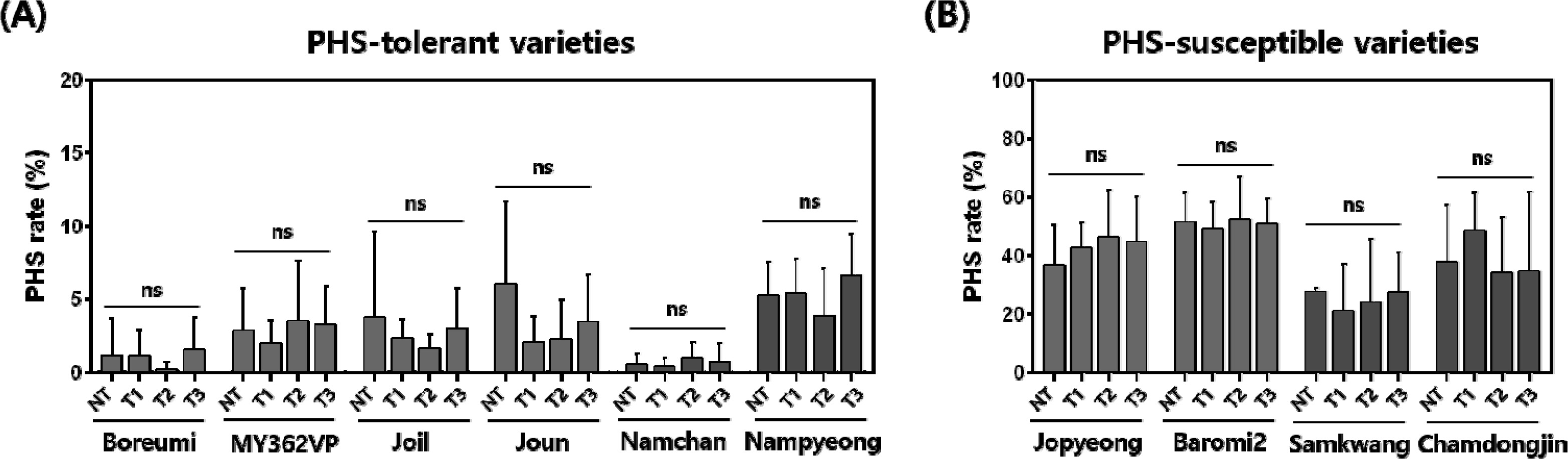
Fig. 1). All varieties showed PHS rate ranged from 0.5% to 51.7% under the NT condition. The average of PHS rate in ‘Baromi2’ showed the highest PHS (49.3–52.7%), while ‘Namchan’ showed the lowest PHS (0.4–1.0%) among all treatments (
Table 1). In the statistics analysis, a two-way ANOVA revealed that varieties had a highly significant effect on PHS (
p < 0.001), but neither treatment nor the variety × treatment interaction was significant (
p < 0.05).
PHS variation is known to be significantly influenced by the interaction between variety physiological characteristics and environmental factors such as accumulated temperature after heading, rainfall and humidity during maturation, and dormancy-related genes (
Kang et al. 2018;
Lee et al. 2021). In this study, significant varietal differences in PHS were observed, whereas the effect of storage temperature was not significant under the four treatments, including cold and freezing conditions (
Fig. 2). Therefore, the results suggest that variety-specific characteristics have a greater impact on PHS rate than storage conditions. Early maturing varieties were generally characterized by higher PHS compared with mid-late maturing varieties (
Park et al. 1984). However, exceptions with lower PHS were observed even among early maturing varieties (
Park et al. 2009), making it difficult to generalize PHS exclusively on the basis of ecotypes. These results are consistent with previous observations showing that differences in PHS occur among rice varieties regardless of ecotype, suggesting that variety-specific characteristics play a major role in determining PHS rate. Previous studies have suggested that variety-specific physiological traits such as dormancy persistence and hormone-mediated regulation may contribute to differences in rice PHS among varieties (
Sohn et al. 2021). In our results, we also observed inherent differences in PHS rates among the varieties. However, panicles stored at 4℃ and -20℃ did not show statistically significant differences compared with the control. These results suggest that cold and freezing storage can be used to maintain consistent PHS phenotypes across samples collected on different heading dates. Furthermore, this is expected to enhance the efficiency of large-scale PHS evaluation and improve the reliability of population-based genetic analysis such as QTL mapping, GWAS, and selection decisions in rice breeding programs.
Acknowledgments
This research was funded by the National Institute of Crop and Food Science (NICS) (Project No. PJ01683104), and additional supported by 2025 the RDA Fellowship Program of NICS, Rural Development Administration, Republic of Korea.
Fig. 1Comparison of degree of germination under different storage conditions. Phenotypic variations of PHS among rice varieties under high humidity conditions. Ten varieties are classified as (A) PHS-tolerant (‘Boreumi’, ‘MY362VP’, ‘Joil’, ‘Joun’, ‘Namchan’ and ‘Nampyeong’) and (B) PHS-susceptible (‘Jopyeong’, ‘Baromi2’, ‘Samkwang’, and ‘Chamdongjin’) groups. Panicles were harvested 40 days after heading and incubated for 7 days under 25℃ and over 95% humidity. Scale bar = 5.0 cm.
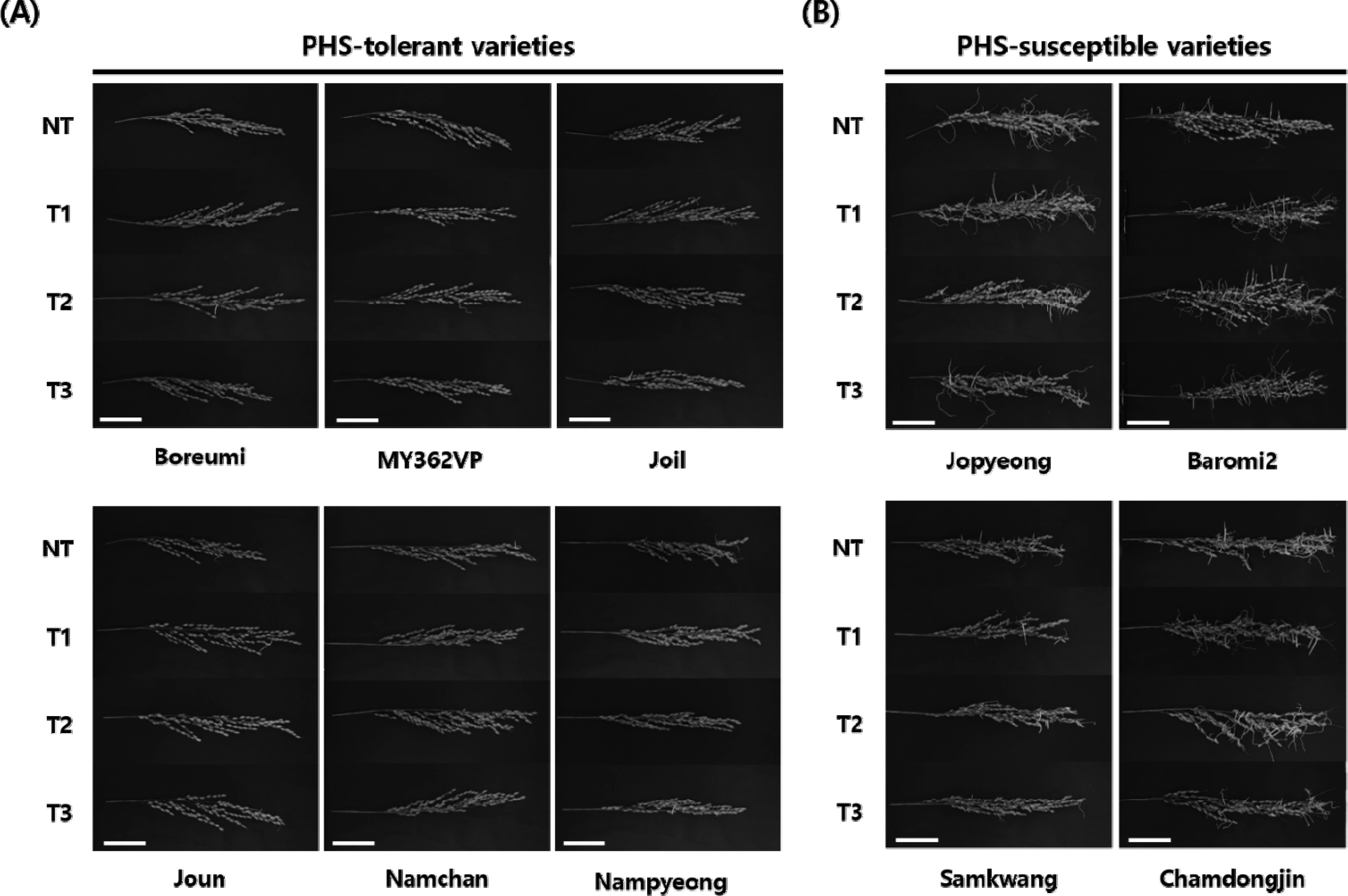
Fig. 2Statistical analysis of PHS rate according to cold and freezing storage. (A) Six varieties with PHS rates below 20% are classified as PHS-tolerant varieties, and (B) four varieties with PHS rates above 20% are classified as PHS-susceptible varieties. Red bars represent the early maturing varieties, and blue bars represent the mid-late maturing varieties. The x-axis indicates the four storage treatments, and the y-axis represents the PHS ratio. NT, No treatment; T1, two weeks at -20℃; T2, four weeks at -20℃; T3, two weeks at 4℃. Bars represent mean values and error bars indicate min–max range (n = 7). ns, no significant
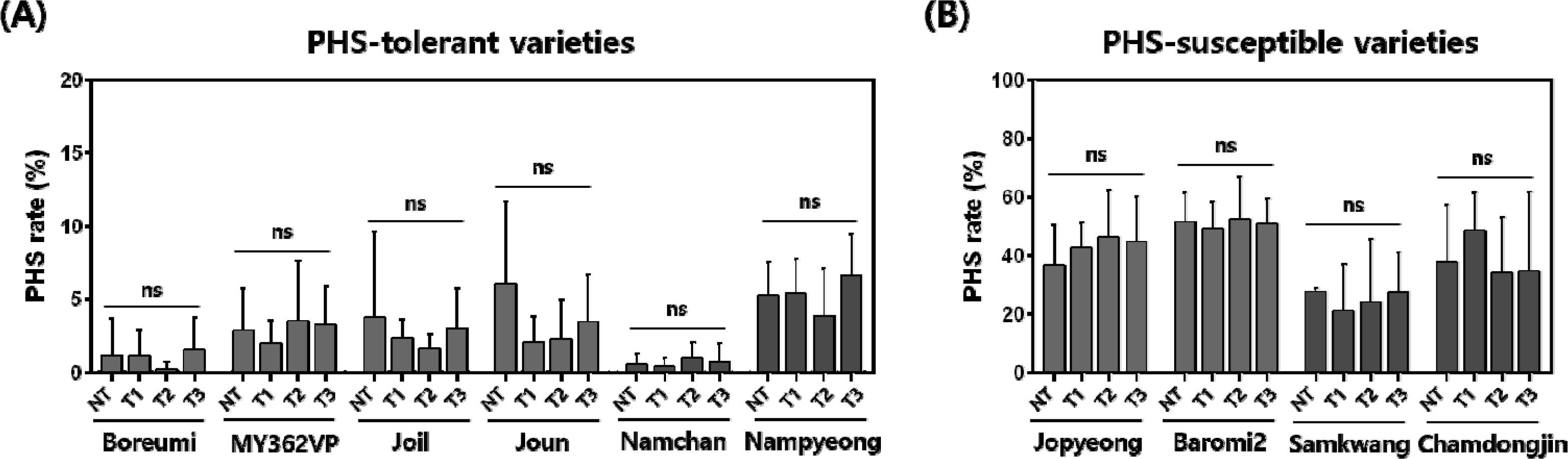
Table 1PHS rate according to storage conditions for ten rice varieties. The table shows varietal variation in PHS rates under different storage treatments. Data are expressed as mean ± standard deviation (n = 7 per storage condition), and Tukey’s HSD test was used for statistical analysis at the 0.05 probability level
Table 1
|
Type |
Ecotype |
Variety (A) |
Storage condition (B) |
|
|
NT |
T1 |
T2 |
T3 |
|
PHS-tolerant |
Early maturing |
‘Boreumi’ |
1.2 ± 2.6 |
1.2 ± 1.8 |
0.3 ± 0.5 |
1.6 ± 2.2 |
|
‘MY362VP’ |
2.9 ± 2.9 |
2.0 ± 1.7 |
3.5 ± 4.1 |
3.3 ± 2.6 |
|
‘Joil’ |
3.8 ± 5.9 |
2.4 ± 1.3 |
1.7 ± 0.9 |
3.0 ± 2.8 |
|
‘Joun’ |
6.1 ± 5.6 |
2.1 ± 1.7 |
2.3 ± 2.7 |
3.5 ± 3.2 |
|
|
Mid-late maturing |
‘Namchan’ |
0.5 ± 0.7 |
0.4 ± 0.6 |
1.0 ± 1.0 |
0.7 ± 1.2 |
|
‘Nampyeong’ |
5.3 ± 2.3 |
5.4 ± 2.4 |
3.8 ± 3.3 |
6.7 ± 2.8 |
|
|
PHS-susceptible |
Early maturing |
‘Jopyeong’ |
36.6 ± 14.0 |
43.0 ± 8.3 |
46.3 ± 16.3 |
45.0 ± 15.4 |
|
‘Baromi2’ |
51.7 ± 9.7 |
49.3 ± 9.1 |
52.7 ± 14.4 |
51.0 ± 8.4 |
|
|
Mid-late maturing |
‘Samkwang’ |
27.9 ± 1.1 |
21.1 ± 16.0 |
24.1 ± 21.7 |
27.5 ± 13.6 |
|
‘Chamdongjin’ |
37.8 ± 19.6 |
48.8 ± 12.8 |
34.1 ± 19.0 |
35.0 ± 26.9 |
|
|
A |
***
|
|
B |
nsz)
|
|
A × B |
ns |
References
- Baek JS, Chung NJ. 2014. Pre-harvest sprouting variation of rice seeds located on each panicle position according to grain filling days. Korean J. Crop Sci. 59(1): 22-26.
- Kang S, Shon J, Kim HS, Kim SJ, Choi JS, Park JH, et al. 2018. Analysis of genetic variation in pre-harvest sprouting at different cumulative temperatures after heading of rice. Korean J. Crop Sci. 63(1): 8-17.
- Kim SJ, Won JG, Ahn DJ, Park SD, Choi CD. 2008. Influence of viviparous germination on quality and yield in rice. Korean J. Crop Sci. 53(spc): 15-18.
- Lee H, Choi M, Hwang W, Jeong J, Yang S, Lee C. 2021. Occurrence of rice preharvest sprouting varies greatly depending on past weather conditions during grain filling. Field Crops Res. 264: 108087
- Park JS, Kim HD. 2009. Viviparous germination characteristics of rice varieties adaptable to central region of Korea. Korean J. Crop Sci. 54(3): 241-248.
- Park KB, Park RK. 1984. Studies on the viviparous germination of Indica×Japonica type varieties in paddy rice. Korean J. Crop Sci. 29(1): 15-18.
- R Core Team.2020. R Core Team R: A language and environment for statistical computing. Foundation for Statistical Computing. Vienna, Austria.
- Shalem Raju R, Hanjagi PS, Awaji SM, Goud BR, Bhaskar SS, Srinivas T, et al. 2024. Enhancing the assessment of pre-harvest sprouting phenotyping in rice: A comprehensive protocol integrating field and laboratory evaluations. Russ. J. Plant Physiol. 71(3): 84
- Sohn SI, Pandian S, Kumar TS, Zoclanclounon YAB, Muthuramalingam P, Shilpha J, et al. 2021. Seed dormancy and pre-harvest sprouting in rice-an updated overview. Int. J. Mol. Sci. 22(21): 11804
- Tai L, Wang HJ, Xu XJ, Sun WH, Ju L, Liu WT, et al. 2021. Pre-harvest sprouting in cereals: genetic and biochemical mechanisms. J. Exp. Bot. 72(8): 2857-2876.
Citations
Citations to this article as recorded by