Abstract
A total of 34 T1 transgenic rice lines overexpressing OgPR1 from wild rice (Oryza grandiglumis) were produced in the previous study. Selection of transgenic plants using hygromycin selection medium was continuously done until T4 generation to find ten homozygous lines. These ten T4 lines were established in the Genetically Modified Organism (GMO) field of NICS (National Institute of Crop Science) in the year of 2007. Phenotypic uniformity and performance was evaluated and compared to the control Dongjin. Two lines were selected for Preliminary Yield Trial (PYT) and Replicated Yield Trial (RYT). Based on morpho-agronomic trait performance, the transgenic plants tended to head later than the control. Culm length was similar to the wild-type Dongjin but the panicle length was relatively reduced. In case of panicle number, transgenic lines showed a little increment compared to wild-type. The shape of grain was nearly the same to wild-type. Yield among lines showed a little difference but was statistically not significant. In terms of physiochemical property of the grain, the transgenic lines showed higher amylose content than the wild-type. When OgPR1-expressing transgenic rice plants were tested against rice blast disease, an enhanced resistance against the disease was observed in the transgenic plants.
-
Key words: Wild rice, Oryza grandiglumis, OgPR1, PYT, RYT
INTRODUCTION
The genus
Oryza consists of 2 cultivated (
O. sativa and
O. glaberrima) and 22 diploid or allotetraploid wild species (
Vaughan et al. 2003;
Jena KK. 2010) and they are the important reservoir of genetic resources and variability for agronomic crops with useful traits such as biotic and abiotic stresses (
Brar and Khush 1997;
Xiao et al. 1998;
Moncada et al. 2001). In the case of
Oryza sativa and
O. glaberrima, they share a common AA genome and can easily be crossed with each other by conventional crossing methods. This sexual compatibility makes it possible to transfer valuable genes from AA genome of wild species into the cultivated rice (
Xiao et al. 1998;
Moncada et al. 2001;
Nguyen et al. 2003). However, it is difficult to transfer genes from wild species into cultivated rice genotypes if they have different genome with the incomplete sexual compatibility or recombination barriers. Nevertheless, several beneficial genes have been successfully transferred to cultivated rice from distantly related species such as
O. officinalis (CC
), O. australiensis (EE), and
O. minuta (BBCC) using advanced techniques of tissue culture and chromosome manipulation (
Jena et al. 1992;
Brar and Khush 1997;
Liu et al. 2002;
Jena et al. 2002).
O. grandiglumis (allotetraploid CCDD) is endemic to Central and South America. This species has large panicles and has potentially useful genes for many agronomic traits such as higher biomass and resistance to diseases and insect (
Heinrichs et al. 1985;
Yu et al. 1997;
Vaughan et al. 2003). A few studies have been conducted to detect genetic variability of
O. grandiglumis (
Shin et al. 2004;
Kim et al. 2005;
Jeon et al. 2008,
2009;
Pak et al. 2009;
Jeon et al. 2012;
Shin et al. 2012). However, attempts to transfer genes from
O. grandiglumis to cultivars have been limited mainly due to incompatibility between unrelated genomes (
Yu et al. 1997). Due to these important traits, this species is of prime importance to the rice breeders. To undermine the function of defense-related gene from wild rice, cloning and characterization of defense-related gene encoding PR-1 from
Oryza grandiglumis (
OgPR1) was previously carried out (unpublished). However, practical evaluation has not been completed in any further generation, especially under the field condition. It is very important to evaluate and check the genetic resource through the subsequent generations. The continuous selection and deployment of generation are the critical procedures for molecular breeding of wild germplasm or any cultivar including wild genetic resources.
Genetic transformation technique has been developed to introduce a gene into foreign species, generally not compatible for cross. Several studies are using this method to assign gene function of unknown or putative genes. Recently, genetic transformation method became a hardcore technique to produce new variety with useful agronomic traits. The biggest advantage of this technnology is to overcome cross incompatibility among various plant species while at the same time it does not require much effort and time to remove undesirable traits after conventional cross. However, it is true that some efforts and time still need to refine the product after introduction of single gene into new genome. Production of GM crop is irreversible trend not only in plant biotech area but in agro-business. So far, many countries have developed various GM crops (
Chandel et al. 2010;
Huesing et al. 2011;
Storer et al. 2012). With future prospect to produce GM crop in Korea, studies dealing with molecular breeding of transgenic plant as this, is a prerequisite for a successful production of GM crops.
In this study,
OgPR1 was transformed into cultivated rice Dongjin to introduce anti-fungal activity. PR genes are involved in many plant-pathogen interactions, such as the synthesis of low molecular weight secondary metabolites (e.g., toxic phytoalexins), cell wall rigidification and the induction of a variety of novel proteins (
Van Loon et al. 1999). Among these PRs, the PR1 class is a dominant group that is induced by pathogens, and is commonly used as a marker for SAR (
Van Loon et al. 1999). Over-expression of some PR proteins in plants has been reported to enhance plant resistance against fungal or bacterial disease (
Alexander et al. 1993;
Broglie et al. 1991). In our previous study,
OgPR1 showed strong resistance against
Botrytis cinearia when introduced into
Arabidopsis thaliana (unpublished). The transgenic plants overexpressing
OgPR1 were screened and crossed/backcrossed to produce near-isogenic line. Selections based on yield component, seed quality and general phenotype have been carried for several generations. Finally, near-isogenic lines harboring
OgPR1 were produced through long period of field and lab experiment. This study aimed to provide some information on the development of field protocol in terms of molecular breeding of transgenic plant.
MATERIALS AND METHODS
Plant materials
In 2006, ten T4 Dongjin transgenic lines were selected by hygromicin test. These plant materials were homozygous but were not evaluated for agronomical traits.
Cultivation condition and maintenance of plant materials
Transgenic rice plants harboring OgPR1 have been selected every year based on hygromycin resistance and GUS expression analysis. In 2006, a total of 10 homozygous lines (T4) were obtained. These lines were selected based on their general agronomic traits and phenotype that were relative to Dongjin. Further selection was done wherein two T4 and two T5 lines were selected for evaluation in PYT for 2 years, and 2 lines of T6 generation for main yield test. Breeding of NILs was conducted at the greenhouse of the National Institute of Crop Science, RDA (Suwon, Korea). Rice transgenic lines were grown in the summer of 2008–2010 at Biohazard Green House and GMO isolated field in the National Institute of Crop Science (NICS), the Rural Development Administration, Suwon, Korea. Rice seeds were soaked in sterile water for 1 day and germinated on wet filter paper and incubated at 30°C under constant light for 3 days. The 35-day-old seedlings were then transplanted into the field with 30 cm × 15 cm spacing. Nitrogen, phosphorous and potassium fertilizers were applied at 9, 4.5 and 5.7 kg/10a, respectively. For the investigation of cross compatibility, fertility of transgenic and NILs were investigated in comparison with that of recurrent parents. The transgenic plants and NILs were evaluated for PYT in two replications and for RYT in three replications.
Assessment of phenology and yield traits
The morpho-agronomic trait performance of transgenic plants and NILs were evaluated in PYT and RYT. Heading date was recorded as the number of days from sowing in the field until 40% of the panicles in the all plants headed. Main agronomical test was carried after dent stage. Plants located in the middle of 10 plants in each plot were chosen for the evaluation of traits such as culum length, panicle length, number of panicles/hill, panicle exsertion and flag leaf length. Data on agronomical traits obtained were average value of the replications. Phenotypic evaluation in PYT and RYT were carried out using 3 to 5 plants for each repeated survey. Culm length was measured in centimeters from the soil surface to the tip of tallest panicle. Panicle length was measured in centimeters from panicle neck to the panicle tip. Panicle number was calculated as the number of panicles per hill (plant). The distance between the bottom of the panicle to the flag-leaf ligule was taken as panicle exsertion length.
Yield factors and yield evaluation
Yield evaluation of the plant materials under PYT was performed 55 to 60 days after heading; full ripe period. Yield components were investigated with the samples obtained from field. The average value of repetition was used as a representative. Spikelets per panicle were calculated as the average number of spikelets in each panicle. Grains per panicle were calculated as the number of fully filled seed per panicle. Ratio of ripened grains was calculated as a percentage: the number of filled spikelets per panicle divided by the number of spikelets per panicle. Days to flowering were determined when 90% of the hills in each treatment plot had at least one tiller that reached anthesis. Days to maturity were determined when 95% of the spikelets within each treatment had turned yellow. Dry matter and yield attributes were determined by randomly sampling 3 hills from each plot. Panicles were hand-threshed and the filled and unfilled spikelets were separated by submerging threshed grains in tap water. The empty and half-filled spikelets were then separated from filled spikelets by using a columnar blower. After drying them thoroughly under the sun, the fresh weight of filled, half-filled, and empty spikelets was determined. Three samples of about 30 g of filled, 15 g of half-filled and 2 g of empty spikelets were collected from each sample. Their corresponding numbers in each subsample were counted and were taken as the number of filled, half-filled and empty spikelets per g.
Grain quality attributes
Grain quality evaluation was investigated with the samples obtained from field. The average value of repetition was used as a representative. Ten grains per line were measured with three replications using vernier caliper for grain length, grain width, and grain length/width ratio. 1000-grain weight was measured in grams as the weight of 250 fully filled grains per field multiplied by four.
Grain physiochemical property
Toyo-taste value was determined using Toyo taste meter (MA-90 series, Toyo Rice Cleanning Machine CO., Japan) in accordance with operation manual (TRCM Co.). The glossiness of cooked rice was determined in two replications using a Toyo-taste value (MC-90A, Toyo Rice Cleaning Machine Co., Ltd., Wakayama). Evaluation of physiochemical property of rice was carried with Near Infrared Spectroscopy (NIR spectrophotometer, FOSS Infratec 1241 Grain Analyzer).
Protein content was determined by micro-Kjeldahl method (Approved Method of the AACC.) using a nitrogen conversion factor 5.95. Amylose content of the brown rice flour was determined by using the method of IRRI (
Juliano 1998). Accurately weighed 100 mg of the defatted sample were taken and 9 ml of 1N NaOH were added. The suspension was thoroughly mixed, and added with distilled water to make a 100 mL solution. For every 5 ml of the sample solution, 1 ml of 1N acetic acid and 2ml of iodine reagent were added. The volume was diluted to 100 ml and the absorbance was measured at a wavelength of 620 nm using Spectronic Instrument (Thermo electron, Evolution 500, USA).
For the Magnaporthe grisea inoculation, two isolates, 02-319 and 90-002 (race KJ-105a), were used in this study. The fungi were grown on potato dextrose agar (PDA) plates and incubated at 25°C under fluorescent light with 12-h photoperiod for 2 weeks. Conidia were dislodged by flooding the plates with sterilized distilled water amended with Tween 20 at 0.05% and scraping the plate surface. The suspension was filtered through three layers of sterile gauze to remove mycelial fragments. Inoculum concentration of the conidial suspension was determined at 5 × 105 conidia/ml using a hemacytometer. Thirty-day-old rice seedlings were inoculated by spraying the conidial suspensions of M. grisea to Dongjin, c.v. Hawsung, Dongjin transgenic and Hawsung NILs. Samples were collected after inoculation at the times described.
Gus expression assay
Expression of GUS gene in rice cells was assayed essentially as previously described (
Rueb and Hensgens 1998) with 5-bromo-4-chloro -3-indolyl glucuronide (X-Gluc) as a substrate. Segment of rice calli, leaf and stems were incubated in phosphate buffer (0.2 M Na
3PO
4, pH 7.0) containing 1 mg/mL X-Gluc, 0.1 % (v/v) Triton X-100 and 10 mM EDTA at 37°C for 4 h. Leaf tissues were washed 4–5 times with 75 % ethanol before visual examination. Tissues that expressed the GUS gene were dark-blue in color.
RESULTS AND DISCUSSION
Agronomic traits of transgenic rice harboring OgPR1 gene from wild rice (Oryza grandiglumis)
A total of 34 T
1 transgenic lines overexpressing
OgPR1 from wild rice (
Oryza grandiglumis) were produced by previous experiment (unpublished). Selection on hygromycin media was continuously done until T
4 generation to find homozygous line. In T
2 selection, 8 lines with relatively large number of seeds were mainly used for seedling selection on media containing 40 ppm hygromycin. This was followed up by GUS staining of plants grown in pots. Finally, 10 homozygous lines were obtained in T
4 generation and used for study (
Table 1). All the seeds from 10 lines survived on the selection medium containing 40 ppm of hygromycin. And this was supported by the positive results in GUS staining analysis. These 10 T
4 lines were planted in GMO field of NICS in the year of 2007 for phenotypic evaluation (
Fig. 1). The selected transgenic lines revealed similar agronomic characteristics to their recurrent parent. However, some lines showed some extent of segregation on culm length and number of panicles, and even showed severe sterility (
Table 1). Heading date, panicle length and growth duration are nearly the same as Dongjin. Most of transgenic lines showed low ratio of ripened grains. One representative individual plant that displayed close phenotype to control Dongjin was selected from each line for use in further experiments. Due to the limiting area of GMO field all 10 lines were not established in PYT. Instead, 2 lines, believed to be the most elite lines, were selected for PYT.
Field selections based on agronomic characteristics of each line and uniformity within line were performed. A total of two lines were selected for PYT evaluation that lasted for 2 consecutive years. In 2008 under PYT, transgenic lines were generally late-heading and had lower value in yield components than the wild-type Dongjin (data not shown). The grain color was similar to that of control but overall appearance was of low quality than the control. Amylose and protein content, and toyo-taste value were similar to control. The investigation of alkali digestion value (ADV) was excluded because japonica type of rice did not reveal clear difference. In the following year (2009) under PYT, transgenic lines were late-heading, producing lower yield (data not shown). Likewise, the grain color was similar to that of control but appearance of grain and toyo-taste value was a little lower than wild-type control.
It is assumed that those minor differences in independent transgenic lines were not directly derived from the transgene but from the interaction of transgene with endogenous genetic background or somaclonal variation produced during tissue culture. Therefore, it is desirable to compare these transgenic lines with wild-type under the natural condition of high disease outbreak or unfavorable cultivation with any known fungal disease. But in this study, there was almost no incidence of fungal disease in the field. Thus, selection of transgenic plant was carried out based on the two year PYT performance in the field.
Two transgenic elite lines were selected because of their morphological similarity to wild-type over the other transgenic lines. These lines were deployed in GMO field at NICS, 2010 (Suwon, Korea). Based on their morpho-agronomic characters, transgenic lines tended to head late (7 or 9 days). Culm length was similar to wild-type Dongjin but panicle length was slightly reduced. In case of panicle number, transgenic lines showed a little increment without statistical significance (
Table 2). For the components of yield, number of spikelet/panicle, percentage of fertile grain, ratio of ripened grains, brown rice recovery, 1000-grain weight, and yield of rough rice/brown rice/milled rice were investigated (
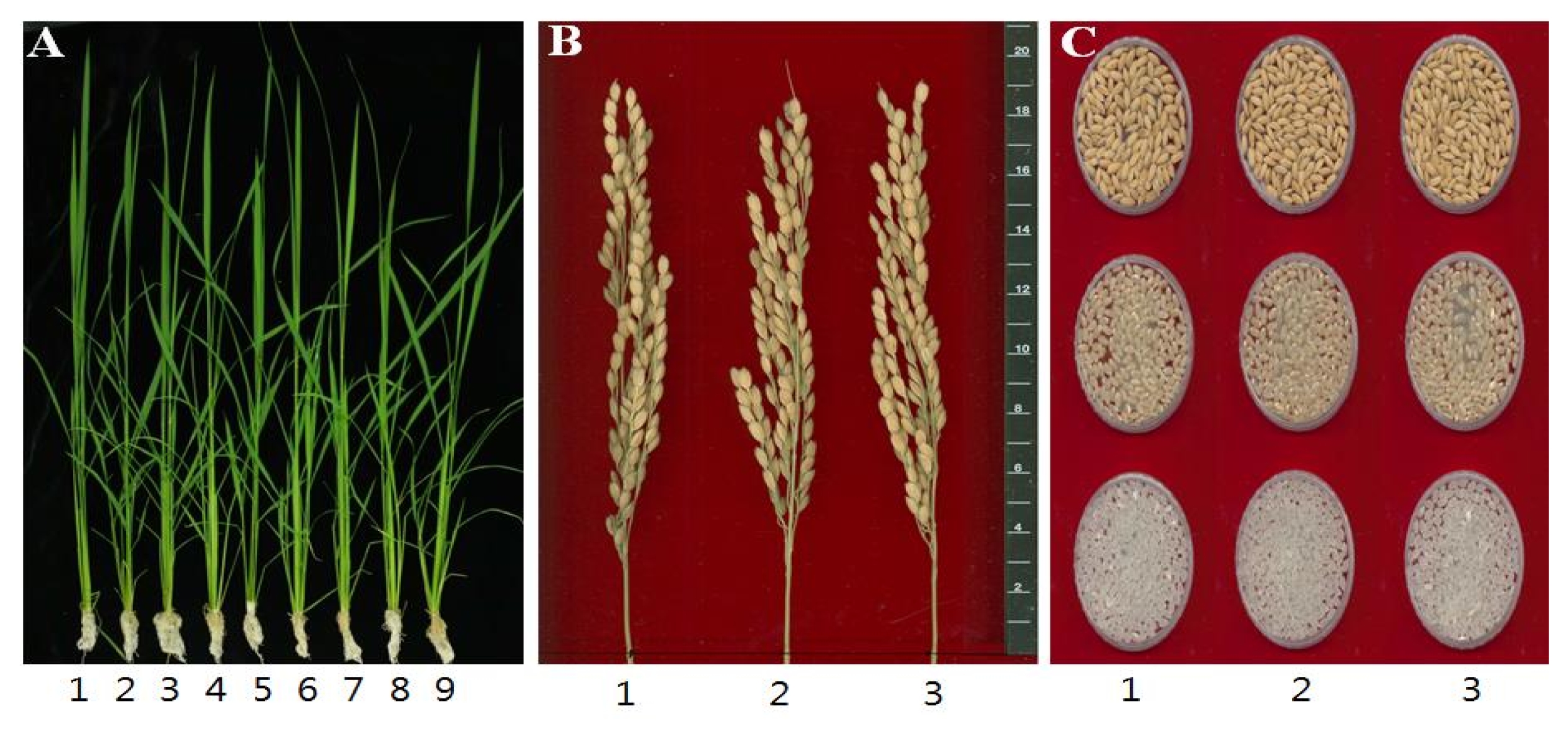
Table 3). The number of spikelet/panicle was seemingly reduced in both transgenic lines but of no significant difference to the control. Percentage of fertile grain of transgenic lines, as well as the ratio of ripened grains, was a little bit higher than wild-type. Further, the 1000-grain weight and yield of rough rice/brown rice/milled rice in transgenic lines were not significantly different from wild-type control. To check any variation in phenotype, the seedling phenotype, mature panicle, and grain shape including color were investigated (
Fig. 2). As shown in the figure, no significant difference was noted between transgenic and wild-plant. Seedling morphology of transgenic lines and wild-plant was nearly the same but some individual transgenic plants were relatively smaller than the control. However, average size of plant was not much different as indicated in
Table 1 &
2. In the case of panicle morphology, transgenic and wild-type were similar as shown in panel B in
Fig. 2. In the analysis of physiochemical property, several traits were checked to see any difference between transgenic lines and wild-type (
Table 4). Grain length and width, brown/rough rice ratio, amylose content, protein content and toyo-taste value were surveyed. The grain length, width and length/width ratio were not significantly different from the control. Amylose content was a little higher in both transgenic lines over the SD value. But protein content was lower in transgenic lines over SD value. Toyo-taste value showed a little higher in the seeds of transgenic lines.
Blast disease resistance of the
OgPR1 transgenic rice plant was tested. Conidial suspensions (5×10
5 conidia/ml) of
Magnaporthe grisea (race KJ-105a) were spray-inoculated onto rice seedlings between the 5th and 6th leaf stages. Disease severity of rice blast on susceptible control (Dongjin) and transgenic plants were investigated (
Fig. 3). As the figure shows, transgenic plant exhibited better resistance against rice blast than the control. This result indicated that introduction of resistance gene from wild rice via genetic transformation is a valuable procedure to bring resistance of wild germplasm into cultivated rice. Selection through many generations did not affect the performance of gene introduced. As pointed out earlier, one of the limitations of the study is the absence of natural fungal disease infections during evaluation in the field which prevented us to evaluate their response to any known fungal diseases. This study did not only demonstrate the production of transgenic rice with fungal disease-resistance gene from wild germplasm but also their deployment and performance in the field. Also, selection based on normal breeding procedure to obtain elite line in T
6 generation through PYT and RYT test were described in this study.
ACKNOWLEDGEMENT
This research was supported by Bio-industry Technology Development Program, Ministry for Food, Agriculture, Forestry and Fisheries, and a grant from the Next-Generation BioGreen 21 Program (Code PJ007978), Rural Development Administration, Republic of Korea.
Fig. 1The selected promising transgenic lines (●) revealed similar agronomic characteristics to their respective recurrent parent(○).

Fig. 2Phenotypic analyses of the rice transgenic lines. A. Dongjin (1) and transgenic plant lines (2–6: Dongjin (OgPR1)-T6(2), 7–9: Dongjin(OgPR1)-T6(3)) at 30 days after transplanting. B. Dongjin (1) and transgenic plant lines (2–3) compared in mature panicles. C. Dongjin (1) and transgenic plant lines (2–3) compared in shape of rough rice (top), brown rice (middle) and milled rice (bottom) grains.
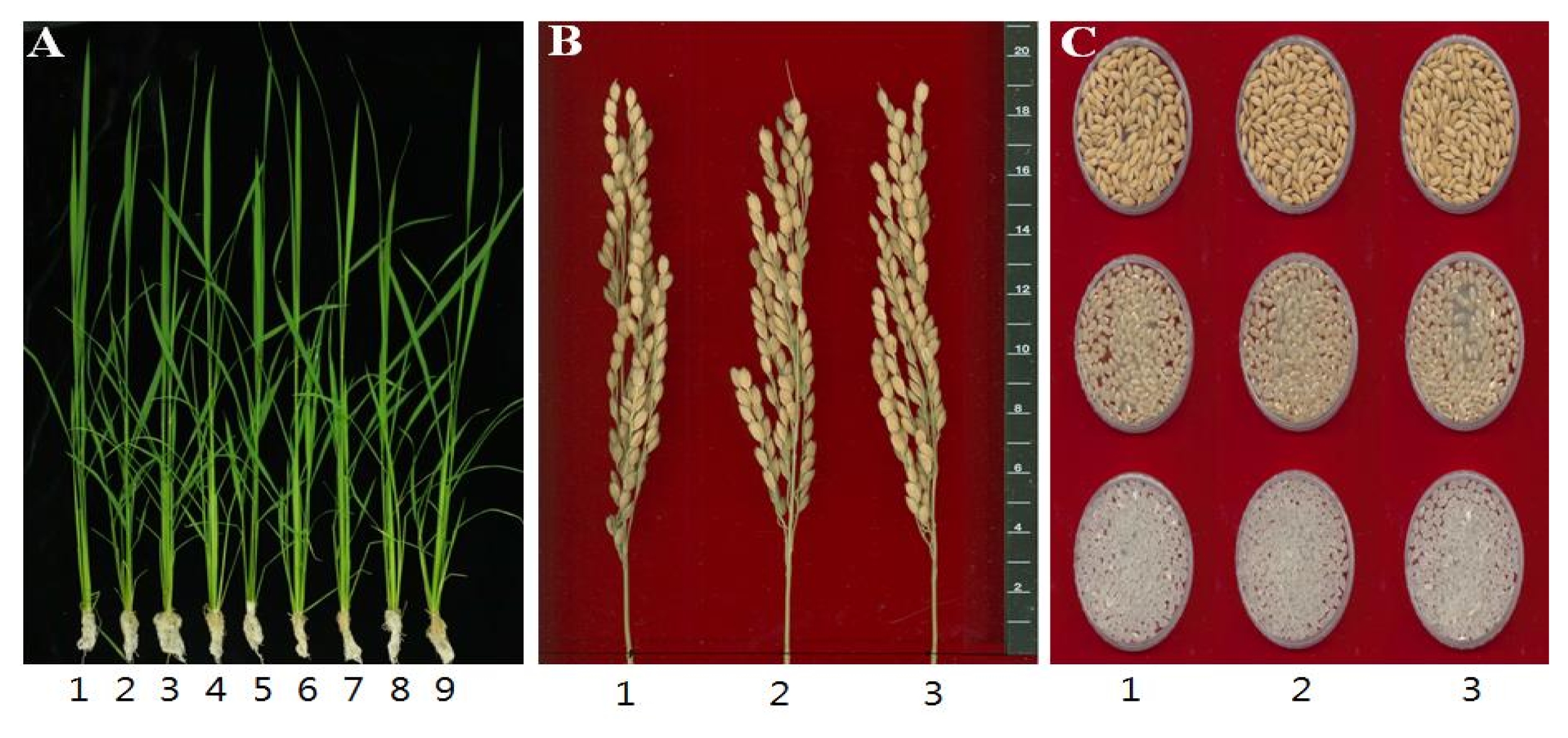
Fig. 3Blast disease resistance of the OgPR1 transgenic rice plant. Disease severity of rice blast on susceptible control (Dongjin) and resistant transgenic plants (1–2:T6 generation). Conidial suspensions (5×105 conidia/ml) of Magnaporthe grisea (race KJ-105a) were spray-inoculated onto rice seedlings between the 5th and 6th leaf stages.

Table 1Basic agronomic characters of the transgenic T4 plant in 2008.
Table 1
|
Line |
Heading date |
Growth duration from seeding to heading (days) |
Culm length (cm) |
Panicle length (cm) |
No. of panicles/hill |
Ratio of ripened grains (%) |
|
Dongjin |
8.24 |
119 |
89 |
21 |
6 |
95.7 |
|
Dongjin(OgPR1)-T4(1) |
8.24 |
118 |
93 |
20 |
7 |
91.2 |
|
Dongjin(OgPR1)-T4(2)***
|
8.24 |
118 |
92 |
19 |
8 |
95.5 |
|
Dongjin(OgPR1)-T4(3)***
|
8.24 |
118 |
88 |
19 |
10 |
93.4 |
|
Dongjin(OgPR1)-T4(4) |
8.24 |
118 |
86 |
20 |
6 |
90.6 |
|
Dongjin(OgPR1)-T4(5) |
8.24 |
118 |
83 |
19 |
8 |
87.7 |
|
Dongjin(OgPR1)-T4(6) |
8.24 |
118 |
87 |
21 |
7 |
89.4 |
|
Dongjin(OgPR1)-T4(7) |
8.24 |
118 |
94 |
19 |
6 |
88.9 |
|
Dongjin(OgPR1)-T4(8) |
8.24 |
118 |
86 |
20 |
7 |
82 |
|
Dongjin(OgPR1)-T4(9) |
8.24 |
118 |
88 |
20 |
6 |
93 |
|
Dongjin(OgPR1)-T4(10) |
8.24 |
118 |
88 |
20 |
5 |
92.7 |
Table 2Major agronomic characters of the transgenic T6 lines of replicated yield trial (RYT) test in 2010
Table 2
|
Line |
Heading date |
Growth duration from seeding to heading (days) |
Culm length (cm) |
Panicle length (cm) |
No. of Panicles/hill |
Panicle exertion(cm) |
Flag leaf length (cm) |
|
Dongjin |
8.16 |
113.0±0.8 |
94.0±0.7 |
19.6±0.7 |
12.3±1.1 |
5.8±0.3 |
25.6±3.5 |
|
Dongjin(OgPR1)-T6(2) |
8.25 |
121.0±0.4 |
95.0±2.1 |
18.4±0.1 |
13.0±2.1 |
5.1±0.7 |
25.1±2.7 |
|
Dongjin(OgPR1)-T6(3) |
8.23 |
120.0±0.8 |
93.0±1.8 |
18.1±0.5 |
12.7±1.0 |
4.4±1.1 |
21.8±1.2 |
Table 3Yield components of transgenic T6 lines of the replicated yield trial (RYT) test in 2010
Table 3
|
Line |
No. of spikelet/p anicle |
Percent of fertile grain (%) |
Ratio of ripened grains (%) |
Brown rice recovery (%) |
1000-grain weight (g) |
Yield in rice (kg/10a) |
|
|
Rough rice |
Brown rice |
Milled rice |
|
Dongjin |
91.0±7.6 |
47.9 |
86.3±1.4 |
82.2±0.3 |
22.3±0.3 |
545.0±1.8 |
448.0±1.6 |
408.0±1.6 |
|
Dongjin(OgPR1)-T6(2) |
89.0±4.5 |
51.9 |
87.9±5.1 |
81.0±0.7 |
22.5±0.1 |
543.0±17.3 |
440.0±11.2 |
400.0±10.1 |
|
Dongjin(OgPR1)-T6(3) |
85.0±2.1 |
49.7 |
87.0±2.6 |
81.2±0.5 |
22.0±0.2 |
553.0±8.9 |
450.0±10.1 |
409.0±9.0 |
Table 4Physiochemical property of transgenic T6 lines of the replicated yield trial (RYT) test in 2010
Table 4
|
Line |
Grain |
Brown/rough rice ratio (%) |
Amylose content(%) |
Protein content (%) |
Toyo-taste value |
|
|
Length (mm) |
Width (mm) |
length/width ratio |
|
Dongjin |
4.97±0.026 |
2.78±0.014 |
1.79±0.004 |
85.4±2.1 |
19.3±0.5 |
6.2±0.1 |
76.3±0.4 |
|
Dongjin(OgPR1)-T6(2) |
5.04±0.004 |
2.78±0.018 |
1.81±0.010 |
90.2±3.8 |
20.9±0.7 |
5.8±0.1 |
79.6±1.7 |
|
Dongjin(OgPR1)-T6(3) |
5.06±0.018 |
2.75±0.024 |
1.84±0.012 |
88.2±3.1 |
20.7±0.7 |
5.7±0.1 |
77.5±0.5 |
References
- AACC.2000. Approved Method of the AACC. 10th ed. Method 46-12. The American Association of Cereal Chemists. St. Paul, MN, USA.
- Alexander D, Goodman RM, Gut-Rella M, Glascock C, Weymann K, Friedrich L, Maddox D, Ahl-Goy P, Luntz T, Ward E, Ryals J. 1993. Increased tolerance to two oomycete pathogens in transgenic tobacco expressing pathogenesis-related 1a. Pro Natl Acad Sci USA. 90: 7327-7331.
- Broglie K, Chet I, Holliday M, Cressman R, Biddle P, Knowlton S, Mauvais CJ, Brogile R. 1991. Transgenic plant with enhanced resistance to the fungal Rhizoctonia solani. Science. 254: 1194-1197.
- Brar DS, Khush GS. 1997. Alien introgression in rice. Plant Mol Biol. 35: 35-47.
- Chandel G, Datta K, Datta SK. 2010. Detection of genomic changes in transgenic Bt rice populations through genetic fingerprinting using amplified fragment length polymorphism (AFLP). GM Crops. 1(5): 327-336.
- Heinrichs EA, Medrano FG, Rapusas HR. 1985. Genetic evaluation for insect resistance in rice. International Rice Research Institute. Manila.
- Huesing J, Romeis J, Ellstrand N, Raybould A, Hellmich R, Wolt J, Ehlers J, Dabiré C, Fatokun C, Hokanson K, Ishiyaku MF, Margam V, Obokoh N, Mignouna J, Nangayo F, Ouedraogo J, Pasquet R, Pittendrigh B, Schaal B, Stein J, Tamò M, Murdock L. 2011. Regulatory considerations surrounding the deployment of Bt-expressing cowpea in Africa: report of the deliberations of an expert panel. GM Crops. 2(3): 211-224.
- Jena KK, Khush GS, Kochert G. 1992. RFLP analysis of rice (Oryza sativa L) introgression lines. Theor Appl Genet. 84: 608-616.
- Jena KK, Pasalu IC, Rao YK, Varalaxmi Y, Krishnaiah K, Khush GS, Kochert G. 2002. Molecular tagging of a gene for resistance to brown planthopper in rice (Oryza sativa L). Euphytica. 129: 81-88.
- Jena KK. 2010. The species of the genus Oryza and transfer of useful genes from wild species into cultivated rice, O. sativa. Breeding Science. 60: 518-523.
- Juliano BO. 1988. IRRI Annual Report for 1987. International Rice Research Institute. Los Baños, Philippines: pp. 43.
- Jeon EH, Chung ES, Pak JH, Nam JS, Cho SK, Shin SH, Kim DH, Kim GT, Lee JH, Kang KH, Chung YS. 2008. Overexpression of OgPAE1 from wild rice confers fungal resistance against Botrytis cinerea. J Plant Res. 121(4): 435-40.
- Jeon EH, Chung ES, Lee HY, Pak JH, Kim HJ, Lee JH, Moon BJ, Jeung JI, Shin SH, Chung YS. 2009. Ectopic Expression of Wild Rice OgGRP Gene Encoding a Glycine Rich Cell Wall Protein Confers Resistance to Botrytis cinerea Pathogen on Arabidopsis. The Plant Pathology Journal. 25(2): 193-198.
- Jeon EH, Pak JH, Kim MJ, Kim HJ, Shin SH, Lee JH, Kim DH, Oh JS, Oh BJ, Jung HW, Chung YS. 2012. Ectopic expression of ubiquitin-conjugating enzyme gene from wild rice, OgUBC1, confers resistance against UV-B radiation and Botrytis infection in Arabidopsis thaliana. Biochem Biophys Res Commun. 427(2): 309-14.
- Kim KM, Cho SK, Shin SH, Kim GT, Lee JH, Oh BJ, Kang KH, Hong JC, Choi JY, Shin JS, Chung YS. 2005. Analysis of differentially expressed transcripts of fungal elicitor- and wound-treated wild rice (Oryza grandiglumis). J Plant Res. 118(5): 347-54.
- Liu G, Lu G, Zeng L, Wang G-L. 2002. Two broad-spectrum blast resistance genes, Pi9(t) and Pi2(t), are physically linked on rice chromosome 6. Mol Genet Genomics. 267: 472-480.
- Moncada P, Martínez CP, Borrero J, Chatel M, Gauch H Jr, Guimarães E, Tohme J, McCouch SR. 2001. Quantitative trait loci for yield and yield components in an Oryza sativa x Oryza rufipogon BC2F2 population evaluated in an upland environment. Theor Appl Genet. 102: 41-52.
- Nguyen BD, Brar DS, Bui BC, Nguyen TV, Pham LN, Nguyen HT. 2003. Identification and mapping of the QTL for aluminum tolerance introgressed from the new source, Oryza rufipogon Griff. into indica rice (O. sativa L). Theor Appl Genet. 106: 583-593.
- Pak JH, Chung ES, Shin SH, Jeon EH, Kim MJ, Lee HY, Jeung JU, Hyung NI, Lee JH, Chung YS. 2009. Enhanced fungal resistance in Arabidopsis expressing wild rice PR-3 (OgChitIVa) encoding chitinase class IV. Plant Biotechnology Reports. 3(2): 147-155.
- Rueb S, Hensgens L. 1998. Improved histochemical staining for b-glucuronidase activity in monocotyledonous plants. Rice Genetic Newsletters. 6: 168-169.
- Shin Sh, Im Hh, Lee JH, Kim DH, Chung WB, Kang KH, Cho SK, Shin JS, Chung YS. 2004. Genes of Wild Rice (Oryza grandiglumis) Induced by Wounding and Yeast Extract. Journal of life science. 14(4): 650-656.
- Shin Sh, Pak Jh, Kim Mj, Kim Hj, Lee Jh, Kim Dh, Choi Hk, Kang Kh, Jeong Ju, Kang Cs, Jung Hw, Chung Ys. 2012. Cloning and Characterization of Pathogenesis-related Gene 10a (OgPR10a) Derived from Wild Rice (Oryza grandiglumis). Korean J Breed. 44(1): 1-10.
- Storer NP, Thompson GD, Head GP. 2012. Application of pyramided traits against Lepidoptera in insect resistance management for Bt crops. GM Crops Food. 3(3): 154-162.
- Van Loon LC, Van Strien EA. 1999. The families of pathogenesis-related proteins, their activities, and comparative analysis of PR-1 type proteins. Physiological and Molecular Plant Pathology. 55: 85-97.
- Vaughan DA, Morishima H, Kadowaki K. 2003. Diversity in the Oryza genus. Curr Opin Plant Biol. 6(2): 139-146.
- Xiao J, Li J, Grandillo S, Ahn SN, Yuna L, Tanksley SD, McCouch SR. 1998. Identification of trait-improving quantitative trait loci alleles from a wild rice relative, Oryza rufipogon. Genetics. 150: 899-909.
- Yu GJ, Kwak TS, Kang KH, Moon HP. 1997. Efficiency of backcrossing and ovule culture in an interspecific cross between O. sativa L. and O. grandiglumis prod. Korean J Breed. 29: 448-452.